[カテゴリー別保管庫] 脳科学メモ
2025年05月21日
■ パレイドリアと「シミュラクラ現象」
大学で意識についての講義をするときに、いろいろ錯覚を見てもらったりするのだけど、そこでの質問やコメントに「シミュラクラ現象」という言葉が出てくることがある。
認知心理学の分野ではパレイドリアPareidoliaという言葉があって、「壁のシミが人の顔のように見えてしまう」という現象をパレイドリアと呼ぶ。「シミュラクラ現象」というのが指しているのは、典型的には「コンセントの3つの穴が人の顔のように見える」というもので、これはパレイドリアの一種であり、わざわざ「シミュラクラ現象」という言葉を使う必要がない。
そもそも「シミュラクラ現象」という言葉は日本でしか通用しない言葉だ。それはwikipediaの「シミュラクラ現象」の項目を見ればわかる。日本語版しか無い。
そういうわけで、講義で学生から「シミュラクラ現象」についてのコメントをもらった場合には「それは学問的にはパレイドリアと呼ばれるものであって、「シミュラクラ現象」という言葉は日本のネットカルチャーでのみ通用する言葉なのでお気をつけください。」と返事するようにしている。
言葉は生き物なのだから「シミュラクラ現象」という言葉自体を禁止してもしょうがない。だから、ネットに向かって「「シミュラクラ現象」という言葉は誤用です。使うのはやめましょう」とか喧伝しようとは自分は思わない。
とはいえ、「シミュラクラ現象」という言葉のルーツは抑えておきたい。初出はいつかLLMに聞いてみたら、「初出を見つけるのは難しいけど、初期の使用例については2000-2005年の範囲でググってみましょう」とアドバイスされた。
すると2005年の記事が二つ見つかった。
それは「シミュラクラ現象(類像現象)」かもしれません。シミュラクラ現象とは3つの点が集まった図形が人間の顔のように見えるという錯覚現象です。ほら、よくマンホールのフタの模様や、天井のシミが人の顔のように見えてくることがありますよね。もともとはSF作家・フィリップ・K・ディックの小説から生まれた言葉だそうで。
「こうゆうもののコトを「シミュラクラ」と呼ぶ。●が三角に3つ並べば顔に見えるとまで言われる。人間にはあらゆるものを顔に見立ててしまう本能が備わっているらしい。(…)何を言いたいかっちゅーと、心霊写真と言われるもののすべて、とまで言うつもりはないが、9割方はこの「シミュラクラ」だとおれは思っているのだ。」
どちらとも心霊写真の文脈で、それは錯覚だ、シミュラクラ現象で説明できる、というものだ。ここでひとつ興味深いことが書かれている。
もともとはSF作家・フィリップ・K・ディックの小説から生まれた言葉
元々シミュラクラSimulacreという言葉は、ポストモダンの哲学者ジャン・ボードリヤールによる「シミュラークルとシミュレーション」から来ている。(BTの記事の解説がわかりやすかった)
そしてそれの影響を受けて、フィリップ・K・ディックはSF作品として「シミュラクラ」という作品を書いている。私は未読だが、人間そっくりに作られたシミュラクラが出てくる話だそうだ。
というわけで、「シミュラクラ現象」という言葉がオカルトとか心霊写真とかのサブカルチャーの文脈で出てきたことを考えると、はじめてこの言葉を使った人は、ボードリヤールを引用したというよりは、フィリップ・K・ディックを引用したのではないかと想像できる。
さて、認知心理学の分野でパレイドリアの専門家といえばまっさきに浮かぶのは高橋康介さんだ。彼は「なぜ壁のシミが顔に見えるのか ―パレイドリアとアニマシーの認知心理学―」 という本を書いている。
Twitter(当時)だと、以下のスレッドが詳しい。
パレイドリアの話をすると「それシミュラクラでは?」という反応がよくあります。wikipediaの力だろうか。それともPKディックのこれ? https://amazon.co.jp/dp/B077QN78N1/ 必要に迫られてパレイドリアとシミュラクラの語源についてかなり調べたことがあるんですが、結局正確なところはわかりませんでした。Twitter 2020
上述の書籍の中でも、「シミュラクラ現象」について手短に触れている。
パレイドリアとシミュラクラはともに同じような概念を示すものだが、パレイドリアが見えてしまう「現象」を示すのに対して、シミュラクラは見えてしまう現象のきっかけとなる「モノ」を指し示す意味合いが強い。…「シミュラクラ現象」といえば、それはパレイドリアとほぼ同義であると思ってよいだろう。(p.46)
ということで、「シミュラクラ現象」という言葉をむやみに切り捨てることなく、考察している。(とはいえこの本で「シミュラクラ現象」が出てくるのは、あくまでもこの部分だけにすぎない。)
というわけで、今後は質問をもらったら、この記事を参照できるようにした。
2023年11月04日
■ 認知神経リハビリテーション学会で自由エネルギー原理入門の講演をしました(スライドあり)
認知神経リハビリテーション学会で講演をしてきました。
じつは認知神経リハビリテーション学会学術集会で特別講演をするのは5年ぶり2回目。前回は盲視と半側空間無視の話をしました。そのときの懇親会で宮本省三先生(会長)に「なんか一曲」と言われてS&Gの「明日に架ける橋」を歌ったエピソードがこちら:「会長からのメッセージNo.89 『ハプニング』」
今回は第23回認知神経リハビリテーション学会学術集会で特別講演「能動的推論と運動制御」を行いました。今までの自由エネルギー原理入門の話を極限まで噛み砕いたものを新たに作成(ベイズの定理の公式すら外した)。さらに新ネタで、これまで避けてきた運動制御に関する記載を追加しました。とくに重要なのは、最適制御理論との比較を追加したこと。
こちらがスライド:
認知神経リハビリテーション学会2023公開スライド
当日使ったものから、著作権を考慮すべき部分について削除または差し替えを行っております。ダウンロード可能。
こちらが要旨:
われわれの知覚と行為は密接につながっている。この事態についてヴァレラは「身体化された心」で「知覚とは、知覚的に導かれた行為のことである」と再帰的に表現した。このような知覚-行為サイクルの更新則を提案したのがフリストンの自由エネルギー原理だ。自由エネルギー原理において行為とは環境をよりよく知る認知の過程の一部であり、この過程を説明する計算論的モデルのことを能動的推論と呼ぶ。能動的推論において学習、発達、損傷からの機能回復はどれも(環境と行為と感覚受容の関係を表現した)生成モデルの変容として統一的に扱うことができる。このようにして能動的推論は、ペルフェッティの「運動とは認知である」「回復とは学習である」を実現している。
また能動的推論は、運動制御について従来の理論と大きく異なる見方をする。従来の理論では、大脳が出す運動指令によって効果器(筋肉)が駆動され、運動指令の遠心性コピーと感覚器からのフィードバックによって正確な運動制御が行われる。いっぽう能動的推論では、大脳が出す信号は行為の結果の感覚入力の予測である。たとえば到達運動においては、手を伸ばした状態での筋紡錘の活動を予測する。この予測と、筋紡錘からの感覚入力との誤差が脊髄で計算され、それがゼロになるまで運動が行われる。
能動的推論による説明はペルフェッティの言う「運動器とは情報器官である」そして行為における運動イメージの重視とよく合致している。 能動的推論はまだ新しい考えであり、実験的検証が充分ではない。しかし、第4世代のリハビリテーション理論である認知神経リハビリテーションにおいて、能動的推論は重要な理論的基盤となる可能性がある。本講演では以上のことについて、なるべく数式を使わずに平易に説明することを目指す。
さて、感想のまえに背景説明をしておくと、認知神経リハビリテーション学会というのは、第4世代リハビリテーション理論として「認知運動療法」を提唱したカルロ・ペルフェッティ(1940-2020)の考えを国内に導入した、高知医療学院の宮本省三先生が会長となって運営されている学会。
認知運動療法については以下の本がわかりやすい。
- 脳のなかの身体 認知運動療法の挑戦 (講談社現代新書) 宮本省三
- 認知神経リハビリテーション入門 カルロ・ペルフェッティ
認知運動療法とはなにか。私の理解によれば、それはペルフェッティの言葉で簡潔に2文で表現できる:「運動とは認知である」「回復とは学習である」
つまり、身体性を重視したうえで、知覚と運動とを分離せずに、両者をひとつながりの過程として捉えるエナクティヴィズムの考えに基づいて(注)、リハビリテーションについて、損傷によって失われた機能を回復するというよりは、新たな身体と環境の関係を構築するという側面を重視する。
(注: じっさい、ペルフェッティはヴァレラとマトゥラーナに言及する)
そういうわけで、この学会は実践的な側面が強くて、発表者の多くは病院勤務のセラピストの方で占められている。(じつはこの学会とは別に、日本リハビリテーション医学会、日本神経理学療法学会、日本ニューロリハビリテーション学会などの学会がある。このへん、医師とセラピストの関係とかいろいろややこしそうなのだが、突っ込んだことは聞かなかった。)
そういう背景があるのを踏まえて、今回のスライドを準備するに当たっては、エナクティブな考えとの親和性を強調する方向へ持ってゆくこと、そのためにも数式で迷子にならないようにする、ということで数式をほぼ削った。(ベイズの公式すら書いてない)
それでも、「知覚において世界を知ること」と「行動によって新たな感覚入力を探索すること」どちらともが適応的であるのだ、ということが伝わることを目指した。
そして、自由エネルギー原理について興味があって、自分で調べたことがある聴衆に届けるよりも、今回はじめて自由エネルギー原理という言葉を知った人が、ちょっとこれから学んでみようかと思わせるように難易度の調整をした。
さてそれがうまくいったかどうか。講演の後に感想を届けてくれた人は「はじめてFEPについて聞いたけど、面白いと思った」「難しかったけど重要だということはわかった」など、よい反響はあったので、ちょっとほっとした。
あと講演後に何度かあった質問は「この次はどうすればよいでしょうか」というものだった。わたしの日本語総説ふたつを紹介しておいた(スライドp.24にあり)。
- 吉田 正俊, 田口 茂 (2018) 「自由エネルギー原理と視覚的意識」 日本神経回路学会誌 25(3) 53-70
- 吉田 正俊, 宮園 健吾, 西尾 慶之, 山下 祐一, 鈴木 啓介 (2023) 「自由エネルギー原理,能動的視覚,サリエンス」 人工知能. 38(6) 787-795.
でもそれでは質問に答えてないかと思った。むしろ質問は「どのように臨床に役立てていくことができるのでしょうか」ということだったと思う。そういうわけでわたしが答えたのは、「生成モデルという視点で現象を分解してみる訓練に使うのがよいのではないでしょうか」というものだった。(FEP論文がしばしばやる、トイモデルでのシミュレーションではなくて。)
(これはあとでスライドに追加しようと思うけど、いまは文章で追加だけしておく。) たとえば半側空間無視のテストとして使われる線分抹消課題(のサッケード版)はスライドp.119にあるような生成モデルを想定することができる。
そうすることによって、「患者の視線が右側に偏る」という現象が、じつのところどの過程から起きているのか、それは「右側に視線が傾いていることに気がついてない」のかもしれないし、「左に目を向けても得るものがない」と思ってるのかもしれない(スライドp.119)。
そこを患者自身に言語化してもらう、そしてそのためにうまい質問を投げかける、このような「仮説、実験、検証のサイクル」を回す、これはペルフェッティの認知運動療法における言語の活用のキモでもある。
生成モデルを作るためには、そこで起きている現象を分解し、なにを感覚として感知し、なにを知覚として我々は推定している(信念を持っているか)、そしてそれらがどのような階層関係を持っているか(スライドp.106にこのような階層関係の例がある)、それらを明確にしてゆくことが必要なのだけど、それってまさに認知神経療法が重視している、「仮説、実験、検証のサイクル」を回すことを助けるでしょう、ということ。
これは「自由エネルギー原理」そのものというよりは、「ベイズモデリングを活用しよう」というメッセージなのだけど、実践家にとって役に立つものを逃げずに提示するならば、これ(「問題となっている現象について生成モデルを作る」)になると思う。
ほかにもいろいろ語りたいことはある。たとえば、特別講演のもう一人は造形作家の岡崎乾二郎さんだったのだけど、懇親会で岡崎さんの隣りでたくさん話をすることができたのはすごくよかった。(スライドp.111に引用した文學界インタビューはおすすめ)
あと宮本省三さんといろいろ議論できたのはたいへん有益で、とくにメタ学習を考える際に推測だけでなく、アブダクションが必要であり、そこにはアナロジーが不可欠という考えに至ったのも、今回の成果だった。(鈴木宏昭「類似と思考」の意義が掴めてきたかも。)
そのへんについてはまた別の機会に書きたい。
以下はBlueskyに書いた雑談。

懇親会に行く道で水道橋駅の歩道橋を渡ろうとしたら、芳文社があるのに気づいた。きらら系列の総本山がこんなところにあるとは知らなかった。(竹書房が飯田橋にあるのはポプテピピックで知ったけど。)
次の日は本郷三丁目を探索したら、万定フルーツパーラーが休業状態であることを知った。奥の三叉路のパン屋もなくなって、駐車場になってた。

札幌に帰ってきて、研究室に戻って諸々の片付けを終了させた。そのあとで、深夜までやってるスーパーに向かう。
自転車を漕いで大通り(環状通)に出ると、ほとんど車がない。たまに車が通り過ぎると、静けさが広がる。そのとき、なんだか「帰ってきた」という実感が湧いてきた。
札幌を「自分の帰るところだ」と体が感じていることを知って、これはエモいなだと思った。でもそのエモさに心が震えることができない。
けっきょくスーパーでは半額寿司と半額刺し身をゲット。Twitterにはそちらのことを書いてポストしておいた。
2022年05月30日
■ ベイズ更新
ベイズ更新では、そのつど与えられたデータから計算した事後分布を、次のデータが与えられたときは事前分布として使うことで、事後分布を逐次更新してゆく。
たとえば一定のパラメーター を持つ生成過程(たとえば2項分布)から生成されるデータ があったとすると、ベイズの定理より、
となる。これはデータ を一挙に与えられた場合だけど、代わりにデータ をひとつずつ順番に取得して事後分布を更新するのがベイズ更新。まず から始めると、
こうしてできた事後分布 を次の事前分布 のところに入れる。(分母のevidenceは に依存しない定数なので で表記しておく。)
なんでそんなことが可能かというと、 を持つ生成過程から生成されるデータ が独立だから、という仮定が入ってる。単純化するために の2つだけで考えてみる。
もしここで を与えたものでの の条件付き独立
が成り立つなら、
となり、事後分布 を次の事前分布 に代入したものになってる。
分母は定数なのでこのままで十分だけど、もしさらに の独立
が成り立つなら、
となって、分母もきれいになる。条件付き独立が成り立っても、独立は必ずしも成り立たないので、いちおう分けて考えた。でも元々の設定「一定のパラメーター を持つ生成過程(たとえば2項分布)から生成されるデータ 」からすると、たぶん独立も成り立つ。つまり、 を選ぶ順番変えても成り立つということだから。
ここまでの話はパラメーター が一定であることを前提としていた。途中で が変わる場合は成り立たない。そういう場合は をそのつど推定するべきhidden variableとして扱うことになる。
StackExchangeにあるとおりだけど、ふたたび だけの例で書くと、
こうなるので、事前分布 * 尤度 ではなくなって、transition process を考慮する必要が出てくる。
参考:
2022年02月20日
■ DKL色空間の図
必要になったので、ひさびさにDKL空間の図を作ってみた。RGB 8ビット*3の立方体からの変換なので平行六面体になる。それは3Dプロットしてみるとよくわかる。
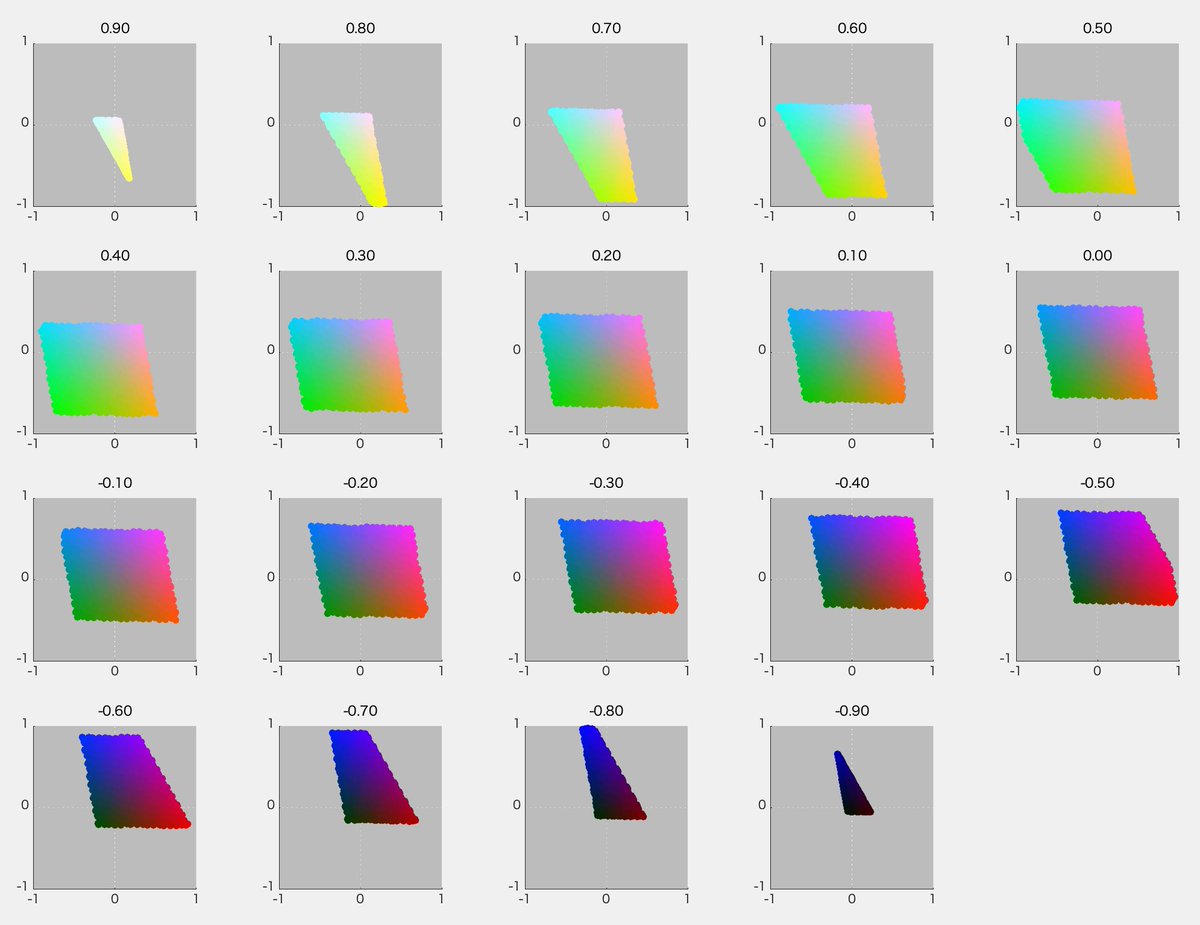
といいつつ計算にハマった。あーわかった、rgbの立方体の12辺上の点だけぜんぶ3Dプロットして、ぐるぐる回してやればいいのか。明日やる。
さっそくやってみた。ちゃんと平行六面体になった(図1)。図2はS-Lumを潰した射影、図3はL-Mを潰した射影、図4はL+Mを潰した射影。図4が正6角形になったのはそれぞれの軸を-1~1に収めるように標準化したから。Red-CyanやBlue-YellowがDKL空間の軸からずれているのが確認できて納得いった。
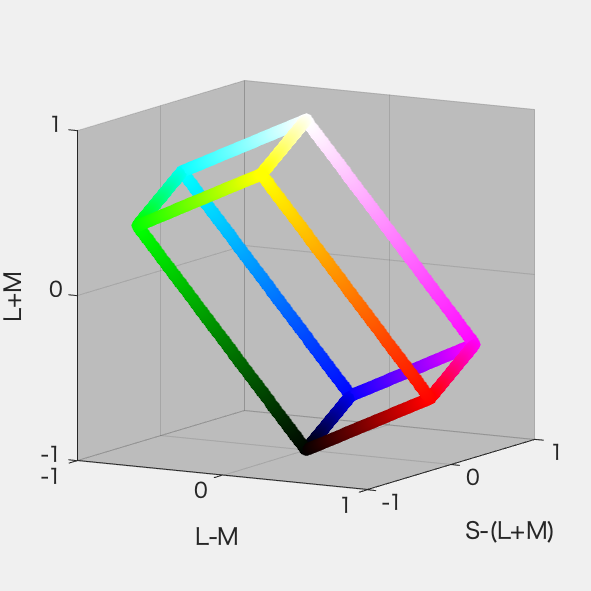
図1
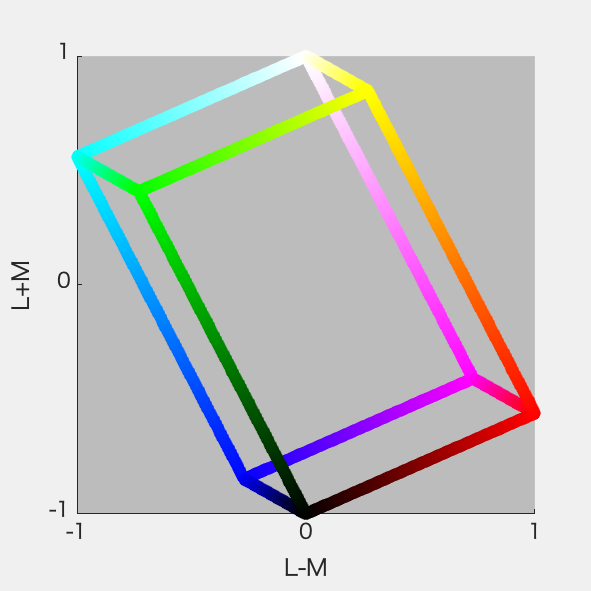
図2
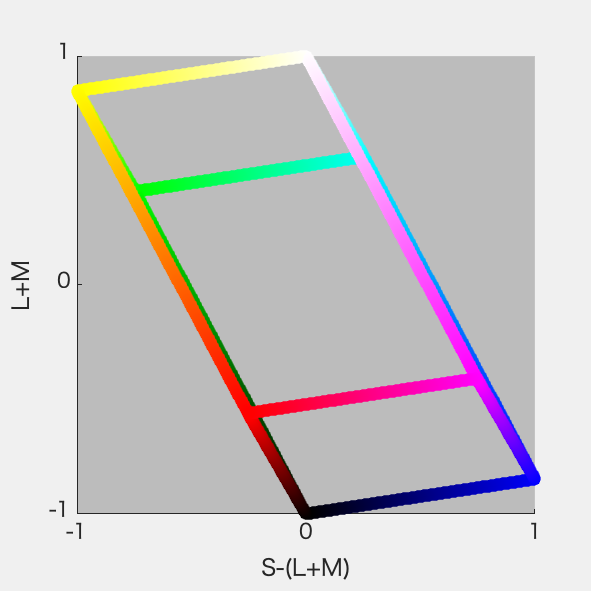
図3
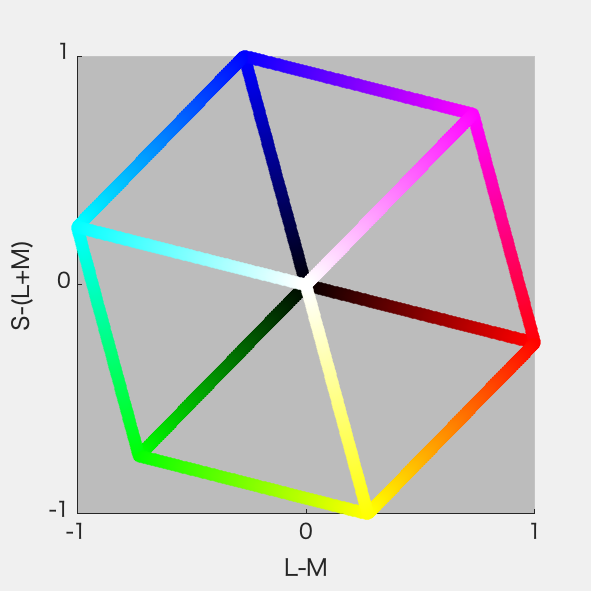
図4
2021年08月02日
■ 研究関連ツイートまとめ(2021年1月)
The New York TimesによるJohn Conwayのライフゲームの記事。 初出を知らなかったけど、Scientific Americanのマーチン・ガードナーのコラム(1970)でJohn Conwayが送ったレターが紹介されたのが初出とのこと。(セルラ・オートマタ自体はフォン・ノイマンだから1940年代。)
なるほど!Varelaらによるオートポイエーシスの2次元セルラ・オートマタの論文が出版されたのは1974年で、たった5つの文献しか引用してないのだけど、そのひとつはこのGardnerのSci Am 1970の記事だった。1974論文内ではライフゲームの発展型とは決して言ってないのだけど。
病院で待っている間に「0と1から意識は生まれるか――意識・時間・実在をめぐるハッシー式思考実験」を読了したけど、思ったよりも面白かった。意識は情報処理ではなくて、それに付随する物質が必要」「時間と空間は実在しない」「エントロピーは時間対称」(渡辺慧)とか、ニコラス・ハンフリーの「内なる目」を参照した意識の社会説(他者の推定する必要から、自分を内的にモニタする能力が進化した)とか。細部はいろいろ納得いかないところもあったが、自分が面白うと思っている方向にいいかんじに当てはまっていた。
これをとっかかりに、いろいろ手を広げることができそう。あと、「時間とエントロピーと重力」とかあのへんを読みたくなってきた。
ここでの「エントロピーは時間対称」というのは時間tで水の中にインクの一滴が固まっている状態があったとして、t+Tでもt-Tでもどちらもエントロピーは高くなっているという話。つまり、スポイトでインクを落とすというような人為的なことをしないならば、たまたまtでインクの一滴が集まった瞬間を見たということを想定しているようだ。
それで連想したのが、カウフマンの本にあった「束縛条件」だった。火薬の爆発を砲弾の移動という仕事に変えるためには、大砲の筒が必要というやつ。同様に、インクを垂らすとか、二重スリットを用意するといった人為的な操作がかならず関わっていて、そうでない場合、「たまたま時間tでインクの一滴が集まった瞬間を観測した」というようなものすごい偶然に身を任せないといけなくなる。まだ言語化できてないけど、「人為的な操作」というのが入ることで、因果とかややこしい話になるのではないかと思った。
「〈こころ〉はどこから来て,どこへ行くのか」 下條さんの章を読むつもりが、中沢新一の章が予想外に面白かった。ハイエクの「感覚秩序」を引いて脳と心の関係を「直接的な因果関係ではなく、構造に関する情報を運ぶ作用をするものを媒介にしながら、つながりあっている」とあり、ホモロジーや圏論にも言及している。中沢新一は読んだことがなかったが、ポスト構造主義から来てる人だから、こういうことは言うかも。ハイエクについても経済での「自生的秩序」に連なる精神の理論を持っていたことに興味を持ったので、次は「ハイエクの経済思想」を借りてくる。
The world is not a theorem スチュアート・カウフマンの新しい論文。生物の進化における多様性を作る方向は(数式などによる)形式化はできない、というこれまでの主張を、アフォーダンスの定式化という側面から議論したものっぽい。
「生命と自由」渡辺慧を読んでいたら「人間の眼は、一つ二つの光子も感じることが知られている」(p.186)というくだりがあって、昔勉強会でこれについての章を読んだ覚えがある(たぶんこれの51章)と思ったが、ソースが見つからなかったのでググってみた。
そしたら、二択で正答率55%くらいで弁別できるって論文が見つかった(Nature Communications 2016)。そこで引用しているもので、この世界の古典的な仕事として、Hecht et al 1942では光子5-8個が閾値、とある。
さらに調べてみたら、乾先生の「光覚の生理学的基礎」 (1981)というのを見つけた。今日はここまで。
つか、二重スリット実験を人の目をセンサーにしてやれば、同じ人の両目に分けて入れるのと、別々の人の片目ずつに入れたときとの違いを考えたらなんか意味があるのでは?とか思って調べ始めたのだけど、自分で言ってて、どういう意味があるかよくわからない。
「生命と自由」渡辺慧読了した。最終章、なんかすごいこと書いてある。熱力学第二法則が当てはまる世界でエントロピーは増える方向に行くので、ある時間よりもあとの時間のほうがよりマクロ的に見て同じ状態に収束するという意味で決定論的であり、因果的である=生命を後ろから後押しする。
いっぽうで、生物の一部はエントロピーを下げる方向に進むが、こちらはある時間よりもあとの時間でどのマクロな状態に進むかがわからないという意味で非決定論的である。ぎゃくにある時間からさかのぼってどういうマクロな状態から現在が帰結したかは予測できる。これを逆因果的過程と呼んでる。
そして人間における価値、目的論というのは(因果律が生命を後ろから後押しするのとは逆に)生命を前から引っ張る(p.196)。だから、生命というのは「自由という価値の追求」なのだと(p.198)。
これはさっき書いたカウフマンの「生物の進化における多様性を作る方向は形式化はできない」に対する答えになっている。つまり、時間を進める方向への予測をしようとするなら、それは不可能。でもその時点を引き起こした原因、価値を見つける逆因果律の方向は形式化しうる。
もちろんこれを読みながら、ずっとエントロピーのところを自由エネルギーに置き換えながら考えてた。自由エネルギー最小化の元で、同じ状態に収束していくわけではなくて、多様化してゆく(進化でも、個体でも、社会でも)ことを、渡辺慧の言う逆因果的過程、もしくは「目的論の部分的復権」ということで考えられないか?というふうに読んでいたというわけ。
「逆因果的過程」と言われると面食らうけど、期待自由エネルギーの計算で、(不確定な)将来の観察から遡って現在する行動選択を確定させるというのは、逆因果的過程でいう原因帰属的な操作をしているようにも読める。
そうしてみると、将来の観察(それはすべて列挙することが不可能な隣接可能領域)から遡って、現在の観察と行動の「尤度」が決まることのほうが本質であるように思う。
2021年06月19日
■ 研究関連ツイートまとめ(2020年後半)
受動的綜合についてZahaviが何を言ってるか調べてた。「現象学的な心」3章p.148では、ものの動きとか雑音とか、知覚的にsalientなものが触発affectionして、そのような先行的な触発に受動的に応答するreceptivityという言い方をしている。ここで引いているのはZahaviの「自己意識と他性」で
6章で「もしわれわれにそれを気に留めるように強いるならば、何らかの仕方で、対象、異質性、差異を通して目立っているにちがいない」「触発は布置の一部である何かによってつねになされ、つねに受動的に組織化され構造化された領野の内部からの触発なのである」p.187 これはサリエンシーだ。
「ベイトソンが書いているように、情報は、差異を生む差異である。そして、何かが差異を生むかどうかは、単純にそれ自体の本有的特性の事項ではなく、われわれの現行の関心—大体がわれわれの以前の経験によって影響される関心—との関係の事項である。」p.188 サリエンスも生成モデルの関数で経験依存。
じつは、本当にやりたかったのは、「感覚的なサリエンスと情動的、動機的サリエンスが、同根であるもしくは相互に強化するような関係にある」という私の主張を補強できるような議論がないか探していたのだけど、だいたいが感覚的(視覚または触覚)の例に終始している模様。
nautil.usのウォルター・ピッツに関する記事を再読してた。McCulloch and Pitts 1943での万能チューリングマシンとしての脳モデルから、Lettvin et al 1959での検証でカエルの眼が脳に伝えるのはアナログな生態学的情報だった、と夢が敗れるという書き方。
この夢が破れるシーンは「栄光なき天才たち」でコミック化してほしい。ドラマチックにしすぎだけど、脳を入出力機械として描写すれば事足りるという考えが今でも残っているとしたら、それはウォルター・ピッツの絶望を十分理解できてないからかもしれない。(<-あ、自分で書いてて書き方が気に食わない)
ところでそのような絶望が本当にあったのだろうか? 事実として残っているのは、1959論文でのWalter Pittsによると思しきパラグラフでこういうところがある。
“In short, every point is seen in definite contexts. The character of these contexts, genetically built in, is the physiological synthetic a priori. The operations found in the frog make unlikely later processes in his system of the sort described by two of us earlier…but those were adduced for the sort of form recognition which the frog does not have. This work is an outgrowth of that earlier study which set the question.”
と、カエルはform recognitionの機能を持っていないので、もともと考えていた、低レベル特徴の組み合わせによるform recognitionという考えはrejectされていない、という立場にあるように思える。
「細胞の生物物理学」7章を読んだ。 系と環境の間で熱と粒子の両方とも行き来する状況(例: 蓋のないビーカーでの化学反応)がグランドカノニカル分布(ギブス分布)で、系と環境の間で熱だけが行き来する状況のカノニカル分布(ボルツマン分布)で、両者はルジャンドル変換で繋がっている。
- グランドカノニカル分布は $p(E,N) = exp(-\beta(E-\mu N)) / 大分配関数\Xi$
- カノニカル分布は $p(E) = exp(-\beta(E)) / 分配関数Z$
分配関数 $Z$ から自由エネルギー $F$ が計算できるけど、大分配関数 $\Xi$ からグランドポテンシャル $J$ を計算してルジャンドル変換でも $F$ が計算できる。「グランドカノニカル分布とその意味」
両者がexpの中身が $-\beta(E-\mu N)$ か $-\beta(E)$ かの違いだけど、それは「微視的状態の観点からすると、熱・粒子浴の明示的な扱いを省略して化学ポテンシャル $\mu$ にその役割を負わせているということ」(「細胞の生物物理学」p.309) なんかちょっとわかってきたかも。
新型コロナにしろインフルにしろ、ウイルス感染が「見えない」「感知できない」ということが問題なので、いい加減そろそろ人間も進化して、ウイルスを感知できる表現型が現れてきたりしないもんかね?
人間に限定して考えるのは違うか。カンブリア爆発で眼が急速に進化した話を念頭に置いているのだから、ウイルスと免疫系のイタチごっこの現状において、ウイルスの方策を無効化する方法を見つけた生き物が反映して、ウイルスは使いこなせる生物にとっては有用な武器となるとかそういうかんじ。
「ウォンバットよ、お前もか! 体が光る有袋類はまだまだたくさんいることが判明」 蛍光を使って我々の知らない視覚世界を生きているのか。やっぱコロナが見えるように俺らも進化すべきだな。(<-まだ言ってる)
2021年05月16日
■ 自由エネルギー原理FEP関連メモ
FEPの説明法を見直していた。期待自由エネルギーEFEにいきなり飛ぶのは難易度が高いので、VFE = Uncertainty + Bayesian surpriseの式を使ってきた。これは不正確な方便のつもりだった。でも、行動aからサンプルされる $s$ が一意に決まるときはEFEにせずに、VFEの式でよいと自信を持った。
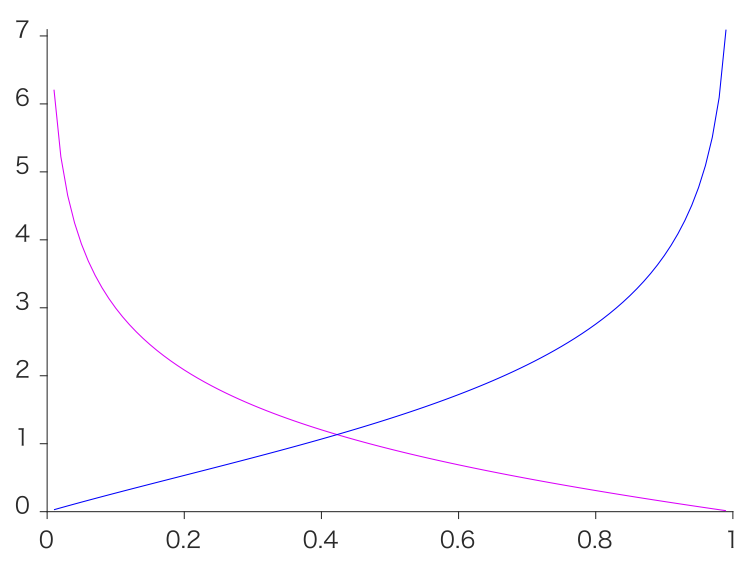
横軸に $p(s=1)$ ($s=1$:左下を見ていて蝶はぼやけてる画像; $s=2$:右上の帳を注視している画像)とおいて、posteriorは固定しておくと、$s=1$ でのUncertaintyはマゼンタ。つまり $p(s=1)<0.42$ で $s=2$ をサンプルする。つまりもし $p(s)$ を自由に動かせるなら、ありふれた $s$ を選ぶ。暗い部屋問題を再現できた。
今のはposterior $p(x|s)$ を固定して、それに $p(s)$ を掛けて生成モデル $p(x,s)$ を作っていた。このように $p(s)$ にアクセスできるなら、暗い部屋問題は起こる。しかし、実際は $p(s)$ は生成モデル $p(x,s)$ を周辺化したモデルエビデンス $p(s|m) = \sum_{x} p_m(x,s)$ なので、agentと環境との関係によって生成モデル $p_m(x,s)$ のほうが先に決まる。(ここでは、生成モデルの形を決めるパラメーターを $m$ と書いている)
そうか、「もっともありふれた刺激 $s$ 」とか言わずに、モデルエビデンス $p(s|m)$ の最大化、と言えばいいのだな。(FEPのような)BIC的なアプローチの代わりにAIC的なアプローチがありうる、って話まで持っていけそう。ぜんぶつなげて話すと、FEPの話ではなくて、ふつうにベイズ統計の授業だなこりゃ。
変分自由エネルギーFはモデルエビデンスの-logの下限(ELBO)なので、自由エネルギー最小化とは、$F_1-F_2 \approx -log(BF_1/BF_2)$ となっていて、ベイズ因子によるモデル選択と同じになる。知覚よりも行動選択の場合だと、$p_{a1}(x,s)とp_{a2}(x,s)$ という2つの生成モデルの比較がしっくりくる。
以前書いた気がするけど、「FEPはなんでも最適化主義ではないか問題」は進化における適応主義論争と形式的には同じで、「理不尽な進化」では適応主義という研究プログラム自体は否定し得ない(最適化というフレームワークで検証をすることの意義)ということでグールドが敗北したことになっているけど、カウフマンが外適応の概念を拾っているのを見て、いやエナクティヴィズム的には再考するお余地があると思った。
「理不尽な進化」も「脳がわかれば心がわかるか」も非常にデネット的なのだけど、グールド敗北で終わらない動機とか、回帰する擬似問題、といったところを残す点がこの2冊では共通してる。
この話への正確なツッコミとしては、「定式化しようと思えば定式化できる(定式化の妥当性、答えの有無に関わらず)」という意味であれば、それは無意味な言明で、それ以降の主張を支える役割を果たしてない。ここで必要な文章は端的に「制約付き最適化って重要だよね」ってことだけだと思う。
2021年05月01日
■ スチュアート・カウフマンの「生命はいかにして複雑系となったか」を読んだ
カウフマンの"Beyond Physics: The Emergence and Evolution of Life"についてのトークのスライドPDFがCERNでのトークの資料として入手できる。このスライドを使ったWisconsin Institute for Discoveryでのトークの動画。字幕はないけど。
前述のスライドのp.3 “Thus “functions” are legitimate categories in science in the historical becoming of the non-ergodic universe above the level of atoms. “Functions” do not exist in physics, which cannot discriminate “functional subsets” of causal consequences.”
ツーソン会議2018ではこれの短縮版のトークの動画がある(35分くらい)。こちらは字幕がある。こっちを視聴した。
カウフマン「生命はいかにして複雑系となったか」、さっそく立ち読みしたら、思ったより薄くて、内容は前述のスライドそのままっぽいことがわかったので購入してきた。さっそく読みだしたら、まさに私が知りたいことだった。
まとめておくと、宇宙の歴史の中でまだ可能なタンパク質の配列のすべてが作られたわけではない、そういう非エルゴード的宇宙において、進化においてどのように生物がエコロジカルニッチを見つけて進化してゆくかはあらかじめ予見することはできない。
(物理での運動方程式が初期条件と拘束条件が必要なのに対して)そもそも生物の進化においては、そのような拘束条件自体が新たに作り出されてゆく。しかもそのような「応急処置」は多様性が増すと可能性が増える。
よって、これから何が起こるかわからない(コインの裏表)どころか、何が起こりうるかすらわからない(確率測度を定式化するための標本空間が決まらない)。
このような非エルゴード的世界を記述するには物理的な説明ではなくてお話としての説明しかない(これは以前私がブログに書いた話とは逆の帰結)。だから、生命のあるこの世界は物理学を基礎とはしているけれども、それに還元されるものではないのだ、というのが結論。
ここからは私の感想だけど、物理的な説明に対するお話としての説明(その生き物にとっての「意味」を通した説明)の優位を強調するのは驚きだったけど、追求する価値はあるなと思った。これこそが人文知が必要となる、必然性の高い道筋なのではないだろうか?これはVarelaの相互的拘束よりも強い要請だ。
そんなわけで、いますごく興奮している。以前からちょっと読んでは積んでた「自己組織化と進化の論理」、あと分厚くて敬遠していた「カウフマン、生命と宇宙を語る」これらを読み進めてみようと思う。
補足: まあこれまで私は、そういう「物理を越えている」部分を情報として捉えた上で、情報は熱力学的な意味で物理とつながる、っていう理屈を想定していたのだけど、一歩立ち止まって、「一回性の世界」をもっとよく見てみよう、というのが今の心境。
追記: あと、ダーウィンの「前適応」の概念よりもグールドの「外適応」のほうを採用しているし、生態学的ニッチの意義の強調も含めて、カウフマンは非常にエナクティブな視点に立っている。進化の意義を強調しても、デネット、ドーキンス的な方向以外があり得る、このことは重要。
「カウフマン、生命と宇宙を語る」借りてきた。ざっと見、新著とそんなに大きく変わっているわけではない。あるプロセスの結果が別のプロセスの拘束条件になるって話もある。ただし、それがループとして閉じるとなってないだけ。
あと、熱力学の第4法則として生物進化を(統計的に)予測するようなものがあると「カウフマン、生命と宇宙を語る」では議論しているけど、新著ではそういう予測はあくまでも統計的なものであって、生命はbeyond physicsなのだ、という立場に立っている。新著では熱力学的な仕事(カルノーサイクル)への言及は最小限だったが、旧著ではひとつの章を当ててる。
カウフマンの"world beyond physicsで「束縛閉回路」という話が出ていた。つまり、火薬の爆発という非平衡なプロセスを砲弾を飛ばすという仕事にするためには大砲という拘束条件が必要となる。生物ではこのような拘束条件の連鎖がループして閉回路になっている、
つまり代謝の反応の連鎖で細胞壁を作って、これが拘束条件となって酵素による触媒反応が可能な程度に気質の濃度を高くする。このようにしてオートポイエーシスにおける作動的閉包をプロセスと束縛条件に分けて捉えている。
それで、カウフマンの本ではこれらが平衡から遠く離れた状態のプロセス、と読んでいたのだけど、物理化学の教科書とか読んでると、これはふつうに個々の反応はギブス自由エネルギーで取り扱えるような減少で、そのかわり、ATPとか生産物とかがその都度出入りしているけど、それは化学的平衡の速度と比べたらもっとゆっくりしたものであるということ。
そこまで見てみると、よくある「生物とは平衡からは遠く離れた状態を維持している」というフレーズが、時間の早いプロセスと遅いプロセスに分けることでかなり化学的な平衡の話で語れることであることを加味して理解しないといけないのではないかとか思った。
シュレディンガーの「生命とは何か」に出てくる「負のエントロピーを食べる」という話があって、いまそれも読み直しているけど、これの書いてある6章の注記(後日追加された)には、負のエントロピーとは正確には自由エネルギーのことであるが、一般聴衆にはエネルギーという言葉が誤解を与えるのでエントロピーという語を使ったと書いてある。そうするとここでの「秩序から秩序を作る」という話は端的にさっき書いたようなATPからADPに戻るときの大きなギブス自由エネルギーの減少ΔG1(<0)を使って、それとカップルしているリン酸化反応による秩序の形成(-ΔG1>)ΔG2(>0)を作れるという話と読める。
(もちろん遺伝子の情報がこのような回路を作るために働いていること込みで。) このような(カウフマンの言う)束縛閉回路については「エントロピーから読み解く 生物学-めぐりめぐむ わきあがる生命-」佐藤直樹に図があった。
ここはちがうか。化学反応での扱いは、系に対してその周り全体を環境として扱うから、全体としては孤立系として扱っているため、ということか。それに対して、生物でのローカルな環境(細胞の中のさらに局部)で物を見ようとすると、物質の出入りがあるから、非平衡として扱わないといけなくなる。
2021年04月30日
■ 加速主義と資本主義リアリズム
ずっと加速主義について読んでおこうと思っていたのだけど、昨晩急にその気になってきたので1日かけていろいろ読んでみた。
そもそもなんで興味を持っていたかと言うと、神経科学、意識研究の分野で、シンギュラリティーだったり、マインド・アップローディングとか、そういう話題をいくつか聞くのだけど、正直自分はぜんぜん興味持てなかった。(端的に、死ぬ前に自分の意識をアップロードしておきたいという欲望がない。) あの方向性について、自分なりに考えをまとめておきたくて、それには加速主義が関わってきそうだと思ってた。
まずはwebの資料から探した。オルタナ右翼の源流ニック・ランドと新反動主義 こちらは2018年に公開された当時にチラ見した覚えがある。今回読んでみて、なるほど、ニック・ランドと新反動主義そのものよりも、思弁的実在論との関わりに興味を持った。
さらにマーク・フィッシャーと再魔術化する世界 こちらも読んでみて、むしろマーク・フィッシャーのほうに興味が惹かれた。とくにアシッド共産主義とモリス・バーマンが出てくるあたり。
ともあれ「ニック・ランドと新反動主義 現代世界を覆う〈ダーク〉な思想 (星海社新書)」 木澤 佐登志 を読んでみた。ドゥルーズ & ガタリあたりから文体の影響を受けているようで、こういう本を読むのは久しぶりで正直面食らった。ともあれひと通り読んでみて、思弁的実在論の話がわかってよかった。あと、どうにも西欧中心主義だなと思ったが、マインド・アップローディング自体も「すべての死者を意識を蘇らせる」というキリスト教的な考えが背景にあるという記述があって、なるほどと思った。
あと出てくるサブカルがドラムンベースだったり、クトゥルーだったりホラーだったりで、自分の守備範囲からぜんぜん離れていた。Vaporwave (「リサフランク420 / 現代のコンピュー」とか猫シCorp.とか)が言及されていたところは面白かったけど、自分、いうほどvaporwave好きじゃなかったわ。
それでひきつづき、「資本主義リアリズム」 マーク フィッシャー を読んでみた。
こちらは微妙に翻訳がわかりにくいのもあるのか、読むの辛かった。そもそも「資本主義リアリズムcapitalist realism」の「リアリズム」の含意がわからない。芸術の形式としてSocialist realismってのがあって、それをもじってCapitalist realismって言葉を使ってた人がいて、それをマーク フィッシャーが拡張した、というのだけど。定義としては「ネオリベ的な資本主義によって他の選択肢はないと思わせるようなシニシズムが蔓延した状態」とあるけど、それのどこが「リアリズム」かわからなくて往生した。けっきょくのところ、政治学でいうところの、理想主義idealismに対する現実主義realismということらしくて、ネオリベ的価値観で現実見ろよってのに左翼的な運動観が勝てない、ってことなのかな、と全体を読んで察した。(そういうことが明示的に書かれてはいなかったけど、あとで調べてこの人が左翼の新しい形を模索しているという文脈を得たので。)
第3章が「資本主義とリアル」なんだけど、この「リアル」が原文を読んだら、大文字の"the Real"、つまりラカンの現実界のことで、"the Real"を資本主義が見せる’reality’(現実)が隠してしまうという話だった。このあたりは訳注がほしかった。けっきょく3章だけは原文あたって読みなおした。
本全体としては、なんかイギリスでFurther Education Collegeの教員をやっていたときの恨み言みたいな側面も多かったけど、問題となっている現在の閉塞感については日本もイギリスも同じで、共感した。
ここまできて今度は「現代思想2019年6月号 特集=加速主義」から冒頭の対談「加速主義の政治的可能性と哲学的射程」と「気をつけろ、外は砂漠が広がっている――マーク・フィッシャー私論 / 木澤佐登志」を読んだ。
対談の方はざっくり状況を掴むのに役に立った。あと、「西欧中心主義だな」みたいなのはやっぱ思うんだ、と納得した。
「マーク・フィッシャー私論」のほうは「資本主義リアリズム」から死の直前に書いていた「Acid communism序文」までの流れが書かれていて、これも面白かった。わたしのブログの過去の記事を見るとちょいちょいケン・キージーだったり、13th floor elevatorsのトミーホールの思想に触れていることからわかるように、このあたりの話題にはうるさい。
そういうわけで「Acid communism」には興味があるけど、「意識変容によって世界を変える」みたいな思想はすでに70年代に試されたわけで、たとえばそれがエサレン研究所経由で自己啓発セミナーとかになった話(「エスリンとアメリカの覚醒」は未読だけど)とか考えると、すでにすっかり「資本主義リアリズム」にとりこまれているわけで、ここに未来があるようにも思わない。「マーク・フィッシャー私論」読んだかぎり、そこは「でもやるんだよ!」精神だったのだろうか。
(なお、マインドフルネス瞑想がGoogleに採用されたり、スローライフが金持ちの道楽だったりという形で加速主義に加担してるって話は「資本主義リアリズム」の訳者の一人による記事 :「スローライフが、むしろ資本主義を「加速」させるという皮肉な現実」)で言及されてる。)
でもたぶんそれと地続きになっているんだろうと思うのは、脳科学の進歩、意識の解明によって、「自由意志とはなにか」「責任とはなにか」についての見直しがされている、そしてわれわれの意識経験がいかに多様で、変容しうるかもわかってきた。これらのことが我々の世界観が変わるうる、大きなインパクトを持っているだろうということ。そう私は思ってる。(これが「意識の科学入門」の裏テーマになってたりする。) じゃあそれはシンギュラリティーやマインド・アップローディングへの情熱とどう違うんだろう、と自分ツッコミしてみる。
…やっと本題に近づいてきたのだけど、ここで時間切れ。GWにこれ以上使う時間はないので、この話題に再訪できるのはお盆休みかそれよりも後だろうか。それではまた。
2021年04月25日
■ 「側頭」ってなんで"temporal"なの?
側頭葉temporal lobeはこめかみtempleから来ている。テンプルってまあみんなボクシング漫画で聞いたことがあって知ってるだろう(「あしたのジョー」かそれとも「はじめの一歩」か)。だから、この命名に時間temporalの意味は込められてない。
でも、いま調べてみたらtemple自体は時間から派生した言葉だそうな。
ちなみにウィキペには「こめかみ部分の髪が一番早く白髪になって歳を感じさせるから」みたいな説が引かれているが、ソース見ても納得いかない。
眼球運動とか網膜の研究者にとっては、目の内側がnasal (鼻に近い側)、外側がtemporal (こめかみに近い側)なので馴染みがある。でも日本語訳だと、nasal=鼻側はいいとして、temporal=耳側となってる。昔この訳語を決めた人の苦労が忍ばれる。
ついでに他の領域の語源も調べてみた。Frontal lobeはfrontだから前面ってことだろうと思っていたけど、実はおでこ、額(ひたい)という意味があるそうだ。こっちからきていると考えたほうがよさそう。
そうしてみると前額断 frontal planeという訳語の意味もわかる。私自身はcoronal planeという言い方のほうが馴染みがあるけど、こちらの訳語は冠状面。
Occipital, parietalも調べてみたら、これは脳葉よりも頭蓋骨から考えたほうがいいということがわかった。頭蓋骨はfrontal bone, parietal bone, temporal bone, occipital boneがあって、それぞれが脳葉lobeと対応している。じつはこめかみ部分は蝶形骨sphenoid boneが該当するけど。
そうしてみると、parietal boneのparietalが「壁」の意味であるということも、縦に長い形状から納得がいく。occipital boneは「後ろの」以上の意味は見つからなかった。ここまでやって思い出したけど、医学系の本棚に「解剖用語の語源」的な本が並んでいるから、医学生はそういうので学んでいるのか。
ついでにこういうのも見つけた。「国際解剖学用語語源辞典」山形, 健三 あいにく検索できないのがもったいない。。ダウンロード回数:38 回、とか書いてあってもったいない。
「医学用語語源対話」という連載があって、これはいいかんじ。
sagittal plane (矢状断しじょうめん)もなんでarrowなのかわからない。弓矢を持つ向き?とか適当に思っていたが、Sagittal suture (矢状縫合)が"is notched posteriorly, like an arrow, by the lambdoid suture"という説明をとりあえず見つけた。
2020年12月26日
■ R演習補講 (2腕バンディット問題を題材に)
北大CHAINのウィンタースクール2020では、集団的意思決定のモデリングについての講義と演習があります。そちらの準備用資料として「RとRStudioの基本について、強化学習の初歩的な例である2腕バンディット問題を題材に説明する」というものを学生向けに吉田が作成しました。
この資料はそちらを公開用に編集したものです。じつのところ私はRやRStudioを使うのも久しぶりで、強化学習についても素人ですが、この機会にこういう資料を自力で作って公開すれば、間違いも指摘してもらえるかもと期待して公開する次第です。
Sutton and Barto (2020) "Reinforcement Learning: An Introduction (2nd ed)"の2章が一応の元ネタですが、それをなるたけ単純化したところが特徴です。(apply関数使う前のところで時間切れになったので、一人分のデータを作るところまでしかたどり着けなかった。)
なお、コードについてですが、今回の講師のひとりである、独コンスタンツ大学の豊川航さんによるR markdownのコードと変数に使う文字は揃えていますが、コード自体は全部自分で書きました。
間違いなど発見しましたらぜひご指摘ください。よろしくお願いします。
2020年10月27日
■ 講演しました「自由エネルギー原理からエナクティビズムへ」
■ 大羽成征さん@shigepong からお話をいただきまして、全脳アーキテクチャ・イニシアティブの勉強会でCHAINの吉田と田口茂さん@ShigeruTaguchiの二人が講演をします。CHAIN協賛イベントです。
■ 私の部分は「自由エネルギー原理からエナクティビズムへ」というタイトルで、機械学習の人が興味持ってるver.3のFEPの話をするテイで、自律性に主眼をおいたver.4のFEPからオートポイエーシス、エナクティヴィズムまで連れ出すという話の流れでスライドを構成してます。
■ ここ最近の私のツイートでの
これらはみんな、この講演のための準備だったというわけです。乞うご期待。
■ 今日のWBA勉強会無事終了しました。私の講演は時間ぴったし(喋りはちょい速め)で終了。質問もいろいろ出て、懇親会も含めて楽しくお話させていただきました。
最終的に参加者は100人を超えてたはず。今回は有料イベントだったので、これにはけっこう驚いた。
こちらにスライドを置いておきます。(当日アップロードしたものは削除して、新しいものを別URLで掲載してます。SlideShareはスライドの差し替えができないので。)
当日は講演時間を考慮して削ったスライドも含めてあります。「追加」と書いてわかるようにしてます。それが無くてもわかるけど、あると親切的なスライドです。映画で言う「ディレクターズ・カット」みたいなやつ。(<-それって監督が自己満足で追加して冗長になるやつ〜)
ついでながら宣伝ですが、例年講義を行っている応用脳科学アカデミーですが、今年度は12/22に「知覚と行動と学習をつなぐ自由エネルギー原理」として開催する予定です。WBA勉強会のスライドの前半部分を膨らませてわかりやすく説明することを目指します。乞うご期待。
2020年07月27日
■ Non-trivial information closure (NTIC)は何を計算しているのか
Consciousness Club TokyoでArayaのAcer Changが"Information Closure Theory of Consciousness"というオンライン講演をしていたので観てきた。これは彼が先日Front. Psychol.に出版した"Information Closure Theory of Consciousness"という論文の内容に基づいてる。
Chang AYC, Biehl M, Yu Y and Kanai R (2020) Information Closure Theory of Consciousness. Front. Psychol. 11:1504. doi: 10.3389/fpsyg.2020.01504
この仕事については以前からその存在は知っていたけど、ちゃんと話は聞いたことがなかったので、この機会にしっかり理解しようということで参加してみたのだけど、Acerが導入部分でNTICについて丁寧に説明してくれたので、参加して正解だった。
(そのあと金井さんのトークのYouTube "On Consciousness: Information Closure and General Intelligence"も見た。こちらはさらに全体像が掴めるのでよい。)
この論文では"Non-trivial information closure (NTIC)"がゼロでないシステムが意識を持っていると主張している。論文の中での議論にはいろいろ言いたいことがあるのだけど(盲視の扱いとか、SMCの扱いとか)、まずはその前に"Non-trivial information closure (NTIC)"という概念についてしっかり知るべきだろう。
ということでNTICの概念を最初に提唱したBertschinger et al. (2006)の"Information and closure in systems theory"を読んできた。このあたりについてまとめてスライドを作ったので、slideshareに上げておいた。
結論としては、NTICというのは、もし時系列データに適応せずに単に3変数データに使ってみた場合には、MI(X;Y;Z) と等価だった。Bertschinger et al. (2006)では、それをsystemとenvironmentの関係に当てはめた上で、E1->S_2のように時間遅れがあるものが感覚入力による影響で、E1->S_1のような同時の影響がモデリングによるものだ、という意味づけをしていた。ここが肝であるようだ。
さらにこの同時の影響については、Martinの論文にもあったように、粗視化と捉える視点もあった。そういうわけで、まだまだ本質には辿り着けてない。
NTICがやっていることとマルコフブランケットがやっていることはどちらとも条件付き独立に関わっていてよく似ている。金井さんのトークでは両者の関係についても言及していたが、このへんについても言いたいことはあるが、まだもう少し準備が必要そうだ。
ともあれ、今回このスライドを作るにあたって、自力で式変形して、エントロピーとか相互情報量とかの基本の再勉強になったのでよかった。
2020年02月25日
■ 熱力学の再勉強中。
ここ最近Fristonの自由エネルギー原理FEPについて自分で勉強したり、各地で講演したり、生理研でチュートリアル・ワークショップを開催したり、とさまざまな活動をしてきた。
FEPでは、ベイズの法則による事後分布の計算を変分ベイズでやるために情報理論で言うところの「変分自由エネルギー」を最小化する。(機械学習ではELBO evidence lower boundという呼び名のほうが浸透しているだろう。)
FEPが興味深いのは、環境と相互作用するagentの内部状態がどのように時間発展してゆくかを決定づける変分原理としての役目を果たすかもしれない、という点だ。この点がFEPの特異かつ怪しい部分であって、もしFEPがただの「ベイズの事後分布の簡便な計算法」であるならば自由エネルギー「原理」と銘打つ根拠がない。
先日京大・生命科学の本田直樹さんが企画した国際シンポジウム "COMPUTATIONAL PRINCIPLES IN ACTIVE PERCEPTION AND REINFORCEMENT LEARNING IN THE BRAIN"に参加して、ドイツ ウルム大学のDaniel Braunの連続講演を聞いてきた。
彼は物理的な意味での(ヘルムホルツの)自由エネルギーの最小化を発想の元にして、知覚と行動の統一理論として自由エネルギー最小化を提唱している。Daniel Braunの自由エネルギー原理も発想は違うけど結果としてFristonのFEPと同じものを最小化している。以下の2つの論文が基本理論で、ここから様々な応用をしている。
- Ortega PA, Braun DA (2013) Thermodynamics as a theory of decision-making with information-processing costs. Proc R Soc A 469: 2 LINK
- Genewein T, Leibfried F, Grau-Moya J and Braun DA (2015) Bounded Rationality, Abstraction, and Hierarchical Decision-Making: An Information-Theoretic Optimality Principle. Front. Robot. AI 2:27 LINK
そういうわけでDaniel Braunの論文も読んでいるところで、そのう ちまとめておきたい。それはまた今度。
でもって今回の本題なのだけど、そんなこんなで、そろそろ情報理論での変分自由エネルギー最小化と熱力学、統計力学でのヘルムホルツ自由エネルギーの最小化との対応付け(がそもそもあるのかを含めて)をちゃんと理解しておく必要があるなと思った。
いや、いちおう大学1,2年のときに熱力学、統計力学の講義があって、試験もパスしてはいるのだけど、一夜漬けでぜんぜんダメな成績だった記憶がある。そのあとブルーバックスとか読んではいたのだけど、熱力学を歴史を追って説明するというストーリーで毎度挫折していた。そこでそれとは異なるアプローチで、しかも定評のある「熱力学 ― 現代的な視点から」田崎晴明 培風館 を読むことにした。
この本では、(じつのところ見えない存在である)熱Qの概念からスタートするのではなくて、(計測可能な)仕事Wの概念からスタートして、熱力学を再構成するという話の運びになっている、というところまでは以前調べて知ってた。それでこの三連休を使って1章から7章のヘルムホルツ自由エネルギーの最小化までとにかく読み進めた。
でもって読みっぱなしだと頭に残らないので、こういうときはとったノートをまとめて公開できるように形を整えてみるのがよいだろう(ブログのネタにもなるし)、と考えてまとめノートを作ってみた。まずこちらがA4一枚に収まるように作成したチートシート的なもの。(画像からPDFにリンクしてある。)
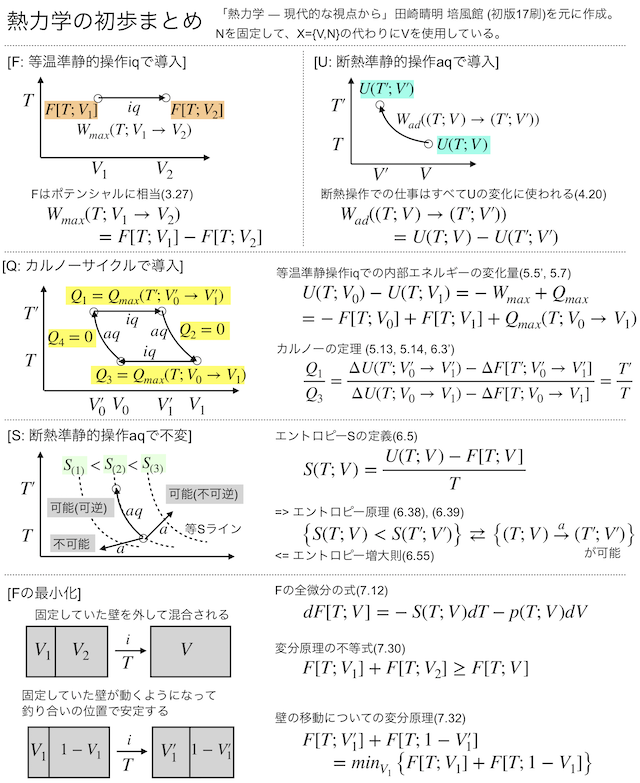
もうひとつが9ページ分でまとめた、よりノートらしいノートになってます。こちらはSlideShareにアップしておきました。
この分量を作るだけでもけっこうな時間がかかった。まだ本文を飛ばしながら読んだだけで、繰り返し読んで、例題を解いてゆくことでさらに理解が深まるわけでまだ先は遠い。熱力学自体の勉強としても相転移くらいまでは理解しておきたいし(臨界現象は生物の理論的にも重要そう)。
やってみて再確認できたのは、けっきょくのところ熱力学でのエントロピー、自由エネルギー、といったものは平衡状態でのみ定義可能なものであって(途中の過程自体は平衡であり続ける必要はないけど)、それを生物のような物質の出入りがありながら構造を保ちつづけている非平衡定常状態NESSに単純に当てはめるわけにはいかない。非平衡でのエントロピーをどう定義するか自体が現在でも課題であり、このあたりの進捗をフォローできるように勉強しておく必要があると思った。(FristonのParticular Physics論文でもNESSとマルコフブランケットから話が始まった。)
そして情報理論での変分自由エネルギー最小化との対応付けという意味では本丸は統計力学の方なのだけど、こちらについてはここさいきん渡辺澄夫「ベイズ統計の理論と方法」を読み進めたので、また別の記事を作成できたらと考えている。そちらはまたの機会に。
2020年01月23日
■ 北海道大学 人間知・脳・AI研究教育センター(CHAIN)の博士研究員の募集を開始しました
1月から北海道大学 人間知・脳・AI研究教育センター(CHAIN)に特任准教授として異動しました。1月から3月の間は札幌と岡崎の往復生活です。
今は仮の住まいで文学部の中に居室を置かせてもらってます。CHAINじたいは文学部とは独立した組織ではあるのだけど。
札幌は何年ぶりかの暖冬で、ここしばらくは1月なのに道路に雪がないという異常な状態でしたが、今週は雪が降って真っ白になってます。

さて本題ですが、吉田ラボでマーモセットを対象とした神経科学的な研究をメインでやっていただく博士研究員を1名募集しています。所属は北海道大学の人間知・脳・AI研究教育センターとなります。研究期間は年ごとの更新ですが、最長で革新脳が終了する2024年3月までです。
募集要項が北海道大学の採用情報のところに掲載されました。募集要項のPDFファイル。明日にはJREC-INでも配信されるはずです。募集方法などの詳細については、吉田のCHAINでのwebサイトにある「博士研究員募集のページ」をごらんください。
もし札幌近郊にお住まいの方で興味のある方がおられましたらお早めにご連絡ください。今度2/6に北海道大学でセミナーを行いますので、その前後で詳しいお話をすることができます。
もちろんこの機会に札幌行くぞって方、留学中で帰国予定の方も大歓迎。博士号取得見込みで4月からの行き先を探している方は応募を検討してみてください。よろしくお願いします。
2019年10月08日
■ 「時間は存在しない」、エントロピー、ギブスのパラドックス(3/3)
前回からのつづき。
「エントロピーが観察者に依存している、もっと正確にはどういう熱力学的状態を問題とするかに依存するという話はJaynesとかいろいろある。」と書いたけど、その部分についてまとめる。
E. T. Jayesは"Gibbs vs Boltzmann Entropies (1964)"においてこう書いてる。
thermodynamics knows of no such notion as the “entropy of a physical system.” Thermodynamics does have the concept of the entropy of a thermodynamic system; but a given physical system corresponds to many different thermodynamic systems.
From this we see that entropy is an anthropomorphic concept, not only in the well-known statistical sense that it measures the extent of human ignorance as to the microstate. Even at the purely phenomenological level, entropy is an anthropomorphic concept. For it is a property, not of the physical system, but of the particular experiments you or I choose to perform on it.
エントロピーは「ある物理系」について決まるものではなくて、「熱力学的システム」ごとに決まる。
たとえば「ロッシェル塩(酒石酸カリウムナトリウム)の結晶」について実験を行うとき、実験1では温度と圧力が興味の対象なので、エントロピーは となる。実験2ではひずみと分極が興味の対象なので、エントロピーは となる。
つまり、「ロッシェル塩の結晶固有のエントロピー」というものがあるのではなくて、熱力学的状態を定義するパラメーターを規定するとエントロピーも規定される。
For example, I have been asked several times whether, in my opinion, a biological system, say a cat, which converts inanimate food into a highly organized structure and behavior, represents a violation of the second law. The answer I always give is that, until we specify the set of parameters which define the thermodynamic state of the cat, no definite question has been asked!
生物(たとえばネコ)は第二法則をviolateするか?というよくある質問に対しても、「そのネコについての熱力学的状態を定義するパラメーターを規定しないかぎり、それは(意味のある)質問になってない」というのが答えだ。
さきほどの粗視化の問題についてWikipediaで調べてみると、MaxEnt thermodynamicsの項目で出てくる。E. T. JayesはMaxEnt thermodynamicsの創始者だ。あとどうやらこのラインの考え方は、QBism(量子ベイズ)にも繋がるらしい。
QBism批判でも人間原理的な主観確率が問題になっているようだけど、上記のロヴェッリが書いたような、あくまでもローカルな相互作用ごとにエントロピーが決まるということならば、人間原理にならないのではないかと思うのだけど、素人なのでまた勉強しながら考える。
こういう話題でよく出てくるのが、「ギブスのパラドックス」というものだ。これもまとめておく。
(状況1) 気体Aの入った箱と気体Bの入った箱とをくっつけて繋げるとAとBが混ざる。当然エントロピーも増大する。(状況2) 気体Aの入った箱と同じ気体Aの入った別の箱をくっつけて繋げる。状況2ではおなじAとAなのでエントロピーは増大しないはず。でも計算上は状況1と同じだからエントロピーが増大してしまう。
このスライド(PDF)がわかりやすかった。
この問題の解決法としては、量子力学においては同種粒子が互いに区別できないから、配置数 を多く数えすぎているために起こる、と考える。よって分配関数 を で割って補正する。
量子力学的説明に訴えなくても、統計力学的なエントロピーに示量性をもたせるように補正することで解決できる。つまり、熱力学的なエントロピー と統計力学的なエントロピー があって、両者の差を決めるambiguity function がある。
を統計力学的なエントロピーに示量性がなりたつように決めてやると、
この の項が、量子力学的説明で粒子を区別できないときに で割る補正をすることと同じ役割を果たしている。(Sterlingのapproximationより )
「熱力学―現代的な視点から」の田崎晴明氏のサイトにギブスのパラドックスについての記載がある。6/15/2000(木) ここでリンクされている「模範解答」の部分。さらに6/19/2000(月)の記事では、上記のJaynes 1965での「異なる熱力学的状態」と同様なことを書いてる:
熱力学の構造というのは、 マクロな視点を指定したときにはじめて「現れて」くるものなのだ。 マクロな視点として、
- パチンコ玉をいっさい区別しない
- 「○×会館」の玉か「パチンコランド××」の玉かだけは区別する
- すべての玉に番号を振ってきっちり区別する
といった幾通りもの視点が可能であり、 それに応じてもっとも便利な熱力学的な構造を選ぶのがよい。
Janyesに加えて、ギブスのパラドックスにおけるエントロピーの意義についてなんどか引用されているのを見かけたのがvan Kampen (1984)で、原文にあたるとこう書いてる。
Thus the paradox is resolved by replacing the Platonic idea of entropy with an operational definition. Quantum mechanics has no bearing on the question.
The question is not whether the particles are identical in eyes of God, but merely in the eyes of the beholder.
N. G. van Kampen, (1984) “The Gibbs paradox,” in Essays in Theoretical Physics, edited by W. E. Parry (Pergamon, Oxford), page 303-312 Google Booksのプレビュー: https://books.google.co.jp/books?id=75Y3BQAAQBAJ&pg=PA303
こちらでも、エントロピーがエネルギーのような物質の特性ではないということが強調されてる。
あともうひとつ、今回調べていて面白かったのが、非平衡統計力学とギブスのパラドックスとの関係について。「微小熱力学系におけるGibbsのパラドックス」 この内容についてはPhys. Rev. Lett. 118, 060601に出版されている。arXiv版もあり。
ギブスのパラドックスのうち、熱力学と統計力学との間での整合性の問題(GP-III)の解決法(示量性に訴える)が微小熱力学系では成り立たないのをどうするか、というのがこの論文の問題。示量性の代わりに非平衡統計力学でのゆらぎの定理と絶対不可逆性から前述の補正項 を決めることができて、
となる、とのこと。情報熱力学を勉強したいと思っていたので、ひとつとっかかりができてよかった。
そういうわけで、最初の疑問からずいぶん遠くまで来たが、途中のギャップを埋めるためには、熱力学、統計力学、情報理論を勉強する必要がある。先は遠いが、まあ永遠に生きるつもりで勉強はしてゆくことにしよう。
2019年10月07日
■ 「時間は存在しない」、エントロピー、ギブスのパラドックス(2/3)
前回からの続き。
このあたりでいい加減熱力学とかボルツマンの原理とか勉強しないといけないなと悟った。とはいえ教科書でじっくり勉強している時間もないので、ブルーバックスとか入門書を読んで最小限まとめておく。あくまでこれはわたし用のノート。こんなの学部生以来だから30年ぶりかも。
「高校数学でわかるボルツマンの原理」を元にしてまとめておく。
[Maxwell-Boltzmann分布(p.156)]
基本セッティング:
- 個々の気体分子のとりうるエネルギー状態:
- ぞれぞれのエネルギー状態の気体分子の数:
- 分子の総数
- 総エネルギー
例: 分子の数が4, 取りうるエネルギー状態が4 (等間隔)
- 総エネルギー の場合に可能な気体分子の数の組み合わせ:
- それぞれの組み合わせで取りうる場合の数
- 同様にしてすべての組み合わせで
- この 通りが同じ確率で分布する(等確率の原理)
- よって一番起こりやすい分布は
[ が大きいときの一般化(p.166)]
- で微分してmaxとなる を決める
- の代わりに をそれぞれの で偏微分して になる を見つける。
- スターリングの公式 より
- 偏微分して とおくと
- 分子の総数 は一定なので による偏微分はゼロ
- エネルギーの総数 は一定なので による偏微分はゼロ
- 以上の3つの式(1), (2), (3)を使った連立方程式を、ラグランジュの未定乗数法を使って解く。
- (4)が や に依存せずに成立する条件は
- よって各エネルギー状態の分子の分布は を入れると、
- を消すために、分子数の総和 を入れて
- 式(5), (6)より を消すと
- この分母の が分配関数。 に依存しない。
- つまり、エネルギー総和 のときに、エネルギーが高い状態 になるほど、存在確率 は低くなる。
- 同様に総エネルギーの式も表現できる。
[ボルツマンの原理の導出(p.196)]
エントロピー (熱力学)と場合の数 (統計力学)を繋ぐ。
Step 1: ヘルムホルツの自由エネルギー を統計力学的な で表現する
- を温度 で微分(体積 は一定)
- 式(9)を用いて、 を温度 で微分してから、式(7)の を代入
Step 2: 微分方程式(10)を解く
Step 3: 式(9)の に式(11)を代入する
[ボルツマンの原理の右辺と左辺の関係]
Arieh Ben-Naim (2007) Entropy Demystified, the Second Law of Thermodynamics Reduced to Plain Common Sense, World Scientific, Singapore こちらのページに1章と8章のプレビュー用PDFあり。
この本の8章でボルツマンの原理の右辺と左辺の関係について言及してた。
左辺の は熱力学的エントロピーだから単位は で、右辺の は場合の数だから無次元。両者を合わせるためにボルツマン定数 を掛けてある。だから、熱力学的エントロピーと場合の数の対数(=情報)は別物であるわけだけど、
Recall that temperature was defined earlier than entropy and earlier than the kinetic theory of heat (…) Once the identification of temperature as a measure of the average kinetic energy of the atoms had been confirmed and accepted, there was no reason to keep the old units of K. (p.204-205)
そもそも温度 は歴史的経緯から単位 がつけられたけど、 となるように温度を再定義してやれば、そのような温度 で計算された熱力学的エントロピー は無次元となる。
よってボルツマンの原理の左辺も右辺も無次元でどちらも情報を表しているのだ、と書いてる。
同じ問題について「エントロピーを巡る冒険」(鈴木炎)の3章にも言及があった。ボルツマンの原理の式の が場合の数であるけれども、熱力学の公式でのエントロピーの式(5)のほうが難解だ。
問題はエントロピーよりも温度の方だと。
<温度>について考えを巡らせ、その本質について思い悩んでみると、一つわかることがある。「熱い」「冷たい」という感覚につきまとうのは、常に「触ってみる」という行為なのだ(…)触れることで、手に熱絵エネルギーが流れ込んできた時、「熱い」と感じる。熱エネルギーが流れ出すと「冷たい」。かたや0度の水と0度の氷は、くっつけたときに熱がどちらの方向へも流れない。だからこそ、われわれは両者の温度が等しいことを知るのである。(…)だが、これは、熱力学第二法則、エントロピーの法則そのものではないか!(p.125-126)
だから、あなたがいま、式(5)を理解したいと叫ぶとき、そこで問われているのは<エントロピー>の意味ではない。<温度>の真の意味が問われているのである。すなわち式(5)が語るものは、<温度>とは何か、という疑問への最終解答ー温度の<定義>なのである。(p.127)
なるほど!式(5)の をよくみる書き方 に変えておくと、
これって温度の定義式みたいだなって思った。
次回に続く。
2019年10月06日
■ 「時間は存在しない」、エントロピー、ギブスのパラドックス(1/3)
「時間は存在しない」カルロ・ロヴェッリ、読了した。すげー面白かった。まさにこれが今知りたいことだった。エントロピーは相対的な概念(速度が観察者と対象との間の相対的な速度であるのと同じ)ではあるが、あくまで相互作用する系の間で規定されるものであり、心的過程を前提とする必要はない、とスッキリと納得できた。
時間の矢とエントロピー増大の法則を関連付ける、という話は何度か聞いたことはあるけれど、それを時間と空間のない「永遠主義」的な立場から、宇宙の中でたまたまエントロピーの低い系に我々がいて、エントロピーの増大を時の流れとして経験する、というストーリーは、批判的に読まなければならないだろうけど、いままででいちばん意味がある考え方だと思った。
あと別のラインでエントロピーのことがずっと気になってた。情報自由エネルギー原理FEPや情報統合理論IITについて考えるにあたってずっと気になっていたのは、情報やエントロピーが環境に実在するように扱われているけれども、観察者に依存しているものではないの?ということだった。
しかもこれはFEPで扱われているような生物の認識の問題で使われる情報理論的なエントロピーの話だけではなくて、物理的な意味でのエントロピーでも関わってくるらしい。
エントロピーはわたしたちが何を識別しないかによって変わってくる。なぜならそれは、私達には区別できない配置の数で決まるからだ。まったく同じミクロな配置のエントロピーが、あるレベルのぼやけでは高くなり、別のレベルのぼやけでは低くなる。だからといって、このぼやけは人間の精神が生み出したものではなく、あくまで実際に存在する物理的な相互作用によって決まる。エントロピーは恣意的でもなければ主観的な量でもなく、速度のような相対的な量なのだ。p.144
速度とは、何かほかのものに対する性質、すなわち相対的な量なのである。エントロピーについても同じことがいえて、BにとってのAのエントロピーとは、AとBの間の物理的な相互作用では区別されないAの状態の数なのだ。p.144-145
ここで言っている配置の数というのは、ボルツマンの原理の話をしている。ボルツマンの原理では
熱力学的なエントロピー と統計力学的な可能な状態での配置の数 とを結びつけているわけだけど、この のこと。
エントロピーが観察者に依存している、もっと正確にはどういう熱力学的状態を問題とするかに依存するという話はJaynesとかいろいろある。これについては今回調べてまとめたのであとで書く。でもそれを「物理的な相互作用」に依存するという言い方をしているのは初めて見た。そしてそれはすごく納得いった。
たとえば、気体Aと気体Bがピストンを押しあうという相互作用においては、圧力と温度というマクロなパラメーターしか効いてこないからこそ個々の分子の位置やエネルギー情報が無視されている。いっぽうで、気体Aと気体Bが半透膜で仕切られていて、分子Aだけが通り抜けられるという状況では、気体Aと気体Bの違いは無視できない。そしてここには観察者は必要がない。
あとここでの「ぼやけ」というのは粗視化のことを言ってる。Wikipediaの粗視化の項にこの件について書かれている。つまり、エントロピー増大の法則というのは、粗視化が必要なときだけ起きる。粗視化とエントロピー増大の法則の関係については、stack exchangeの回答にあったこの図がイメージしやすかった。(ところで「無知であること」と「粗視化」の違いが私にはまだ明確でないのだけど、あくまでもリウヴィル方程式が出てくるような場面でのみ粗視化の概念が必要となると理解している。)
過去と未来の違いはすべて、かつてこの世界のエントロピーが低かったという事実に起因しているらしい。(p.142)
小さな系Sにとっては、熱時間の流れ全体から見たエントロピーは一般に高いまま推移し、せいぜい上下に揺らぐくらいである…ところが、わたしたちがたまたま暮らしている途方も無く広大なこの宇宙にある無数の小さな系Sのなかにはいくつか特別な系があって、そこではエントロピーの変動によって、たまたま熱時間の流れの2つある端の片方におけるエントロピーが低くなっている。これらの系Sにとっては、エントロピーの変動は対象でなく、増大する。そしてわたしたちは、この増大は時の流れとして経験する。つまり特別なのは初期の宇宙の状態ではなく、わたしたちが属している小さな系Sなのだ。(p.154-155)
これめちゃ面白いんだけど、「我々にとって」エントロピーが低いということがどういうことなのかに依存している。上記のように、エントロピーを考えるのに、われわれ主観的な観察者を考える人間原理は不要で、あくまで相互作用の問題なので。
わたしたちとこの世界の残りの部分が特殊な相互作用をしているからこそ宇宙が始まったときのエントロピーが低かった、というのはどういうことなのだろう?(p.146)
ここの説明でトランプの例が紹介されている(p.147)。つまり、12枚のトランプがあって、6枚の赤の束に6枚の黒の束を重ねてシャッフルするとだんだんバラバラになってゆく。つまりエントロピーの低い状態からエントロピーの高い状態になった。一方で、12枚のトランプをすべてガン牌(麻雀用語)できる場合は、はじめの段階でスペード6とかハートAとか12枚すべてを知っているから、シャッフルしても「バラバラ」にならない。つまりエントロピーは変わらない。
この例は「宇宙が始まったときのエントロピーが低かった」ことの説明のところに来ているけど、むしろ上記の、エントロピーとは我々が区別できない配置の数に依存する、の説明の方が向いているように思ったけど。
そんなわけで、もうすこし深掘りしてみることにしよう。ループ量子重力理論じたいを学ぶつもりはないけど、ロヴェッリの論考についてはarXivにプレプリントがあるとのことなので、そのあたりを読んでみようと思う。
- arXiv 2016 “Meaning = Information + Evolution”
- arXiv 2013 “Relative information at the foundation of physics”
- arXiv 2015 “Is Time’s Arrow Perspectival?”
まずEdgeの文章"Relative information"が短いので読んでみた。でもこれはあまりに一般的に情報のことしか書いてないので面食らう。
じつのところRelative informationというのはなんのことか、"Meaning = Information + Evolution"を読んでみた。ここでRelative informationを定義しているのだけど、系Aと系Bがあったとして、 と それぞれのエントロピーを計算して差をとったものって書いてあるけど、いやそれってふつうに総合情報量の定義 そのものなんじゃないのか?ここでいっきょにわからなくなった。積分はLiouville Measureでとるって書いてあるから、ここに粗視化が出てくるのだろうとは思うのだけど。
今日はここまで。明日に続く。
P.S. ちなみに"Meaning = Information + Evolution"の後半に出てくるKolchinsky-Wolpertによるsemantic informationの定義というのは面白い。最終的に出版されたのはInterface Focus. 2018
ここに動画あり:"Observers as Systems that Acquire Information to Stay out of Equilibrium by David Wolpert" スライド(PDF) もあり。スライドの最後のページを見てもらえばわかる。FEPよりもこっちのほうを最大化していると考えたほうがよいかも。こっちもいま読んでいるので、そのうちまとめたい。
P.P.S. David Wolpertってサンタフェ研究所の「ノーフリーランチ定理」の人。この動画はFoundational Questions InstituteのFQXi2016のトークというもので、ジュリオ・トノーニや大泉さんもトークしている。
2019年09月09日
■ 「圏論の道案内」のはじめの方を読みながら考えたこと(つづき)
「「圏論の道案内」のはじめの方を読みながら考えたこと」のつづき。
圏論の初学者としてひとつ面食らったのが、恒等射のときに同じ対象が2回出てくること。私はベイズの因果グラフの表現に慣れているから、同じものが2ヶ所に出てくるのは違和感なのだけど、圏論では対象はあくまでも射の性質を規定するものだから、同じ対象が別のところに出てきてもよい。これは初学者にとっては自明でない前提だ。

つまり、ベイズの因果グラフというのは、あくまでも対象のほうが先にあって、それらのあいだの因果関係として有向グラフの矢印が付加されるものなので、こちらでは同じ対象が別の場所に2度出てきてはおかしい。当たり前だと思うかもしれないけど、こうやって言語化してみて、初めて納得がいく。
こうやって書いてみると、圏論で同じ対象が複数出てきてもよいのは、あくまでも射が主役であって、それの性質を規定するものとして域と余域があるからで、恒等射では A=dom(f) かつ A=cod(f) という表現も、そこまで考えると(私にとってはやっと)納得いく。
なるほど!これは素晴らしい。因果グラフが実体論を引きずっているのに対して、圏論はあくまで射・矢印がすべてなので、object は二の次。矢印の方からdomain とcodomainが引き出されてくる。表面的にはわかっていたが、吉田式の噛み砕き方で理解が深まった。 https://t.co/LtdCZJy1ty— @ShigeruTaguchi September 8, 2019
そうです、さらにいえば、その下のf1とf2が同じかどうかは可換であるかどうかで決まるので、射の方すら、空間的に同じところを占めているかでは同一性を決めてない。この意味でも実体論から離れてる。たぶん。
うん、なるほど、射の実体論さえ抑止するという点で、圏論は本当に徹底してますね。西郷・田口本でも、数学は「固定」したら終わりで、むしろ徹底して「自由」でなければならない、という話をしています。「この特定の三角形」から離れられなければ、幾何学もありえない。こうした方向性の権化が圏論— @ShigeruTaguchi September 8, 2019
2019年09月08日
■ 「圏論の道案内」のはじめの方を読みながら考えたこと
西郷さんの「圏論の道案内」 を読みはじめた。とても面白いが、ハマりすぎないように、家からの行き帰りの時間だけに限って読んでる。
p.28で圏論の初学者が「任意の対象A,Bについて、AからBへの射はないか、あっても一つだけだ」と誤解しがちであることについて書いている。
私は圏論の初学者として、誤解する気持ちはわかる。圏の定義2.1(p.20)に戻ると「どんな射についても域と余域がただ一つ存在する」(要約)とあるが、読者(私)はここをさらっと読んでしまうので「この文章がいったいどういう可能性を排除しているのか」に心が及んでいないのだ。
文章がなにか意味のあることを言うときには、なんらか他の可能性を排除している。たとえば「岡崎は暑い」という文章は「岡崎は暑くない」というあり得たかもしれない他の可能性を排除している。
だから圏の定義2.1の話では、即座にどういう可能性を排除しているのかを例示してやることで、読者は圏の定義2.1を自分の中でパラフレーズして、意味のあるものとすることができるようになる。つか私はそういう操作ができないと「理解」できた気がしない。
さてそれでは圏の定義2.1はどういう可能性を排除しているか、私が好きな「プログラム学習」方式で例示してみたい。「プログラム学習」の例としては「Scheme手習い」がある。Google Booksに本文のいちばんはじめの部分(p.3)があるので、そこを見てもらうと、だいたい言いたいことが分かると思う。
では「Scheme手習い」の方式を踏襲して、定義2.1をコッテリ確認してゆく例を作ってみよう。私としては、こんなふうに手元にノートを取ることができれば、自分で理解できたなという実感が持てる。

というわけで、これは圏論本についての批評ではなくて、私はどのようにパラフレーズできたら理解できたことになると思っているか、についての話だった。
FEPの説明についてもそういうことを考えた。私のFEP解説での蝶と蛾の例にしても、state * observation * actionを最小限の組み合わせで操作してはじめて(私自身が)わかった気になった。
だから、強化学習の教科書でいきなりベルマン方程式が出てくるのが納得いかない。でも人によっては写経のように頭から読み下してしまえばそれで理解できるらしい。じつのところ私はそういう理解を疑っているのだけど、そうではなくて、そのように理解できる人こそがその分野に進むという生存バイアスの結果なのかもしれない。ともあれ私の理解の仕方が特殊である可能性を排除できないので、ここでは例示して理解する方法の有効性だけを主張するのに留めておきたい。
強化学習について勉強していたときに同様なことを考えてtwitterに書いたことがある(Twilog20190813)。そのとき書いたことについてもここでまとめておく。
TDアルゴリズムからQ-learningまでについての動画をいろいろ見てた。この動画がいちばんわかりやすかった。まず行動選択無し、報酬の不確定性なしで説明してから、徐々に要素を追加してゆく。スゲー親切。パート1から3まであるけど合計42分で終わる。視聴数少ないのが不思議。
Grid world問題がどれもこれも複雑な3 * 4マスの例を使って説明しようとするのが、私にはぜんぜん訳わからない。1 * 3マスでよくね?
あと、とりあえずベルマン方程式書いて始める動画がほとんどで、ぜんぜん訳がわからない。まずクソかんたんな状況(1 * 3とか)を説明して、それから帰納的に式導入すればいいのに。これって情報科学勉強している学生にとっては、まず式から入るほうがわかりやすいとかあるのか?
1x3だと簡単すぎて当たり前になっちゃわないかなぁって。真ん中の状態だけpolicyが効くし。でもline state の例はsuttonの教科書にもあったような。— @KazuSamejima August 12, 2019
@KazuSamejima 「簡単すぎて当たり前」な問題を解くところを見せて、そこから読者に帰納的にベルマン方程式を「発見させる」ように進めたらどうかということなのです。グリッドワールドの説明は、ベルマン方程式を与えておいて、意味のある最小限のリアルな問題を解けることを見せることに注力しているようだけど。
そうですね。ベルマン方程式は方策を決めたときの価値関数が満たすべき式として導出してしまって、ダイナミックプログラミングのアルゴリズムがどう動くかのデモとして格子空間を使う、説明がほとんど。帰納的に発見させることができるのなら、そちらのほうが自然ですね。うまいデモはわからないけど。— @KazuSamejima August 13, 2019
以前も書いたけど、プログラミング言語でのfor文だって、まず
ii=1; disp(ii); ii=2; disp(ii); ... ii=100; disp(ii);
(matlabでの例)みたいなことをやらせて、いい加減馬鹿らしくなったところでfor文を導入してほしかった。どうやらそういうことをしなくても分かる人は分かるようだが。
以前書いたけど、私が新しいことを学ぶときに最適だと思うのは「プログラム学習 有機合成化学」のような短い設問形式でステップを細かく刻みながら学ぶやつ。「Scheme手習い」も同系統だけど「風変わり」とか「クセがある」とか書かれている。
この話に私がこだわっているのは、べつにいまある解説とかに文句をつけたいとかではない。数物系特有のアタマの使い方というのがあって、どうやら私はそれから弾かれているようなのだけど、向いていようが向いてまいが自分で知りたいので、なんか流れに逆らうように感じるということだ。
同じ話は哲学の人の話を聞くときにも感じていて、学会とかで原稿をそのまま読み出すというのに遭遇して異分野の人間として面食らうのだけれど、どうやらあの分野の人達は、ゼミの文献購読でひたすらそういうことをし続けてきているので、苦に思わないようだ。
私にとって哲学のような概念を使う話は、複数のargumentがマインドマップのように矢印でつながっているものだと思っていた。だからそれを人間が読める形にするために(人間は一次元的に文字を追って読むことしかできない)文章という形式に変換して、聞く側はそれをデコードしてマインドマップに戻しているのかと思っていたのだがそうではないらしい。
これもいろんなタイプがいます。文字を文字のままで理解するほうが向いている人もいれば、言語で言われたことをイメージ化して理解している人もいます。私は後者。抽象的な概念も、図に描けない変な形をして見えたり、個性的な「味」があったりします。— @ShigeruTaguchi August 13, 2019
しかしいずれにせよ、哲学業界の人は、それぞれの思考の特性をもとにしながら、しゃべりだけで抽象概念やargument をいじくる訓練を受けてきているのは確かですね。— @ShigeruTaguchi August 13, 2019
ちなみに、頭の中の変なイメージは、2次元, 3次元の図形にうまく落とし込めないので、言語的表現の方が相性がよかったりします。言語はわりとアクロバティックなことも可能で、「入ってくる前に出て行く」(レヴィナス)とか、色々変なことが言えます。— @ShigeruTaguchi August 13, 2019
同様に、数物系の人たちは、式が与えられると、たとえばベルマン方程式に具体的な数字を入れて実際に動いている様子まで戻してやらなくても、抽象的な式のままで操作ができるということなのだろうか。ここまで書いて気づいたけど、どうやら自分は抽象的なものをそのまま操作することが不得意なようだ。
(この話の続きの部分を別エントリとして作成しました。こちら:「「圏論の道案内」のはじめの方を読みながら考えたこと(つづき)」)
2019年07月19日
■ 一回性の現象と統計的な現象との連続性に注目してみる
カウフマンの新著"A World Beyond Physics"のレビュー"The new physics needed to probe the origins of life"が面白かった。
この文の中でカウフマンの"the nonergodic world"という概念に触れていて、タンパク質として可能な、200の20乗の組み合わせのうち、まだほんの一部しかこの宇宙の歴史上に現れてない、つまりエルゴード性が成り立たないようなところで、進化という一回性/歴史が起きているようなのだ。
これを見て考えるに、このような来歴に縛られた発展過程を見ているというのは、進化でどのように種が現れたかとかいう話だけでなく、生命の発生においてもそうであるらしい。
そう考えてみるとなんだかわかってきたのだけど、この「non-ergodic/historic/一回性」であることと「ergodic/statistical」であることって、スケールに依存した相対的なことなんだな。
進化を一回性の現象として見ているのは、われわれにはこの地球での進化しか見えてないからであって、(おそらく存在するであろう)宇宙の他の星すべてでの生命と進化をまとめて見る視点からは、進化すらも統計的な理論として捉えることが可能になる。でもそれができないから我々は進化を、その履歴に影響される現象としてしか理解することができない。
もし我々が分子のサイズで周りの気体分子を観察したなら、それはある分子がぶつかってそれがこっちに向かって跳ねてきたといった、ビリヤード台の解説のようなものになり、それは一回性の、履歴による、因果としての描写とならざるをえないだろう。(ここで、空間スケールを小さくしたから時間スケールも短くして、時間平均が充分取れないものと仮定している。)
そうしてみると「一回性の科学なんてものはないのだから、一回性の減少に見えるものは、エルゴード性があるスケールに(仮想的にでも)視点を持っていったうえで扱うしかない」という立場もある気がして生きた。
たとえば、生命の発生についてたまたまRNAができたことによる履歴の効果でこういう生命ができたことにこだわらずに、統計的にはこれこれこういう生物もできた、という例を多数生成させたうえで、それらをまとめて説明できるような統計的な法則こそが生命を説明する法則であるのではないか。一回性の事実にこだわっているかぎり、生命と進化の科学的な説明はありえない。
意識とか心の発生についても同じ理屈が成り立つだろう。意識や心ならすでにたくさん人数あるだろってことにもならない。なぜなら、一人の人間が、しかも他でもなくこの私が、10万人の人間の主観的経験を入れ替わり経験して(しかも充分長い時間、無相関な形で)、そのうえでそこから法則性を導き出さなければならないという帰結になる。たぶん。あくまでもその立場ならば。
もしかしたらいまアタリマエのことを言っているように見えるかもしれない。というのも構築主義の人はすでにそれをやってるように見えるから。しかしそこで可能な一つの例を生成するのと、複数生成する法則を見つけるのは別の話ではないだろうか?
自分で言ってて手に負えなくなってきたのでここまで。
先日のこの話題に関連することをEvan Thompsonが言及してた。つまり、FEPはエルゴード性を前提としているだろってこと。
Friston's free-energy principle is based on the premise that living systems are ergodic. Kauffman begins his new book with the premise that life is non-ergodic. Who is right? My money is on Kauffman on this one, but what do I know? A World Beyond Physics https://t.co/dYsGKa02PP
— Evan Thompson (@evantthompson) May 7, 2019
でもそれは上にも書いたように、コインの裏側というか、連続したものの両端であって、われわれは普段はそんな不確定性な一回性を生きているわけではなくて、必要に応じて現象学的に見るとき、もしくは瞑想を通して見るときにそのような一回性が見えてくるということではないか。そしてそう見ることによって初めて両者がどのように世界を作っているかが見えるようになるのではないかと思うようになってきた。
2018年11月29日
■ 「意識研究ネットワーク」の年会(CoRN2019)を1月に岡崎で開催します
こんど1月に岡崎で意識研究に関する国際シンポジウムを開催します。参加申し込み、ポスター発表申し込みを受付中です。ぜひご参加ください。情報は以下の通り。各分野のMLに流しているのと同じ文章です。このブログではもう少しシンポジウムの狙いなどについて書いてみましょう。
Consciousness Research Networkは、意識研究のアジア・パシフィック地域での共同研究推進のネットワークづくりをめざして活動をはじめました。WebサイトでAdvisory Boardのメンバーを見てもらうとわかりますが、だいたい国際意識学会(Association for Scientific Study of Consciousness, ASSC)に関わっている人が多いです。しかし、ASSCおよびツーソンとは異なる第3の極となることを目指して、まずはアジア・パシフィック地域でのネットワークづくりをしているという状態です。
昨年第一回の年会を台北にある国立陽明大学で行いましたが、講演者19人、参加者100名以上という盛況な会となりました。第二回の年会はこの日本で、このわたし吉田がlocal organizerを行うということになりまして、アラヤの金井良太さんと相談しながら、そして途中からは名古屋外大の佐藤亮司さんにも加わっていただいてプログラムを作成してきました。
本年会の特色は「討論セッションとそれに先立つチュートリアルセッション」の時間を設けた点です。昨年の神経科学大会後のサテライトシンポジウムでBeyond NCCで4つのお題について議論する、というものだったのですが大変盛り上がりました。盛り上がりすぎてけっきょく3つしか終わらなかったっていう。これの発展形を今回加えたいと思ったというわけです。
テーマに関してですが、私のこだわりで、科学者と哲学者がもっと会話できるようにしたいということから討論セッション#1のテーマは「意識研究のために哲学者と科学者はどのように協力すればよいのだろうか?」、討論セッション#2のテーマは「意識研究において理論的なアプローチは必要だろうか?」というものにしました。
討論セッションのテーマを考えるにあたって、はじめに持っていたイメージは「科学を語るとはどういうことか —科学者、哲学者にモノ申す」だったんだけど、そんでもって討論セッション#1のテーマは「意識研究のためには哲学なんて要らないんじゃないの?」「意識研究のためには科学なんて要らないんじゃないの?」みたいな荒れそうな感じを考えてました。でも荒れるだけだと相互理解が深まらない。そこで議論をする人には前提知識を共有しておいてもらいたい、そう考えて、チュートリアルセッション#1では「心の哲学と意識のハードプロブレム」について哲学者(東大のJohn O’Deaさん)にレクチャーしてもらって、参加者はそれを同じ日の午前に聞いておく、というふうにしたというわけです。
討論セッションではパネリストの方に5-10分程度話題提供をしてもらって、そこから議論を始めるということにしています。討論セッション#1のパネリストの一人は東海大学の田中彰吾さんが確定してます。田中彰吾さんはリハビリテーションの現場などで哲学者として関わってきた方ですので、どのように哲学側から寄与できるかお話いただけることでしょう。もうひとりのパネリストを探しているところなのですが、科学者側から語れる人がいいですね。自分がやりたいという人は吉田まで連絡ください。
もうひとつの討論セッション#2としては、Beyond NCCのときに盛り上がった(紛糾した)情報統合理論IITについてもう一度、海外の人も含めた上で議論したいと考えました。ただし、あのときの議論を考えると、情報統合理論IIT自体の理論構成の細部に立ち入って議論するよりは、より一般性の高い問題、つまり「意識の理論というものはありうるのか、あるとしたらどういう形になりうるのか」ということこそが議論するべきところではないかと思いました。パネリストとしては科学者側がアラヤの金井良太さん、哲学者側が広島大学の宮園健吾さんに話題提供と問題提起をしていただけることになってます。討論セッション#2に先立つチュートリアルセッション#2では、そのような理論的アプローチのひとつとして、大泉匡史さんに意識の統合情報理論についてレクチャーを行っていただきます。
今回のシンポジウムの裏テーマは科学者と哲学者の対話となっています。ひとつの理由はConsciousness Research Networkの前回参加者が日本はほとんどが科学者で、台湾、中国はほとんど哲学者だったということがあって、国際共同研究を促進する際には科学者と哲学者との対話がそもそも必要だろうと考えたというのがあります。
もう一つの理由としては、私がASSC15(2011年に京都で開催、代表は松沢哲郎先生)でプログラム委員をやっていたときの印象ですが、私自身が科学者ということもあって、日本国内の哲学者に十分リーチできていない、十分議論できる機会を持てていないというものがありました。今回はそこをなんとかしたい、ということでそれ以降にできた繋がり、とくに北海道大学の田口茂さん、名古屋外大の佐藤亮司さんを通して意識研究に関連性のある哲学者に声をかけていった次第です。
さあどうでしょう?討論セッションもチュートリアルも面白くなるんではないでしょうか?討論セッションが盛り上がるかどうかは参加者しだいですので、ぜひナイスなツッコミができる方の参加をお待ちしております。岡崎でお会いしましょう!
ポスターセッションもやります。ぜひ演題出してください。意識研究そのものでなくても大丈夫。将来意識研究をしたい人が自己紹介するつもりでいまやってることを持ってきてくれればよいです。こちらもぜひよろしく。
- 日時: 平成31年1月23日(水)13時 ~ 25日(金)18時
- 会場: 岡崎コンファレンスセンター(名鉄東岡崎駅より徒歩10分)
- 参加費:無料
- 研究会webサイト:https://www.conresnet.org/
- 基調講演:
- 北澤 茂 (大阪大学)
- 一般講演:
- Qiufang Fu (Chinese Academy of Science, China)
- 本城 咲季子 (筑波大学)
- Po-Jang (Brown) Hsieh (Duke-NUS Medical School, Singapore)
- Hakwan Lau (UCLA, USA, University of Hong Kong, HK)
- Ying-Tung Lin (National Yang-Ming University, Taiwan)
- 田口 茂 (北海道大学)
- Lu Teng (NYU Shanghai, China)
- 渡辺 正峰 (東京大学、マックスプランク研究所)
- 討論セッション
- #1 “How to collaborate philosophy and science for consciousness research?”
- パネリスト: 田中 彰吾 (東海大学) + TBA
- #2 “Theoretical approaches to consciousness research: Is it necessary? Is it possible?”
- パネリスト: 金井 良太 (株式会社アラヤ) + 宮園 健吾 (広島大学)
- #1 “How to collaborate philosophy and science for consciousness research?”
- チュートリアルセッション
- #1 “Introduction to philosophy of mind and the hard problem of consciousness” by John O’Dea (Univ Tokyo)
- #2 “Introduction to integrated information theory (IIT)” by 大泉 匡史 (株式会社アラヤ)
- オーガナイザー:
- 金井 良太 (株式会社アラヤ)
- Ying-Tung Lin (National Yang-Ming University, Taiwan)
- 南部 篤 (生理学研究所)
- 佐藤 亮司 (名古屋外国語大学)
- 吉田 正俊 (生理学研究所)
参加費無料。参加登録、ポスター申し込み受付中。
くわしくは研究会webサイトから:https://www.conresnet.org/
2018年08月27日
■ Asynchronous State
「EIバランスによって"Asynchronous State"ができる」という話題をJCで採り上げることにした。でも実験データ自体は「同時記録した100個のニューロンのスパイク相関は予想外に低い」これに尽きる。どうしたものか。
EIバランスによってカオスができることとか、刺激応答によって応答のばらつきが落ちることとか、こういうことちゃんと考えて、resting-stateでの神経活動のスパイキングニューロンネットワークのモデルを作れるようにちゃんと理解したい。
Okun et alのほうもただそのとき同時に記録できたもの(サイレントな集団は無視している)をpopulation couplingとして捉えるということで、なんだか判然としなかった。重要だってことはわかるし、計測のほうが追いついてないって事情も分かるのだけど。
JCの準備終わった!けっきょく、spikeはirregularである(Softky & Koch) => EIバランストネットワークでirregularなパターンが再現できる(van vreeswijk and sompolinsky 1996) => このネットワークでregular<->irregular / sychronous<->asynchronous という関係をモデル化できること(Brunel 2000) => ではin vivoでもasynchronousなのか=> Renart 2010 in vivoデータ => ではこのようなasynchronous state (というかlow spike correlation)はなにでできるか => Eの活動をIの活動がちょっと遅れてトラックするから(Renart 2010 IFモデリング) => このようなdecorrelationの意義は? =>弁別をする際にnoise correlationを減らすことで効率のよいdecodingに寄与する、というストーリーにした。
Renartに持ってくるところが強引だが、そもそもこの論文、asynchronousであることの証拠にspike correlationしか出てこないので、それをフェアに評価しつつ紹介するとしたら「モデル側ではasynchronousであることの計算論的意義とかは散々言われてきたけど、それをin vivoで100個のスパイクを同時に記録して示したのは初めて」というところを強調することになりそう。
ほんとうはもっとEIバランスとカオスとか、EIバランスと各種疾患の議論とか、そっちに伸ばしたかったのだけれども、そのまえに実験的事実と理論的裏付け自体に時間をかける必要があることに気がついた。
Henry Markramのcell論文が出てくるのとScientific Americanでの批判記事が出てくるのが同期してるってのはなにか深いわけでもあるのだろうか?
Fig.17でRenart et al 2010の再現をしている。これ自体は(dendriteなどの構造を考慮しない)LIFニューロンのネットワークで再現可能なので、オーバースペックではあるのだけど、ともあれ自発発火のレベルまでちゃんと再現してくれている。
2018年07月30日
■ 研究関連メモ2015年9月
まじかよ、phantom limbのrewiring説に見直しが必要? Brain 2015 Reassessing cortical reorganization in the primary sensorimotor cortex following arm amputation このネタ、Hurley and Noeのexternalist説vs Internalist説で使ってきたけど、アップデートが必要そうだ。
いいもんみつけた:「興奮・抑制均衡入門」「内容は, ほぼ van Vreeswijk & Sompolinsky (1998) “Chaotic balanced state in a model of cortical circuits”の解説です. 」
Yellowish blueの話はScientific American 2010ではじめて読んで、それからいつかやりたいと思ってる。これは意識の研究になると思う。
ふと思い立ってdeep learning + saliency mapで検索してみたらざくざく論文出てきた。これで大手を振ってdeep learning論文を読める。(<-え?) ともあれ、すくなくともventral visual pathwayではsaliencyの計算だけを独立でやっているというよりは、object recognitionと一緒にやっているであろうから、この考え方は正しいだろう。一方でsubcorticalのサリエンシー検出回路はobject recognitionをしていないのでまったく違った原理で動いている可能性がある。
こういう視点からうまいこと特徴付けてやることができると、cortical vs subcortcal saliency検出のシステムという議論を説得的に行えるかもしれない。よ~し、お父さん、ヒントン2006のmatlabコード動かしちゃうぞ!(<-その前にあれとあれとあれやれ!)
これはMarrの計算論的原理の違いまで持ってきて議論できるからインパクトあるんでないの?だれかやって!
しかもこれやると、ボトムアップの低レベル特徴によるサリエンシーだけでなくて、object recognitionまで含めたfaceとかそういったオブジェクトまで含めて視線データの推測に使うことができる。つまり、サリエンシーモデルの後付け拡張でなくて、サリエンシー+物体認知でやる。
けっきょく我慢できずHintonのScience 2006のコードを動かしてる。いま2hrかけてBPを24/200回したところだがファン回りまくり。もう収束してるしから休め、と言いたい。

mnistdeepauto.mの結果(左)は200epoch回してもまだerrorが下がり続けている。mnistclassify.mの結果(右)は35epochで止めたがもう充分というかんじ。あとで重みを見てやる。面白い。

昨日の飲み会(飲んでないけど)では「マーモでrubber hand illusionを!」とか言ってみたけど、マカクでGraziano et al Science 2000があったのを思い出した。
でもラバーハンドイルージョン的な自己(parietal)と離人症でいう自己(anerior insula)って多分別モンで、石田さん、鈴木さんの論文みたいにSII-aINSで繋がってもいいのかもしれないけど、「自己」にも背側経路と腹側経路とかあったりしないかな、とか昨日話をした。
あとマーモやマカクでTPJ刺激でOBEって話もした。でもどうやって検証しろと?(<-自分で自分にツッコミ)
Grazianoのパブリケーション・リストを見たら、小説や音楽作品まで入っていて、なにこの人、ダ・ヴィンチ?と心底震え上がった。
「内容は情報と熱力学を使って生体通信を理解しようという話です」こういうの勉強したい。
でもたぶん無理。高3ときの物理の試験で5/100点とって校内下位5%に入ってから私は物理は諦めている。いまさら「自由エネルギー最小原理」とかちゃんちゃらおかしい。俺は微分方程式など捨てて「お話としての説明」だけに満足しようと思う。
だから俺は「モテない男のミソジニー」みたいなものを物理に対して持っており、物理帝国主義を嫌悪し、そして密かに憧れる。それはたぶん、計算主義や表象主義を徹底させた合理主義を密かに嫌悪し、(わかりもしないくせに)直接知覚や反表象主義に半可通的に接近しようとする俺の傾向と通底している。
PsychoPyとEyeLinkを組み合わせるとPyGameの制限で刺激提示をプライマリモニタにしないといけない。で、しばしばPsychoPyがセカンダリから帰ってこなくなる。Spectacle https://spectacleapp.com/ というので対処できることを知った。
Walter Pittsの生涯については、Lettvinの証言とかの短い記事は読んだことがあるが、これだけ詳しいものは初めて見た:The Man Who Tried to Redeem the World with Logic
この部分とか:“Nature had chosen the messiness of life over the austerity of logic, a choice Pitts likely could not comprehend.” messinessキタ!
元ネタのひとつはこれか。あいにく読めない。Walter Pitts by Neil R. Smalheiser Perspectives in Biology and Medicine 43.2 (2000) 217-226
2018年06月25日
■ ブロードマンの脳地図の欠番
先日のTwitterでのやりとりで神谷さんがブロードマンの脳地図の欠番について言及していた。
あえて欠番をつかうとか。
— 'Yuki' Kamitani (@ykamit) 2018年6月22日
Brodmann16 (←ひねり過ぎで意味不明)
これはなんかのときの話のネタになるかもと思ってざっと調べてみた。
ブロードマンの脳地図では13-16野と49-51野が欠番になっている。たとえばMark Dubinのwebサイトにある脳領野のリスト。今回はとりあえず13-16野だけを話題とするけれども、なんで13-16野が無いかというと、これはあくまでヒトの脳地図だからで、他のnon-human primatesでの地図では13-16野は島insulaに割り当てられている。
このことはZilles and AmuntsのNature Reviews Neuroscience 2010には以下のように書かれている。
Each cortical area of his human map is labelled by a number between 1 and 52, but areas with the numbers 12–16 and 48–51 are not shown in his map. Brodmann explained these ‘gaps’ with the fact that some areas are not identifiable in the human cortex but are well developed in other mammalian species. This holds true particularly for the olfactory, limbic and insular cortices. The insular cortex is segregated into areas 14–16 in Old World monkeys (for example, Cercopithecus) and into areas 13–16 in prosimians (for example, Lemuridae). Brodmann could not find homologous areas in the human brain.
この総説のSupplementary information S1でブロードマン(1909)英訳版の図が転載されていて、旧世界ザル(オナガザル)や原猿(キツネザル)にはinsulaに13-16野があることが示されている。
ブロードマン(1909)英訳版はGoogle Booksで少し読むことができる。119ページのFig.89ではヒトのinsulaがJ.ant, J.postとなっていて、番号が割り当てられていない。また122ページのinsularの記述では1904年の段階では4つの領域に分けていたが、1909年版では2つに分けるとしてあり、番号は割り当てられていない。Google Booksでは123ページにはアクセスできないが「より正確な領野の同定には今後の研究を待たなければならない」と書いてある。もしかしたら将来的にはヒトのinsulaでも13-16野を割り振るつもりだったのかもしれない。
ブロードマンの脳地図が未完成であるということについては河村満氏の記事「ブロードマン没後99年に寄せて」(週刊医学界新聞)に記載がある。ブロードマンは50歳直前に亡くなっていて、それまでに脳地図もアップデートを繰り返している。河村満氏のもうひとつの記事「情動領域とBrodmannの脳地図:とくに12野について」(臨床神経学)にもう少し詳しい記載がある。さらにこのシリーズの決定版と思しき12ページ長の記事が「ブロードマンの脳地図をめぐって」(神経研究の進歩)にあるようだが、残念ながら当施設では購読してないので未読。
ブロードマンの人生についてはNeurosurgery 2011という記事に記載がある。けっこう苦労人で、ドイツの中をあちこち移動している。フランクフルトからベルリン、チュービンゲン、ハレ、ミュンヘンというかんじで。ベルリンでVogt夫妻のもとで主著であるBrodmann (1909) "Localisation in the Cerebral Cortex"を出版してから、テニュアの教授になるために教授資格論文(habilitation)を提出したけどリジェクトされている。Vogtの(Facultyへの)怒りのコメントが引用されてる。
“every effort to provide [Brodmann] with a modest, but secure living has failed, mainly due to non-understanding. The Medical Faculty in Berlin thereby carry great guilt on their shoulders.”
それでブロードマンは1910年にチュービンゲンに移動して、医師として働きながら空き時間に解剖学の実験室をセットアップをしつづけて、1913年にやっとチュービンゲン大学の医学部の教授になる。でも第一次世界大戦が始まり、ブロードマンは志願して精神病院で働いたので研究は中断している。そのあと1916年にハレで検死解剖が可能なポジションを得て、1918年にクレペリンに招かれてミュンヘンのPsychiatric Research InstituteのTopographical Anatomy部門長になって、これからというところで肺炎で急死。
まあこのあたりについては「ブロードマンの脳地図をめぐって」(神経研究の進歩)を読んでもらったほうがよさそう。ではまた。
2018年01月23日
■ 「平衡電位になっても細胞内Kイオン濃度は減らない」って話
以前のブログ記事「静止膜電位はどうやってできるの?」で書いたように、(たとえば)Kイオンが平衡電位に達するためにKチャネルを流れるイオンの量は非常に小さいので、細胞内のKイオン濃度には影響しない。デルコミンの「ニューロンの生物学」p.91に実際に計算してあるのを見つけたので引き写しておく。
直径 の細胞があるとして、細胞内のKイオン濃度は , 細胞外は となっている。このとき
細胞内のKイオンの総数 = 細胞体積( ) * Kのモル数( ) * アボガドロ数( 個
いっぽうで細胞膜の静電容量 なので、細胞内電位がKの平衡電位である に帯電させるために必要な電気量は あたりで となる。
この細胞の表面積は なので、必要な電気量は となる。
これをKイオンの個数に変換するためにはファラデー定数とアボガドロ数をつかって 膜を移動するKイオンの総数 / ファラデー定数( ) * アボガドロ数(
よって[細胞内のKイオンの総数]に対する[膜を移動するKイオンの総数]の比率は となる。つまり100万分の1程度しか細胞内Kイオンは流出しないので、無視できる量であると考えられる。
でもこの話には続きがある。イオンの流入が神経細胞の体積からしたら微々たるものってのは本当だけど、スパイン内の局所領域の は影響受けるかもしれない。また、細胞外間隙は考えられているよりもずっと狭く、 はアストロサイトでの取り込みとかと合わせて調節されているはずだ。
上記の計算をスパイン内の局所領域について考えてみることにしよう。スパインの体積は河西研のサイトより、 くらいで揺らいでいるとのことなので、ざっくり直径 の球で近似してみる。
さっきの計算との相対値だけで済まそう。球の直径は から へと になった。[細胞内のKイオンの総数]には体積で効いてくるから になる。[膜を移動するKイオンの総数]には表面積で効いてくるから になる。
ということは[細胞内のKイオンの総数]に対する[膜を移動するKイオンの総数]の比率 は 倍になって 程度となる。というわけで細胞体全体よりは影響が大きくなったけど、まだ5千分の1で済んでいると言えそう。
のほうについてはどうだろうか?
ここ最近の進展を調べてみたら、"Potassium diffusive coupling in neural networks"って総説を見つけた。ざっくり要旨から推測するに、 のオシレーションが海馬の近接する神経細胞の活動同期に関わっているという話で、なるほどさもありなん。
神経細胞のネットワークはシナプス伝達や活動電位によってできた正確なデジタルの論理演算機なんかではなくて、こういうアナログな漏れ出しに影響を受け、進化の過程でそれを活用するようなmessyな解法に依存しているんだ、みたいな話は好き。
2017年11月17日
■ 2015年9月英国IPC2015およびSussex大学訪問記
ギリギリまでいろいろ準備してて、やっとオックスフォードに行く準備が出来た。寝室に掃除機をかけ、雑巾で畳を拭き清める。サセックス大学でのトークは今から作る。神経科学大会のシンポから半側空間無視を抜いて、上丘のスパイキングネットワークモデルを広げる方向で。
オックスフォードで開催されるinternational pupil colloquium (IPC 2015) でのトークは紆余曲折あってなぜか15分(質問込み)に。こちらは30分シンポジウム用に作ったものを改変しないといけない。無茶だ。
明日は朝早く羽田から出発。東横インから無料バスが出ていることを知った。British Airwaysは24時間前からチェックインできるし、モバイルアプリもあった。ということでだいたい準備整った。あとは寝坊さえしなければ。
イギリスに到着!ヒースロー空港の入国を通過するのに35分かかった。空港からオックスフォードに向かうバスに乗ったところ。最終目的地到着まであと80分。
飛行機の中でスライドとしゃべり完成させた。いつもの say -f talk.txt -v Alex -o talk.aiff でしゃべり音声ファイルを作って8分ちょっと。実際にしゃべるとたぶん12分くらいになるだろう。だいたい準備出来た。
会場はオックスフォード大学のうちPembroke Collegeというところで、この古めかしい建物の中はきれいな施設で、宿泊施設も講演会場もぜん揃ってる。

昨日は22時までなんとか寝ないように粘ってから寝たら、途中4時に目覚めたけど最終的には7時まで眠ることができた。9時間睡眠って赤ちゃんかYO! 健康的だ!窓の外は雨が降っていて寒そう。イギリスだ。短パンポロシャツが使える機会はなさそう。
出張中に急ぎで対応しなければならない事項が勃発。シンポジウムもそこそこにメール作業へ。結果として飛行機に乗ってるあいだにスライドを終わらせておいて正解だった。今日は街を歩いて回る余裕はなさそう。
食事は朝昼晩TESCO express (スーパー)でサンドイッチと缶詰。つかポンド高すぎて外食できない。バーガーキングで7ポンド払ったら1300円相当とか笑える。
Pupil Colloquiumの後は @ksk_S さんに対応してもらってサセックス大学でセミナーをさせてもらうことになった。Webサイトに出てる。
時差ボケで眠いが、23時まで粘って発表練習してた。喋ってると眠さを抑えられる。通しでだいたい10分30秒。本番はもっと引っかかるから12分くらいか。まあ妥当な線。
知り合いがほとんどいない学会のうえ、座席についての夕食という状況で正直困ったが、隣りに座ったスペインから来たoptometristの院生がいい人だったので、それぞれの国の事情とか話しながら案外楽しく過ごした。
講演は無事終了!だいたい時間ぴったし。あとで名前の知らない初老の先生から"the most enthusiastic talk"と評されたので(いつものことではあるのだけれど)、それは必ずしもよくないですよね、と言ったらいい講演でしたよと言ってもらえた。今回は練習の成果があった。
一緒にチェアをしたArash Sahraieが講演前のコーヒーブレークで「フリービューイングの話、面白いね」と言ってくれたのだけど、準備していたのはSci RepsのYN課題の話なのだった。一瞬トークの内容を変更することも考えたが、練習全部ぶっ飛ぶのでそのまま行くことにした。
それから特筆するべきは、自分のトークの時間にLarry Weiskrantzが聞きに来てくれたことだ。彼は「盲視」という言葉を作った人でこの分野のゴッドファーザーだ。ずいぶんお歳を召していて耳も遠いようで挨拶するだけにとどまったが、ついにここまで辿り着いたかと感慨深い。
そして共同研究のきっかけも掴んできた。食事の後にさっそく何ができるかディスカッション。そんなに仕事増やしてどうすんの、という話もあるが、種を蒔けるときに蒔く。さいきん正直しんどい時期を迎えているのだが、なんだかもういっちょやったろうという気が出てきた。とはいえ今日は疲れた。寝る。
IPC2015、最終日のディナーも終了。隣りに座った大御所の先生に「君はとてもいい声をしているから歌を歌ったほうがよい。これが君にとって今回の学会における最大の教訓だ」と言われた。そんなこと言われたことないですよ、と答えても大真面目だった。明日から歌手に転向することにしますた。
Sussex大学のスライドもだいたい完成。前半awarenessで後半saliencyという構成。いま思案しているのは、[皮質下でのsaliency]と[皮質でのPE(bayesian surprise ~= saliency)]との関係の議論まで盛り込むかどうか。
皮質ではPEはpredictionとカップルしていて順逆ループを構成しているけど、皮質下でのサリエンシーはたぶん別の構造を持っている、みたいなことを議論してみたい。そういうことするのと、スパイキングネットワークの話を入れるのとどっちが面白がってもらえるかどうか。

ブライトンに到着した!例の観覧車のあたり。B&B(と言っても朝食は出ない)の小さい部屋に入って、ベッドに寝転がって、さてくつろごうと思ってドクペを開封したら泡が噴出して、ベッドとズボンが台無し。ラップトップにも一部かかったがギリギリ回避。というわけでいきなりブルー。
Sussex大学でのセミナー終了! 参加人数は25人くらいで部屋の椅子は8割がた埋まる感じだったので、盛況と言ってよかったと思う。けっきょくYes-no課題、電気生理+ムシモル、サリエンシーで話をしたら時間いっぱいに。真ん中は削ってしまうくらいでバランスとしてはよかったようだ。
トークのあとで聞いた感じでは、データと結論がはっきりしていて説得的だとか褒めてもらえたので、だいたい良かったかんじ。あと、Michael F. Landが聞きに来てくれたのは良かった。日常生活での視線計測の大家だから期待していた。
Michael F. Landからのコメントは「フリービューイングでは被験者はimplicitなゴールを作ってしまうものだがどう対処している?」というものだった。これはFAQかつ難問で、Yarbusの課題の影響の議論を踏まえている。
{kind=link}
答えとしては、「これは難問であって、複数の課題(たとえばwhere’s Waldo課題を含める)で比較しないと解決しない」「ただし現在のデータに関しては、centerに向かう傾向を除くために、peripheryに向かうサッカードだけで解析しても同じ結果が出る」ということを伝えた。
Anil Sethも来てくれてサリエンシーについて質問があったけど、こちらは「トップダウン注意はどう考慮すればいいと思う?」というこれまたFAQかつ難問。「課題のゴールを明示的にマーキングするなどの方法、そしてBayesian Surpriseで現在のモデルからの予想外を検出」というアプローチについて答えて、サリエンシーの問題がpredictive codingでのprediction errorとして捉えられることについてセミナー後に話をした。Friston 2012論文での顔を見ているときの視線データのモデル化についての話になったので、マカクは目を見ないけどマーモセットは目を見るので、そちらはデータを既に持っているのでモデル化するかも、とかそういう話をしてた。というわけでSussex大学への来訪もいろいろ収穫大きかった。
夜の帰り道(23時)に「ブライトン日本フェスティバル」に通りがかったらKissみたいなかぶりものしたバンドが演奏してた。観客に混じって一緒に踊っていたら「日本おめでとう!」とか言われて一緒に肩くんで踊った。帰ってきてから、ラグビーWCで日本が南アから大金星というニュースを知った。
UK出張の全行程が終了。ヒースロー空港に到着。3時間以上前に到着して、チェックインオープン前から並んだが、安全審査は5分で通過。拍子抜けした。
昨日の夜の寝苦しい時間から、バスに乗っている時間までのあいだに将来のいろんな可能性について極論含めていろいろ考え、なんだか頭が疲れた。それでも一休みすればもう一度なんとかしようという気持ちが湧いてくる。レジリエンス!
2017年08月24日
■ 「よくわかるフリストンの自由エネルギー原理」作成しました
スライド「よくわかるフリストンの自由エネルギー原理」を作成してslideshareにアップロードしました。
このスライドは栢森情報科学振興財団 第17回 Kフォーラム2017での発表をもとにして作成しました。そのときのスライドの構成は「1) 盲視についての吉田の研究の紹介 2) 自由エネルギー原理についての直感的な説明 3) 自由エネルギー原理を意識の理論にするためには」という3パートからなっていて、今回のスライドは2)の部分に該当します。3)についてもそのうち発表したい。
このスライドは以前作成したスライド「アクティブビジョンと フリストン自由エネルギー原理」の更新版と捉えていただけるとよいかと思います。以前のスライドについてはブログ記事(「セミナー「アクティブビジョンと フリストン自由エネルギー原理」スライドをアップロードしました」)にさらに情報があります。その当時は自由エネルギーの式をさらに簡略してあって、それだけ見てもどう自由エネルギーが計算できるかわからなかったのですが、今回は単純なケースではあるけれども、実際に自由エネルギーの値を計算しています。
FEPを正確に理解するためには機械学習(変分ベイズ)、そしてニューラルネットワーク(確率的勾配降下法)についての知識が必要となります。しかしそもそもそこまでしてFEPを理解する意義があるか、判断の材料を提供するために、神経科学者、心理学者、哲学者に向けてFEPの最小限の理屈を説明したい、というのが本スライドを作成した動機であります。そういうわけで、この説明では(世界一)単純化した例を用いているということを承知しておいていただきたい。
私自身はFEPをどのくらい重要だと思っているかというと、批判的に継承したいと考えていて、Kフォーラムでは3)の部分でその話をしました。そのうちそちらもまとめる予定。
2017年08月06日
■ 最尤法、MAP法、ベイズ推定についてのまとめ
ずいぶんと昔の話なのだけど、「比率のデータにエラーバーを付けたいんだけど」っていう記事を作ったときに、最尤法でも尤度関数の分布を使ってエラーバー(ベイズ的なcredible interval)を付けることができるよね、というのをやったことがある。そのときどうもしっくりきてなかったことを整理してみる。
最尤法の具体例として(この例自体は今回の話に必須なわけではないが)、二項分布でコインの表裏を で表現するとして、n回のコイン投げデータ が確率モデル(二項分布)のパラメータ (コインの表が出る確率)の尤度関数 は
となる。最尤法ではこの尤度関数 の最大値となるパラメータ を推定する。
これはベイズの枠組みで言えば、無情報のprior を使って計算したposteriorの分布の最大値となるパラメータ を推定することと等価だ。
いっぽうでベイズ推定の場合には、尤度そのものを使うのではなくて、ベイズの定理からposteriorの分布を推定している。
もしここで点推定したければ、
となる。
こうしてみると、posteriorの分布を推論する(inference)ところと、そのあとパラメーター を点推定(estimate)するところを分けて整理できるなと思った。
さっそく作ってみた。
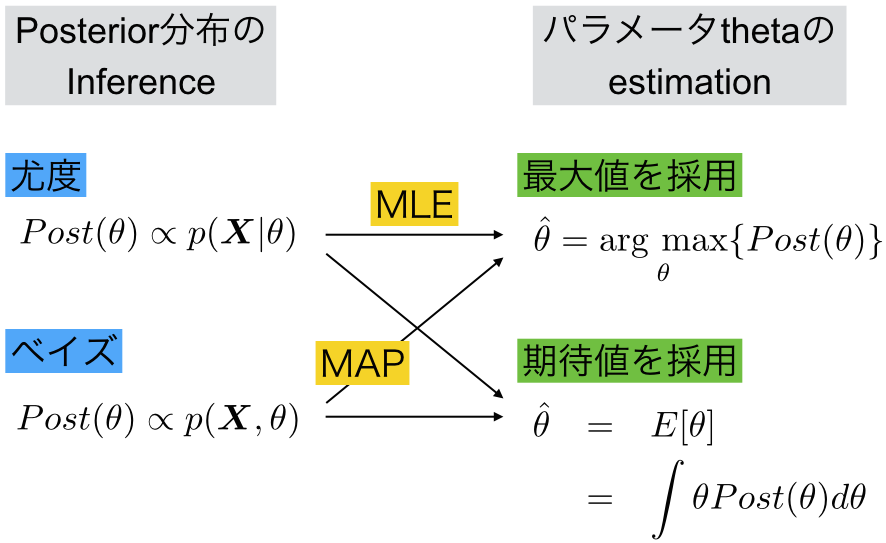
こんなかんじ。ベイズの定理に基づいてposteriorの分布を推論(infer)する方法に対して、priorの情報を使わない尤度を用いた方法がある。どちらもパラメーター の推測(estimate)を行う際には、最大値を採用する方法や期待値を採用する方法がある。
尤度関数の分布を求めた後に最大値を採用する方法が最大尤度法MLE(maximum likelihood estimation)であり、ベイズの公式でjoint probability (=generative model)の分布を求めた後に最大値を採用する方法がMAP法(maximum A posterior)だと。
つまり最大尤度法MLEという言葉を私が気持ち悪いなあと思ったのは、尤度関数を計算するところと、最大値を推定するところとが両方いっぺんに入っているからだということがわかった。「分布関数を計算する尤度推測(likelihood inference)」とそのあとの「最大値を採用する点推定MLE」みたいな言い方をするほうが混乱しなくていいんじゃないの?
なんか、MLEは点推定するけど、ベイズは点推定しないみたいな言い方はmisleadingだと思っていたので。
もちろんこれはベイズ史観であって、MLEを作ったフィッシャーの狙いとは異なるからこんな言い方はしないんだろうけど、そういった歴史的経緯以外にこの捉え方でまずいところってあるんだろうか?
でもってじつは本題は、このように整理すると、さらにhidden variable があるときのEMアルゴリズムと変分ベイズを並べることで2*2のマトリクスが作れるよって話だった。でもそこまで図を作る前に息切れしてしまった次第。
ところで、当たり前っちゃあ当たり前なんだけど、 も も も の関数であって、同じ一つのグラフの上に書ける。
それは当たり前のことなんだけど、式を字面だけ読んでいると、なんだか が の関数であるような(ぼんやりとした)勘違いをしてしることに気がついた。もちろん、べつに条件付き確率の左右にはどちらがgivenかといったそういう意味があるわけではない。
2017年07月07日
■ EMアルゴリズムの勉強メモ
自由エネルギー原理を理解するためには機械学習での「変分ベイズ」を理解する必要があって、さらにその手前の段階に「EMアルゴリズム」がある。EMアルゴリズムにおいてもKL divergenceを最小化して下界Lを最大化する過程が出てくる(PRMLの9.4章の図9.11-14)。
この図と式を字面を追っていくことはできるけど、シンプルなモデルでじっさいにグラフを書いて理解できるようにしたい。いちばん簡単な例はなんだろうか? Nature BiotechnologyのPrimerの記事で混合二項分布を使ったいい感じにわかりやすいものを見つけた:Do, C. B., & Batzoglou, S. (2008). What is the expectation maximization algorithm? Nature Biotechnology, 26(8), 897–899.
ここにある事例を使ってmatlabでグラフ書きながら理解してみることにしよう。
潜在変数がない場合 (=>最尤推定)
コインの裏表のデータがある。データXはコインの表の数(列1)と裏の数(列2)を表していて、10回での裏表の数を計算して、それを5回繰り返しす。つまり、データ で、たとえば だと、10回中5回表が出たということ。
じつはコインには二種類、コインAとコインBがあって、それぞれで表が出る確率が違う。この5回でどっちを使ったかのデータがわかっていて、それをZで示している。つまり、データ で、 (A or BでAを使った確率)
それではcoin Aおよびcoin Bそれぞれでの表の出る確率 はいくつか?
これは最尤推定で解ける。しかも二項分布の場合は最尤推定を使わなくても、coin Aでは thetaA = 24/30=0.80 , coin Bでは thetaB = 9/20=0.45 という計算だけで済む。
潜在変数がある場合 (=>EMアルゴリズム)
ではもし、いつcoin A, coin Bが出たか(Z)が不明の場合にもcoin Aとcoin Bの表が出る確率を推定することはできるだろうか? つまり入手できるデータはXだけで、Zのほうは入手できない潜在変数という場合でも可能か?というのが問い。
そうすると問題はこのときの尤度
を最大化するような を求めたい、ということになる。
そこで尤度 を最大化する代わりに、
における を最大化する。
これのlogを取ったものの最大化を考える。
(1) 推定する変数 および の初期化
, : コインA, Bそれぞれの表が出る確率
および
(2) E-step: を計算する
ベイズの公式より
これらが計算できればE-stepでの目的である として
右辺は(2)と(1)から計算できる。
実際にはこれを計算する:
は のときの が起こる尤度だから
初期設定より、
よって は以下のように計算できる。
同様にして、 は のときの が起こる尤度だから
初期設定より、
よって は以下のように計算できる。
まとめると となる。
同様にして について計算すると
を計算するには上で計算した と があればよい。
より
これが の推定値
(3) M-step: の推定
coinAが出る期待値は
(4) 収束条件を見てストップするか判断
thetaが収束したかどうかを確認して、E-Mの繰り返しを止めるか決める。
ではE-Mを20回繰り返して、thetaが収束するかどうか見てみることにしよう。
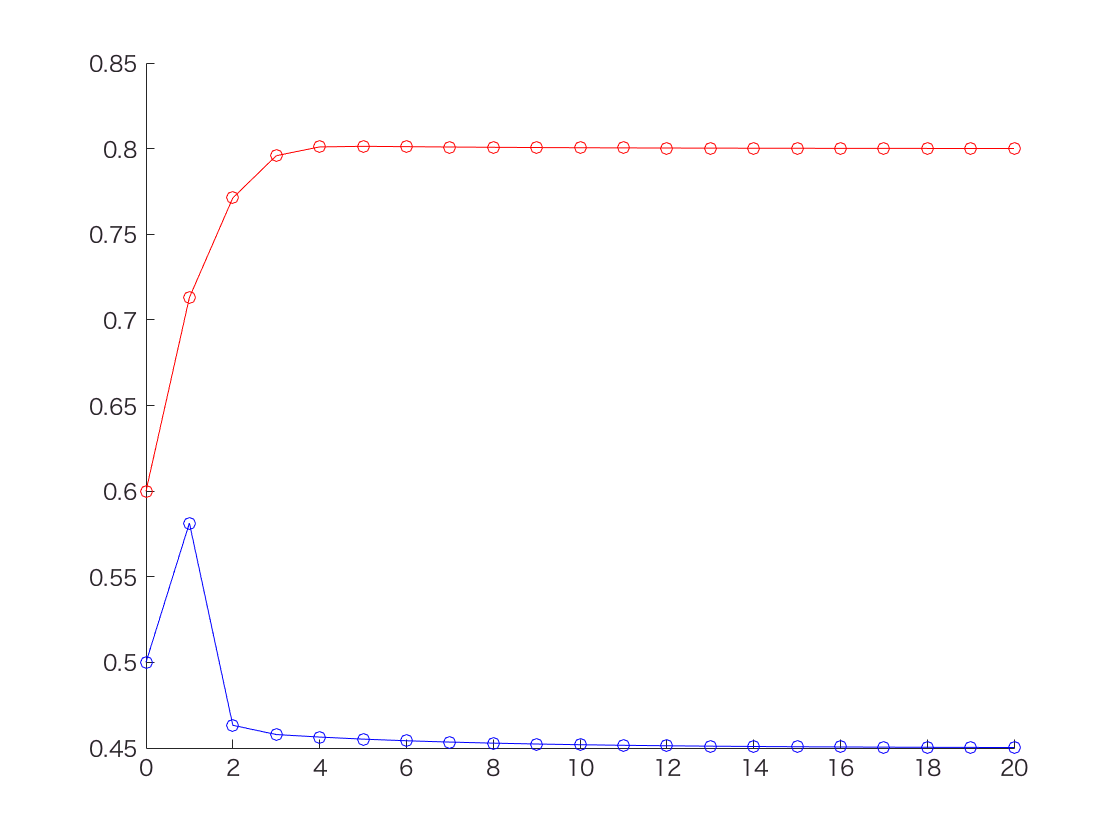
このように、Zが既知での最尤法のときに計算した値(thetaA = 0.80, thetaB = 0.45)と同じ値に収束していることが分かる。
ところでNature BiotechnologyのPrimerの記事では10回繰り返したところでthetaの推定値が thetaA = 0.80, thetaB = 0.52 となってる。たぶんPrimerの記事のほうがなんか間違えてると思う。
2017年06月02日
■ 研究関連メモ(Box2D, Monaca 2015年7月)
ProcessingでBox2Dを勉強するためにexampleフォルダのAttractionApplyForce.pdeを動かしてみた。
擬人的に表現するなら、ほかの誰もが引力に負けて太陽に吸着されるのに、たった一人が生き残り、惑星となって回り続けた。
もうひとり惜しいやつが居たが、途中で引力に負けた。最後の一人がどこで引力に負けるかと10分待ってみたが、独立性を保ち続けた。(<-「お話としての説明」と「科学としての説明」の問題?)
「Monacaでつくるモバイル・アンサンブル」行ってきたけど面白かった。クラウド上のIDEからp5.js使ってprocessingのコードを書くと、iphone上で立ち上げているMonacaデバッガでそのコードが動く。
だから、複数のiPhone, iPod Touchがあると、web上のIDEからの実行命令ひとつでモバイル機器がすべて一挙に動く。さらにサーバーを介してモバイル機器間で通信できる。赤松氏のSnowflakesを参加者で実演した。 赤松氏によるデモ(Youtube)
Supercollider自身というよりもp5, arduino, ofxと組み合わせてインタラクティブにやることに興味があるので 今日は満足した。講師は赤松正行氏だったので「『2061:Maxオデッセイ』のMax7対応版は出ないのですか?」と聞くつもりでいたがすっかり忘れてた。
植村さん@uemurag 経由で知ったティム・クレインの”New Directions in the Study of Mind” 鍵は「非物理主義的」で「非還元主義的」なアプローチ。
プロジェクトの狙いを説明するブックレットが用意されていたので読んでみた。「非物理主義的」で「非還元主義的」というとたいがい「物理主義的」「還元主義的」アプローチでは説明できない反例を出す哲学的議論になるけど、そういうのだけでなく、このプロジェクトでは「ある心的現象に対する非物理主義的説明というものがあったとしたらそれはどういうものか」を「イデオロギー的」にではなくて「探索的」に行う、とある。べつにオカルト的に行くわけではなくて「物理主義への懐疑的態度と心への科学的なアプローチの組み合わせ」とも書いてる。
ここでいう非還元主義的というときは「脳全体のネットワークの創発的状態で意識ができます」みたいなのは当てはまらない。NCCを単一細胞から脳全体の状態に置き換えただけだから。コッホがIITに肩入れしているのはあれがより洗練した形での還元主義だからなわけで。
脳の状態だけに依らず、体とか環境とかも含めたシステムとして心が出来るんだ、ってのも非還元主義的ではない。神経科学的立場から「非物理主義的」で「非還元主義的」可能性を考えるなら、脳と体と環境が全く同一(力学系的な軌道まで含めて)であるにも関わらず心的状態が異なる場合だろう。
プロポーザルの例として書いてあったのは「どうすればNCCの探索というプロジェクトを意識が自然の中で占める位置を決めるのに役に立てることが出来るか」とかだった。
(追記:プロジェクトの期間はJuly 2015 to July 2017とのこと。ブログ化したのが遅いので時期外れになってしまった。)
2017年05月25日
■ 研究関連メモ(aphantasia, CCSM 2015年7月)
FB経由で知ったaphantasia (心的イメージを作ることができない)という話。講義で使うNed Blockの話で「哲学的ゾンビの話を講義ですると2/3は理解するが1/3は(その可能性すら)理解できない」という話に近いかも。
さらにその着想のもととなった症例報告では冠動脈形成術を受けてから心的イメージを失っている(がimagery課題はできる)。よってBlindsightの向こうを張って‘blind imagination’と表現している。
つかこれみんなホントのところどうなのか聞いてみたい。私は心的イメージは無いわけではないけど、彼らのVVIQによれば”Perfecting clear and lively as real seeing”ではなくて”Dim and vague; flat”の方だと思う。
今みたものを目をつぶって思い浮かべたときに直観像的に画像が目に浮かぶというよりは、黒い背景に筆でそのシーンを書き込むように、なんかmotorを起源としたものして持っているように思うのだけれど。
このこととsensorimotor contingency的な考えへの親和性には相関があるのかも。われわれそれぞれがなんらか心と脳の理論を持っていてその妥当性の判断基準は実は心像やら夢やらそういったものの処理能力によって制約を受けているのかも。つまりembodiedってことだけど。
洗顔か何かで顔をこすりすぎて、鼻の穴の入り口にかさぶたが出来てしまった。そしたら視野のなかでかさぶたが目立つ。どういうことかというと、普段は視野から見える鼻は片眼からしか見えないから両眼視野闘争で負けて見えないのだけど、かさぶたでサリエンシーが上がって勝ち残ったのだな。
Robert Kentridgeが盲視での色の論文を出していたことに気がついた。Curr Biol 2012でreferしていなかった。これは恥ずかしい。
Kentridgeが書いてくれたDispatchでは、ヒト盲視において色(PNAS2007)や動き(PNAS2011)の処理の原理が異なっていること、盲視におけるcueingの必要性など、この論文で書ききれなかったことを補ってくれている。
他の人がどのように自分の仕事を引用しているか調べると、自分では使わないようなフレーズが出てきて参考になる。たとえば、いつもanimal model of blindsightと書いてきたけど、experimental blindsightって言い方はよさそう。
VMware FusionでUbuntuを使っているんだけど、USキーボードとKarabinerで運用しているので、Macのほうでコマンドキーを押してUbuntuに移ってくるといちいちCaps Lockがかかるのがウザい。「Caps Lock キーの機能が逆になることがある」
解決法としては、Ubuntu上でもう一回コマンドキーを押せばいいのだが、そうするといちいちDashが立ち上がるのでESCで閉じる。これまたウザい。この問題をずっと放置していたのだけれど、CCSMというやつで解決できることを知った。
CCSMをsoftware centerからインストールして、Desktop > Unity plugin > key to show dashをsuperからsuper + shiftに変更。これでcommandキーを押してもdashがポップアウトしなくなった。快適だ!
2017年05月08日
■ 細胞外電極はなにを見ているか2017年5月版
もう10年近く前のことになるけど、以前ブログのエントリで「細胞外電極はなにを見ているか」それから「細胞外電極はなにを見ているか リニューアル版」というのを書いた。このときのコメント欄を見てもらうと分かるのだけど、私の理解が正しくなくて「volume current source density」と表示しないといけないところを「電流」と書いているために混乱しているところがあった。
それからあとOKさんから指摘をいただいたけど、PDFファイルのp.9の注釈10 「ある小領域に流入出する電流 $I$ が電場 $E$ を引き起こす。これはどんな環境でも成り立つ。$\sigma\nabla E = -I$ 」ここは完全に間違っている。
これらを直したいのだけど、部分的に直すのではなんともなりそうにないので、別の文書を作成して、基本からひととおり説明を作ることにした。目標としては、二つの式、(1) 電流源密度CSD解析の式
\[ \sigma\nabla^2\Phi = -I_m \]そして(2) ある一つの小領域での電流源 $I_m$ が $r$ 離れた部分に置いた細胞外電極に作る電位の式
\[ \Phi (r) = -\frac{I_m}{4\pi r} \]をマクロなスケールでのマクスウェルの法則と準静的条件から導く、ということをしたい。
実のところ長年なんとかしなくてはと思ってはいたのだけど、さいきん『ニューロンの生物物理』第2版を読んで第9章に非常に詳しい説明があるのを知った。それでそこを起点にしていろいろ勉強したので、そのノートをアップするという次第。
以下のslideshareへのリンクから。
2017年04月30日
■ 研究関連メモ(スパイキング・ニューロンを用いたモデリング関連)
ここさいきんspiking network modelについての仕事を進めているので、この機会に勉強しようと思ってIzhikevichのDynamical Systems in Neuroscienceを精読し始めた。
そしたら、第1章の段階で劇的に面白くてわかりやすい。これのFig.1.15の説明で、4種類の分岐によってニューロンの発火様式が説明できるというのを見て、だいたい満足してしまった。ある意味これ以降はこの図に書いてあることのより正確な説明なわけで、本としても見通しが良くて素晴らしい。
数式よりも図で理解させるというのが徹底してるし、図が過不足なく説明したいことだけが書いてあって、図にあるけど理解できないこと、というのが無いのでフラストレーションがたまらない。 1章(PDF)は著者のサイトから読める。
その昔、薬学部に入ったときに「生命を捉えなおす」を読んで感激しつつも、清水博先生が退官直前だし、プログラミングもできなかったし、高校の物理で挫折した自分には向いてないと思ってこういう人生を選択したけど、いま自分が高校生だったらこっちへ進んだかも。Pythonでできるわけだし。
Dynamical Systems in Neuroscience (Izhikevich)は4章まで読み終わった。図が多いのでページ数のわりによく進む。とりあえずNaとKだけのモデルで二次元の相空間で入力電流の大きさによってサドルノード分岐とかするところまで来た。激楽しい。
いまうちでRichardが使っているのはAdExモデルなので、それのもととなるIzhikevichモデルを理解して、とりあえずの目標としてはNEST使えるようになるところまで行きたい。
NESTだけでなく、Brianとか学習向けとしてはよさそうだし、何から触ればよいのかはまだ思案中。とりあえずIzhikevich本のmatlabコードは以前からいじってる。Processingでのコードというのも見つけた。
AdExニューロンモデルと"Neuronal Dynamics"の著者であるWulfram GerstnerのMOOCを見つけた。何個か見たけどよい。イズケビッチ本の次はこれか。
Brianのデモをいじくってた。Anaconda上のpipでインストールして、iPython上でデモプログラムを動かす。HHニューロン4000個で上が興奮性3つ、下が抑制性3つ。楽しい。
ひさびさに良いニュースがあった。もうすこしBrianとかいじったりとかする方向を伸ばす余地が出てきた。なにをしようとしているのかというと、IITとか予想コードとかそういったものを神経生理学の知見を踏まえて刷新するためには、昨今のasynchronous stateとかEIバランスとかそういったものを踏まえて、spikeとsynaptic conductanceのレベルから力学系的に扱うような粒度でネットワークの状態を評価するべきで、そのために必要なことでここ20年放置していたことを勉強し直そうというわけだった。言うことだけは威勢がよいが。
だからこれも意識研究への道であり、決してサイドプロジェクトではないのだけれど、なんか回り道している感はある。でも、薬学以来のパッチクランプとかあのレベルの知識がここでこそ活きるのではないかとか思ってる。
2017年04月19日
■ 研究メモ: 球面集中現象、一級マイクロサッカード鑑定士など(20160831まで)
「次元の呪いと球面集中現象」の話が好き。僕らは身長とか足の速さとかテストの成績とかの多次元から成り立っていて「全てにおいて平均的」(超球の中心からの距離=0)ということはありえなくて、みんなが同じくらいどっかずれてる。これって救いだろ?
この図は1-100次元の独立な正規分布の原点からの距離を計算して、期待値(=sqrt(次元数) )で割ってヒストグラムにしている。だんだん0に近いところに分布しているデータが少なくなることがわかる。
ただしこの分布はその多次元が独立であることと正規分布であることを前提としている。でも本当はそれぞれの次元は相関しているし、収入の分布みたいに正規分布してない。
田口茂さんの「現象学という思考」や鈴木貴之さんの「ぼくらが原子の集まりなら(略)」を読んでから、「表象主義批判」ではなくて「表象の起源を問うこと」こそが重要ではないかと考えるようになってきた。「哲学入門」(戸田山和久著)でルース・ミリカンを知ってから、このへんを読もうと思った。
「意味と目的の世界」ルース・ミリカンから始めようと思うが、植村恒一郎氏のブログでまとまった記述があるのを見つけた。これ読むと、人間とそれ以外の動物との違いとして後者には「オシツオサレツ表象」しか無いという議論をしていることが分かった。
このブログ記事では「アイちゃんに水を差すつもりはもちろんないが…」という表現があるけれども、松沢さんは「想像するちから」でチンパンジーは今を生きており、未来に絶望しないと書いている。これはミリカンの言ってることに近そう。
「動物の時間表象は、複数の時間系列にとどまり、前方にどこまでも伸びてゆく線形時間ではない。この線形時間こそが、「未来を変えること」「新しいものの創造」を可能にするのだが、動物においては、これがほとんど欠けている。」引用元
というわけで気分が盛り上がってきたので「意味と目的の世界」を読んでみようと思う。ミリカン本人のサイトに出版前原稿がある。第1章では「パーソナルな目的」と「サブパーソナルな目的」のあいだに明確な区切りなんて無いよって話をしてる。いきなり面白い。
「シリーズ心の哲学〈3〉翻訳篇」にミリカンのbiosemanticsが訳出されていることを知ったので早速借りてきた。原文もresearchgateからゲットできた。
ミリカンの「固有機能」って言葉がキモいなあと思って原文調べてみたら"proper function"だった。要はproper noun(固有名詞)との対応付けで、言語との絡みであとで役に立つのだろう。戸田山本では「本来の機能」と訳してあった。同じもののことだとは気づかなかった。
"Mental representation, communication and the transition from animal to human" このスライドがわかりやすくてよかった。
Gärdenforsの論文も見つけた。Mental representationといっても二種類あって、cuedとdetachedがある。cuedは「イマココ」と直結していて、detachedはそこから徐々に離れてゆく。人間とそれ以外の動物の違いはこちらのほうが扱いやすそう。
「心の中の悪口について」のブクマコメント。こういう、なかなか言いにくい内面のことについてのデータが350人くらい集まっている。これって宝の山なんかないかと思うのだけど、どう料理すればよいのかわからない。
二つの正規分布するarrayを作って、array間の相関係数をいろいろ変えたものを作りたかったのだけど、MATLAB Centralでやり方を見つけた。 これでこんな図が作れた。
これはpairwiseだけど、複数のarrayでもいけるのか?と10次元バージョンを作るためにR = ones(10,10)*0.3; R(eye(10)==1) = 1;として、L = chol(R)を作ってみたら、たしかにいけた。原理はわからないが、すげえ(<-勉強しろ)。
「数年前に起きた東名高速の40キロ渋滞。この原因は、たった一台の追い越し車線への割り込みが原因だったことが分かっています」という記事を読んだ。
でも複雑系でこのように原因を特定する意義ってあるのかな? 砂山に砂を振りかけてゆけばあるタイミングで雪崩を起こすけれども、その雪崩を引き起こした砂に原因というか責任を帰属させるのは「お話としての説明」でしかなくて、制御において意味のあることとは思わないのだけど。
じっさい、ここで提案されている解決案もその「たった一台の追い越し車線への割り込み」を排除するような個別例への対策ではなくて、「アリは混んできたら詰めないって戦略を実践」といった統計的な対策であるわけで。
小学校の国語の時間にみんなの前で音読できない子と無理強いする先生の話。 いまにして思えばこれは場面緘黙症なのだが、当時は僕もわからなかった。今はちゃんと認知されているのだろうか?
この記事のはてブを読むとほとんどが「音読の意義」というタイトルへのレスポンスばかりで、本題の「どうして先生はそういうときに無理強いするかな」 って話になってない。正直これは酷いと思うけど、同時にどうやれば正しく伝わるのかなとも思う。
今日いま此処の俺は一級マイクロサッカード鑑定士を名乗ってもいいと思うくらいに波形を見続けた。
誰もいない部屋で、遊佐未森聞きながら、夏草の線路を歩く野球帽の少年のような心持ちで解析中。
Regular saccadeの直前にあるmicrosaccadeを見つけると、とてもレアなので、「レアマイクロサッカード、ゲットだぜ!」と言いたくなる。(<-言いたくならない)
そういう眼で見ると、これはサッカードの途中で軌道を変えたのではなくて、マイクロサッカードとレギュラーサッカードが同時に起こった例があるように思える。それを証明するのは難しいけれども、もし両者が並行して起こりうるなら、生成メカニズムの議論に寄与できるかも。
窓に並べたフラスコ瓶に月の雫集めながら解析していたら、一時間で1090試行しか解析できなかった。メロウな曲調では効率が落ちることがわかったので、バキバキにアガる曲のほうがよいか。EDMかけて解析する。
さあやっとこれから解析開始。まだデータは2/3残っている。うわあ なんだか凄いことになっちゃったぞ (<-他人事っぽいカンジで)
夏休みの宿題を8/31の夜中にやるような心持ちでマイクロサッカードの解析。あの日あの時あの場所で早めに手を付けていたなら、いまごろ僕らは(ry
ホッテントリに「『共感性羞恥』というあの現象 分かる分からないで盛り上がる人々」と 「経営者には“サイコパス”が多い」が並ぶのが、なんだか感慨深い。
2017年02月21日
■ 研究メモ: 逆転スペクトル、PLIなど(20160731まで)
以前逆転スペクトルが本当にありうるかという話で、L coneのあるべきところにM coneが、そしてM coneのあるべきところにL coneがある、という場合のことを書いたけど、これは実際にありうるのだろうか?
そもそも「L coneのあるべきところ」というのがどう定義されるかというと、L-MとL+Mを作るようにRGCと繋がっているということなのであって、生きている大人でいきなりこの交換が起こるようなSF的な話でなくて、生まれながらにしてそのような交換があったとしても、その差は生まれ得ないのではないだろうか?
という話は置いておいても、2色性色覚というのはL->M->(M->M ...)とある染色体に不等交叉が起こった場合という話。
では、二重に組み換えが起こって、M->L みたいな配列が起こる可能性はないだろうか? それならば 一回目の組み換えで L->L->M とM->M ができて、もう一回組み替えればM->Lができる? それともこんなかんじのdouble crossoverしか起こりえない?
{kind=link}
(後日談:研究所でゲノムの専門家の人に聞いてみたら、M->L みたいな配列はできないだろうとのこと。)
サブクリニカルなものを反映した意識経験というものに興味があるのだけど、発言小町のこのスレッドでは異臭症の訴えが並んでいる。これらは蓄膿症でもないし、体臭でもないだろう。こういうのを解明したい。
「脳が壊れた」鈴木大介を買った。横浜へ行く新幹線で読む予定。これとか「壊れた脳 生存する知」とかから抜き書きして半側空間無視での空間への意識経験についてまとめようという狙い。
予想通り、「脳が壊れた」で半側空間無視についての絶妙な表現を見つけた。
- 「どちらかというと『左方向を見てはならない』という強い心理的忌避感、障壁がある状態」
- 「『視界の左側に猫の轢死体が転がっている』…『左前方に…義母が全裸で座っている』感覚」
- 「絶対見てはならないものが左前方にある!だから僕は右を見る。左半分の世界はないことにしたいんです僕は」
この「ないことにしたい」という言い方ははじめて見た。病態失認は伴っていないからだろうか。
2016年神経科学大会初日のハイライトはKarl Zillesのpolarized light imaging。無染色切片を偏光レンズで撮影することでミエリンの向きを推定する。技術自体は100年以上前からあるものだが(Wiener, 1912)、それをヒト死後脳で高解像度でスキャンしてさらにその画像を元にDTIでいうfiber trackingの方法を使ってfiberの向きを推定する。
たとえばFrontiers 2016のFig.5とか見ると視交叉での線維の交差がきっちり推定できる。
karl Zillesの仕事としては2010年くらいからあったようだが、今回はじめて見て衝撃受けた。DTIとかで苦労して推定しているところを段違いの解像度で推定していて、ヒト海馬でのperforant pathを見つけたとか、どのネタもスゴい。
マカクでpostmortemでDTIとか計画していたけど、なんとかこういうテクと組み合わせられないもんかと思った。でも原理はシンプルだが、獲得する画像が膨大で、定量的なことができるようなものを構築するのは相当大変だってことは想像がつく。
今日の午前中はこれに参加:Symposium S3-F-1 Frontiers of consciousness studies in mice
シンポジウム終了。クリストフに質問しそびれた。質問を頭のなかで組みたてていたのだけどまとまらなかったので。端的に聞きたかったことは「そもそもmiceに意識はあるか?(なければ意識研究の題材にすることはできないでしょ)」というものだった。
でも頭のなかで質問を作っていたときは「Adam Kepecsのundertainty論文を紹介していたが、『メタ認知』という言葉は使っていなかった。マウスにメタ認知はあるか?Kepecs論文からそれは言えるか?もし無いのなら意識研究の題材になるのか?」みたいなことを言おうとしてた。
でもあとで考えるに、そんなややこしい言い方をせずとも、「マウスで意識があると推定できるような行動的および生理的な証拠はどのくらいあるか?」と聞けばよかったのだった。(<-すごく面倒くさいタイプ)
村山さんへの質問もかなりもにょっていたが、もっとシンプルに聞くならば「体性感覚へのトップダウン入力の効果を見るならS2からの入力を見るのがstraightforwardだと思うのになぜM2なのですか?」と聞けばすっきりしてた。いつも質問しながら頭のなかをまとめているのでこうなる。
なんかの学会でポスター発表をしたときに、面識のない人がやってきて言いがかりっぽいコメントをして去っていこうとしたので、「すみませんが名前を教えていただけますか」と問うたが、結局そいつは名乗らず逃げていった。今にして思えば、ネットでのクソリプが現実にも実在するっていう例だな。
2016年12月31日
■ 研究メモ: 中井久夫の統合失調症論、スパイキングニューロンネットワークのシミュレーターなど(20160710まで)
以前も書いたことがあるけれども、意識研究の側面からは統合失調症の前駆期の主観的経験に興味がある。それは「気づきの亢進」と言われるものだけど、なかなか系統だった記述を見つけることが出来なかった。
中井久夫の本を読むべきだということはわかっていたけど「統合失調症をたどる」という本に書いてあることを読んで、まさにこれだと思った。この本も当事者研究的というか二人称的アプローチをとっていると言える。たとえば
- p.124「超覚醒感と圧倒的な抑留された睡眠切迫感とでもいうべきものの共存」
- p.125「思考はどんどん伸びていって分岐に分岐を重ねる。考えが花火のように枝分かれする。いままでわからなかったことが次々にわかる感じがするが、口に出しては言えない。」
- p.128「あらゆる可能性が一望の下に収められるような感じがあるように思われるが、もはやそれを口に出していうことは出来ない。言語活動は停止しているわけでなく、内的原語はむしろ超限的に増大している」
伊庭幸人さんのツイート(1)および(2)にもあるように、伊庭さんの「モデル選択とその周辺」の付録 : 「統計学的な病 一中井久夫の分裂病論をめぐって一」では中井久夫の分裂病論がモデル選択の観点から解釈できることを書いている。
「徴候優位性とは、ささいな徴候もそれを雑音とみずに反応することであり、モデル選択の言葉でいうならoverfittingの状況に対応すると考えられる。」 「乱数発生能力の減少はバイアスの評価を不能にさせ 、overfittingをもたらすかもしれない」
ここの議論はクリス・フリスによる精神症状のベイズ的説明を先取りしていてたいへん面白い。
「この後にくる混乱の描写は、計算機科学的にいえば「とめどない彷徨」による計算資源の枯渇(「一般フレーム 問題」における計算量爆発)を思わせる。」
ここで混乱状態を計算量爆発になぞらえているところは、統合失調症でワーキングメモリの能力の低下が起こっていることと対応付けると面白いかも。というのも、異常サリエンス仮説やベイズ的説明は「気付きの亢進」と「jumping-into-conclusion」とによって陽性症状を説明できるけど、WMの低下は陰性症状の反映としてべつに扱われている傾向があるから(陽性が先で陰性があと)。でもこの計算量爆発を考えると、両者は統一的に説明できるかもしれない。
意識学の確立のために俺ができることはなにかと考えたら、神経現象学を(盲視にしろ半側空間無視にしろ)とにかく実践してみることだという考えに至った。
神経現象学に必要なphenodymanicsとneurodynamicsを捉えるということで、「爆発の研究」とか「悪循環のコピペ」とかを想定していたのだけど、でもそれって「現れる存在」で言うところの"catch and toss" analysisであって、入力-出力様式を脱したとはいえない。
実際に「創発」を考えなければいけないのは、このループにもっとショートサーキットがあって、同時にたくさんループが回っているような状態でどれが原因でどれが結果だかわからない状況までいったものを状態の変化として捉えるような状況のことなのだろう。
するとフリストンのpredictive coding + active inferenceも「キャッチしてトス」図式だなあって見えてきた。それはそれでよいのだし、最小限のsensorimotorループを作ってそれを正確に測る、ということからスタートすべきで、そこでフリービューイングを使うとちょうどいいんではないかとか考えていた。
「神経系大規模シミュレーションのためのソフトウェア ~NEURON とNEST~」スパイキングネットワークのシミュレーターの記事は日本語ではとても少ないのでこれは貴重。まとめ部分では、NEURONとNEST(および他のシミュレーター)の共通言語を目指したPyNNが開発されていること、NESTとGENESISを繋ぐ試みとしてのMUSICの話が紹介されていた。
これも役に立った:ニューラルシミュレータNEST
Brian Simulatorこっちの日本語での解説はたぶん皆無なのでだれか作ってほしい。
あと、どのシミュレーターは何が得意かとかの比較も知りたい。このサイトみるとたくさんあるけど、 NEURON, GENESIS, NEST, BRIANくらいで考えればたぶんよさそう。
前者二つは樹状突起まで考えるやつで、大規模なネットワーク考えるのにはNESTが向いてて、BRIANはどちらかというと教育用?とりあえず自分の目的(上丘のスパイキングネットワークモデルの構築)からはBRIANを動かして入門するあたりが妥当かと考えているのだけど。
いま共同研究者とやっている仕事では彼が自前でシミュレーター作っているのだけど、fittingさえ終わってしまえば、汎用のシミュレーターも使えるはず。ただし、周辺抑制とかの結合様式が重要なので、そこを指定できる必要がある。
奈良および神戸にいたJan Morénの上丘モデルの仕事ではNESTを使ってた。それ以降の銅谷研のモデルでもNESTを使っている様子。
2016年11月08日
■ 研究関連メモ(20150615まで)
なんか、外界の視覚刺激の情報量みたいな言い方をするときに、それがまるで感受する有機体とは独立したもののように言われうとおかしいよなと思う。シャノン的な意味での情報量だったら送り先と受け手のあいだでプロトコルというか可能な信号の組み合わせが決まっているからこそ情報量が定義できる。
だから、視覚刺激の情報量と言われたら、とりあえず網膜上での視野像に変換してそれを錐体がどのくらいの空間・空間解像度とどのくらいのダイナミックレンジで応答するかみたいな言い方になるはず。
こういうことはたぶん「ユーザー・イリュージョン」のどっかで議論されているはずと思って図書館で借りてきた。とりあえず和訳本p.64あたりの要約で、aという文字が情報量を持っているのはほかの25文字ではなくaが伝えられてきたことを私たちが知っているからで、もし伝えられるのがアルファベットだけであるということを知らなかったら、aという文字にどれだけの情報量があるかはわからない。
「シャノンの情報は、送り手と受け手がどんな共通の前提を取り決めているかを知るまで、定義できない」という表現を見つけてきた。なるほどいま「プロトコル」という言葉で言おうとしていたことと同じだ。
統合失調症の動物モデルというときにはそれはtrait (=geneticなものまで含めた持続的な「特性」)なのかstate (=精神症状が出ているまさにそのときのようなtransientな「状態」)なのかという問題がある。
これは精神疾患一般に昔からある概念だけど、中間表現型という点からの論文を見つけた。Curr Psychiatry Rev. 2006 "Trait vs. State Markers for Schizophrenia: Identification and Characterization through Visual Processes" 動物モデルとの言及もどっかで見たはず。つまり遺伝子変異動物によるモデル、神経発達的モデル、薬理的モデルをどう組み合わせるかとかそういう文脈で。
C++かC#かとかってのは何に使うかによって決まると思ってる。最近の私にとってはハードウェアで決まってる。たとえばTobiiのアイトラッカーではC++、.NET、C#、Pythonと選択肢があったけどmatlabがなかったのでしかたなくC#で書いた。いまはMatlabが使えるけど。UnityもC#かjavascript。
EyeLinkはC++かmatlabかpython。可能な限りmatlabでやるようにして、それで無理なときだけ必要最小限の労力でC++とかC#とかPythonとかいじってる。
なるたけ手を広げずに済ませたいけど、Processingでプログラム作ったらけっきょくJavaもしらなくてはいけなくなりそうで困ってる。PsychoPyはスタンダロンだけで済ませようと思っているけど、ハードウェアとの接続を考えるとけっきょくPython環境を作ってライブラリをインストールしないといけない。そこまでやるかどうか躊躇しているところ。
5月になるとトイレに貼ってあるくもん雑学カレンダーの「いったんてんとう虫を食べた鳥が二度と食べようとしないように派手な色をしているんだ。てんとう虫はとってもまずい味がするんだ」って部分に「食ったんか!」と突っ込むのが日課だったのけど、今年は違う。
だって、そもそもてんとう虫って捕食動物にとって目立つ色なのだろうか? 赤と黒だし。それで思いついたけど、4色性の視覚を持つ鳥にとってはサリエンシーが高いのかもしれない。ハチの黄色と黒の警戒色も天敵にとってサリエンシーが高いのだろう。つまり相手の視覚に合わせたサリエンシーをデザインしている?
先日の駒場講義でも強調したことだけど、視覚意識の「contentの切り替え」の研究から広げてゆくにあたって「半側空間無視-空間・身体」と「統合失調症-自己・現実感」とを見てゆくことが重要なのだ、みたいなストーリーからいけば、ここでMetzingerが取り上げているBlankeとかラバーハンドイルージョンとかそういった現象からどうやって意識と自己の理論みたいなことをつくるかということを考えたい。Bayesian Brainとかsensorimotor contingencyとかそういうお話も意識のunityを考える方でこそ意味があるはずだ。
(情報処理としての意識、って話になるとかならずや何が意識に上って何が上らなくて、そして何が情報処理されて何がされない、といったところに終始してしまうので。つか盲視でさんざんこれまでやってきたし、これからもやるのだが。)
Processingでセカンダリモニタにフルスクリーンで刺激を提示する話。以前は難しかったようだが、いまはpreferenceの“Run sketches on display”でディスプレーを選択してshift+Runでいける。参考URL:Window Size and Full Screen
2016年10月28日
■ 眼球運動の無意識な動きを活用する
私が「意識の科学的研究」ってテーマを強調するときは「意識経験について説明できるくらいに脳と心と体の仕組みを明らかにすべき」って意味であって「無意識と比べて意識を研究するべき」って意味ではない。そもそも盲視(無意識の視覚)を研究してるんだから。
僕自身二十歳の頃から視線インタフェースには親しんでいて、視線でプレイするブロック崩しとかも作ってきたけど、健常者向けの意識的な「視線入力」には懐疑的。ただ、視線計測に基づくインタラクションには様々な可能性は広がっていると思う。— 稲見昌彦 Masahiko Inami (@drinami) 2015年6月7日
マウスみたいに意識的に目を動かして入力に使うより、無意識な目の動きをインタラクションに使うほうがいいんじゃあないかって思ってたんで、すごく合意できる。
マイクロサッカードは計測精度が必要なのであご台で頭固定しないといけないのが難点。でもいまアイトラッキングはかなり注目されていて、どんどん安くなってきているし、キャリブレーションも一点でよくなっているし(ゴーグル型で眼と二つのカメラの位置とが固定していることを仮定できるため)、面白くなってきそう。
いま考えているのは以前作った2つを組み合わせてやるといいなというもので、「Proccessing + SuperCollider + Tonnetz」(ブログ記事)
では、マウスでクリックした位置をprocessingで処理して、SuperColliderに送って鳴らしていた。
それから生理研一般公開2014のときに作った「Eye-tracking for dial tones」(ブログ記事)
ではアイトラッカーを使って視線を1か所に保持すると音を鳴るというものだった。この両者を組み合わせてやればいいなと思ってた。
でもって、このプッシュホンのやつみたいに鍵盤を押すように意図して音を出させるやつだとマウスを使うのと大差ないので、普段の自発的なサッカードをそのまま使って音を鳴らしてやればいいんじゃあないかと考えてた。ヒトは普段一秒に数回サッケードする。その大きさは5degくらいをピークにロングテールになってる。この特性に合わせてうまいこと音が鳴るようにしようとか思ってsupercolliderいじってたのだけれど、なかなか進まない。
つまり、fixationしているときに音を鳴らすんではなくて(それだとマウスでポチポチと同じ)、サッカードしているときのvelocitityとかを取ってくる。そうするとイベントが断続的になるのでattack/decay/sustain/releaseを効かした音を重ねてゆけるっていうアイデア。
(追記20161028) けっきょくまだ計画中のまま。
2016年10月08日
■ 盲視では「見えてる」? それとも「観えてる」?
(2015年5月のtwitter書き込みより再構成。)
Sci Reps論文のプレスリリースで、盲視についてどういう表現にするか思案中。これまでは「見えないのに無意識に見える」盲視とか書いてきた。なんかもっとうまい言い方はないか。「見えてないけど、観えている」盲視とか。なんかオカルトっぽいな。ボツ。「視えてないけど、判ってる盲視」なんかダメ。見る、観る、視る、識る。判る。分かる。
トヨタ生産方式とかの文脈で「見える化」と「視える化」の使い分けの例とかを見つけた。なんか造語でもするか。
逆か。「観えてないけど、見えている」盲視。こっちはなぜかオカルトっぽくない。ふしぎ!
ここ見ると元のやつでいいかもって思う。「「見る」->視覚によって、物の形・色・様子などを知覚する。「観る」->判断を下すために、物事の状態などを調べる。(三省堂提供「大辞林 第二版」) 」URL:「「見る」と「観る」」
ベイトソンが引用していたウイリアム・ブレイクの「賢者は輪郭を見る」、これは「観る」だろう。”Wise men see outlines and therefore they draw them”
原文を見るとこれと並置されて"Madmen ..."という表現もあることを知った。
When Men will draw outlines begin you to jaw them
Madmen see outlines & therefore they draw them
ブレイクの”outlines”への言及はここにもある:”Nature has no Outline, But Imagination has.” (The Ghost of Abel)
とここまで潜ってはみたものの、これをどう活かせというのだろう?
といろいろやっていたのだけど、けっきょくプレスリリースは「見えてないのに分かってしまう」盲視はヒトでもサルでも同じという表現になった。
2016年09月22日
■ 研究メモ: 主にサリエンシー関連 (20150429まで)
MMNというのは本質的にはoutlier detectionであって、視覚でいうサリエンシーと同様注意に関わる現象である、という言い方をすると誤解が生じることがわかった。事象関連電位の人にとってはMMNはpreattentiveで、P300がattentiveだから。
つまりこれは注意の研究の二つの流れの話(トリーズマン方式のpre-attentive vs. attentiveとポスナー方式のbottom-up attentionとtop-down attention)を混同しないようにする、というのがポイントだったのだ。
より正確に言うならば、視覚にも聴覚にもサリエンシーはあって、視覚での空間的なサリエンシー(サリエンシー・マップ)+時間的なサプライズ(ベイジアンサプライズ)、聴覚での空間的なサリエンシー(まだ充分モデル化されてない)と時間的なサプライズ(time-freqで計算される)とがある。
それぞれの時間、空間特性の違いがあるということ。そしてMMNに記憶の要素があるというのは、あくまで自動的な過程であって、そういう意味では視覚でのIORに記憶の要素があるのと対応している。
あとは視覚だと目が動いてその履歴に依ってmotorのIORが出てくるわけだけど、P300でやっているようなselectionのときにはselectionの行動自体が顕在化しているわけではないので、sensoryなIORに対応したものがあるはず。このへんまで考えてみると、視覚でいうovertなIORとcovertなIORの関係が聴覚ではどうなっているんだろうか、とかその辺に興味がわく。
いやいや、IORとtemporal surpriseはべつものなのでそれを混ぜるとわけわからなくなる。そこは取り消し。以前やったシミュレーションの結果を使って、聴覚oddball課題の視覚アナログ(赤丸->ブランク->赤丸->ブランク->青丸)とかを計算してみればよいのだな。
以前の@ksk_S さんとのやりとりを見なおして"The entropic brain"を読んでたら、ここでもサリエンシーネットワークが出てきて、SOCとE-I balanceとネットワークのカオスとサリエンスネットワークとフリストンとオートポイエーシスと解離症状と精神症状のサリエンシー仮説とベイジアンサプライズとIIT ver.3が全部つながって、居てもたっても居られなくなってきた。
今年の駒場講義(6/10 「意識の神経科学:「気づき」と「サリエンシー」を手がかりに」)で部分的にでも盛り込んでみたいけど、自分で手を動かしてシミュレーションするか、相当深く論文読んでから出さないと迫力出ないので、もう少し練っておく。
去年も同じようなこと言ってたけど、けっきょくactive inferenceの図を作るところまでだった。自分のブログ記事見なおして思ったけど、IITの勉強はぜんぜん進んでないな。
Change deafnessという現象があることを知った。Change blindnessの聴覚バージョンということだな。ふつうに考えたらサッカードによるchange blindnessはvisionのstabilityを作るために貢献しているものだろうから、change deafnessにおいてはサッカードは寄与しないように思える。auditoryでは関係なさそうだ。いっぽうでもしchange blindnessとchange deafnessの両者に上丘が関わっているのなら、change deafnessにおいてはッカードが寄与するということがあってもよい。
さらに説明文を読んでみると、“Conscious change detection was correlated with increased coupled responses in the ACC and the right insula”とある。おー、ここでサリエンスネットワークとつながった!イイね!
会議だとポンチ絵をそのまんま貼り付けたようなスライドが出てきて、手抜きで悪だなと思うけど、あれはあれで意図がある。ああいうのは説明者は全部説明しないし、オーディエンスは話半分で聞きつつスライドと手元の資料をざっと見して自分に関連する部分に質問するとかそうやって使うもんなんだろう。
分野間連携的な場面で発表するとどうしても「このようなことを言うのは釈迦に説法ではございますが」と言いたくなるが、本当に釈迦に説法だったらそもそもそんな説明入れないし、「いましゃべっている文脈でこれまでによく知られていることを捉え直します」という意味なら言い訳せずに説明したい。ほんとうにできているかと言われると自信はないけど。
最近だんだんその種の会議に参加するようことが増えてきて、忙しい先生方が手抜きしつつもツボははずさないで説明する場面を見ることが増えてきたので、自分自身のプレゼンの作成方針や指導方針などとのすり合わせを意識しながら考えたことをちょっと文章化してみた。
EnChromaがどうやってprotanopiaを補助しているかというと、この会社のサイトに詳しく説明が書いてあるけど、L錐体とM錐体との感度の差が大きい波長だけを通すフィルターを使っている。
だからL錐体とM錐体の感度が完全に重なっている人だとこのメガネは効果がないが、そういう人はprotanopiaのうちの20%以下だとしている。あくまでフィルターだから、短波長の色を提示したときの経験を超えることはないわけで、たとえば私がこれをかけてLCDモニターを見ればCIE空間上の三角形よりも広い(でもCIEの閉曲線のなかを超えない)ところの色を経験することができるはず。
以前DKL空間とかで計算したときの方法を使えばDKL座標での値を計算することができるだろう。ここで議論されてる。
2016年09月15日
■ 研究関連メモ(20150225-20150409)
ウィキペの「平沢貞通」の項目を読んでいたら、平沢貞通が狂犬病予防接種の副作用によるコルサコフ症候群に罹患していて、それによる作話と逮捕の関連についての記載が出てきた。コルサコフ症候群の説明のためのエピソードとしてストック。
ずいぶん前にブログで大学院講義のために記憶障害についてまとめたことがあって、単純ヘルペス脳炎とかウェルニッケ―コルサコフ症候群といった、宣言記憶に特異的な障害(手続き記憶は保持される)とかの例を書きだしたことがある。
どうやらウィキペの記述の元ネタは「小説 帝銀事件」松本清張著(角川文庫)らしい。「小説 帝銀事件」・松本清張著(角川文庫) もう少し直接的な資料に当たりたいが、今日はここまで。
研究についての批判の話だけど、これは文系ゼミで著書への批判をする場面だということに注意するべき。学会での質問とかラボセミナーでの質問とかで学生に向けてこういう言い方をして詰まらない質問ができなくするのは正しくないわけで。
理系のJC発表の指導の場面においては、私なら段階を踏んで説明するだろうか。はじめは発表者は論文の隅々まで(設定したパラメーターひとつひとつまで)徹底的に読んで理解することが大事なので、そのためには重箱の隅でもいいから批判ができるようにencourageする。
引用したブログ記事に書かれていることは次の段階の話であって、そのようにしてブレインストーミング的に思いつくだけ挙げた批判が、その論文の主な主張に影響をあたえるような致命なものであるか評価をした上で、論文の主な主張が充分受け入れられるものなのか、どういう保留事項をつけるべきなのか、
というふうに自分がこれまで読んできた論文や研究を体系化した枠組みの中でいま読んでいる論文の位置づけをフェアに行う。ここで建設的だか破壊的だか、本質的なことが言えるようになるといい読みができているといえるわけで、ってJC指導と論文査読って地続きだなと改めて感じる。
論争的なトピックでどちらかの側に属する立場にいる場合には、認知的歪みでこの位置づけが(ラボ全体ですら)ずれているということはおおいにありうる。
「HMMとspike sorting」みたいな論文を漁りながら、でも究極には、「スパイクソーティングしないで波形データのすべての情報を使って電極近辺で起こっている事象を推定する」のがいちばんいいんではないかと考えた。この論文はデコーディング。
つまり、single-unitへのこだわりを一旦外して考えると、知りたいのはそこにあるニューロンへの入力様式と出力様式とその変換ルールで、記録部位周りにあるニューロンをスパイキングネットワークでモデル化した上で、それぞれのパラメータを推定するのが、ほんとうにしたいことのはず。
芥川龍之介の歯車は閃輝暗点だと思うけど、よく読むと片眼でのみ起こるって書いてる。「僕は又はじまつたなと思ひ、左の目の視力をためす為に片手に右の目を塞いで見た。左の目は果して何ともなかつた。しかし右の目の瞼まぶたの裏には歯車が幾つもまはつてゐた。」網膜片頭痛ってのもあるらしいが。
麻痺する片頭痛 「網膜片頭痛とは突然視界が狭くなったり片方の目が見えなくなったりする片頭痛のこと。目の前にキラキラしたものが見える片頭痛特有の閃輝暗点(せんきあんてん)とは違いますので区別してください。」これ見るとなんかやっぱ違ってそうなかんじ。
閃輝暗点がだんだんfoveaから周辺視野に広がってゆくのと対応して、cortical spreading depressionも後頭葉の最後端から広がってゆくわけだけど、それは一番後ろが血管支配的にいちばん虚血になりやすいからということだろうか。
EyeTribeについてひさびさに調べてみたら、PyGazeのなかにimplementされてる!これでPsychoPyで使えるようになりそう。以前Processing + SuperCollider (youtube)というのを作ったことがあるけど、いまならEyeTribe (-> json) -> Python (-> osc) -> Supercollider でいけそう。ただ、マウスでクリックするのと違って、眼球運動は一秒に数回のサッカードとそのあいだの注視の繰り返しだから、その時間特性に合った音と構成にするべき。
そうやってつらつら考えていたら、そもそもEyeTribeではそんなによい精度が出ないのだから(tobii X2-60でもそれなりに大変だった: Eye-tracking for dial tones (youtube))、キャリブレーションが正確でなくてもよいようなものの方がよい。
そこまで考えると、saccadeやblinkだけ検出すれば良いんじゃ? -> だったらそのときにchange blindnessだ! -> つかそれだったらアイトラッカーである必要すらなくって、EOG(眼電位)で充分だなとか、サッカード時のCBって音にも起こるんだろうか?とかだんだん拡散していったら、ウォーリーを探せ課題でサッカードを検出してターゲットを出したり消したりするような実験パラダイムを思いついたので、前例があるか調べているところ。(<-もはやsupercollider関係なし)
2016年08月28日
■ 「人間は視覚から83%の情報を得ている」の元ネタってどこにある?
『目の見えない人は世界をどう見ているのか』見えないことから見えるもの「「見える人」の中には平面イメージが無意識のうちに刷り込まれている」「「見えない人」にはそもそも「視点」がない」なんかすごく重要なことが書いてあるようなので買う。
@kohske 別件なんですけど、高橋さんはmulti-sensoryに関わってましたよね。よくある「人間は視覚から83%の情報を得ている(音が11%、匂いが3%、触覚が2%、味が1%)」って話の元ネタってご存じですか?ちょっと調べてたらあれってmythかもって気づきました。
@pooneil 私もその話は知ってますが、元ネタは知りません。視覚は1千万bpsという話も同様で、エビデンスはないような気がするんですが、D-mutlisensoryとかで聞いてみましょうか。
— kohske (@kohske) 2015年4月22日
@kohske 似たような話で「われわれはどのsensory modalityを記憶しているか」というのがあるけれど、これもどうやらEdgar Daleの図式をあたかも実験事実のように引用した結果らしいのです。
なんでそんなことが気になったかというと、さっき「目の見えない人は世界をどう見ているのか」を買ってきたのだけれど、いきなり冒頭の文章が「人が得る情報の八割から九割は視覚に由来すると言われています」で、これってどのくらい実験事実に支えられたことなんだろうかと思ったから。
ちょうど自分がやってきたサリエンシーの解析で視覚のfeature (輝度、色、方位、動き)のなかでどれがフリービューイングに影響するかの寄与率を計算してた(正確ではないけど)ので、もしかしたらそれを感覚モダリティーに拡張して寄与率を計算できるのではないかとか考えてた。
(訂正) @kohske たとえば、WikipediaのNonverbal communicationの項目にこの数字は出てくるけど、ここで引用されているのは一般書。
この数字はいろんなところで出てくるけど、たとえば古めの本を探すとこの本のp.321のFig,14-2から14-4までとか。Paul Trattnerって出てくるけど、これの元ネタは見つからず。
教育関係の本に多い印象。これのp.3にあるFig.1-1, 1-2とか。
この本の189ページを見ると、運転中のデータとしてMichael SivakのPerception 1996が引用されてる。これはいっけん有望に見えるかもしれない。
あいにくうちの機関はこの雑誌取ってないのだけど、要旨を読むかぎり、この論文は実験をしたわけではなくてコメンタリ的なもので、「これまでにそのようなデータはないし、そもそもそのような比率を計測する方法もない」という議論をしているようだ。
2016年06月15日
■ 駒場学部講義2016 「意識の神経科学:「盲視」と「統合失調症」を手がかりに」レジメアップしました
駒場学部講義2016 「意識の神経科学:「盲視」と「統合失調症」を手がかりに」レジメアップしました。
今年は神経現象学の話を入れてみた。うまくいくとよいのだけれど。乞うご期待!
- / ツイートする
- / 投稿日: 2016年06月15日
- / カテゴリー: [脳科学メモ] [駒場学部講義2016]
- / Edit(管理者用)
2016年06月07日
■ 静止膜電位はどうやってできるの?
神谷さんのツイートで言及されていた神経細胞の膜電位の話についてだけど、
高校生物の静止膜電位について質問です。静止膜電位は、外側が+で内側が-になる…と習ったのですが、それはなぜでしょうか? プラスイオンが外側に多く、内側に少ないからか?と思ったのですが、専門書を見ると、むしろ内側のほうにプラスイオンが多いです。(Yahoo知恵袋より)
これはかなり根本的な間違いをしている。神経細胞膜の内側も外側も(マクロには)+イオンと-イオンは釣り合っていて、電気的に中性になっている。+イオンと-イオンとが分極しているのは神経細胞膜の近傍だけであって、分極したイオンが膜を挟んで引き寄せ合って分布することで神経細胞膜はコンデンサーとして働く(図1)。
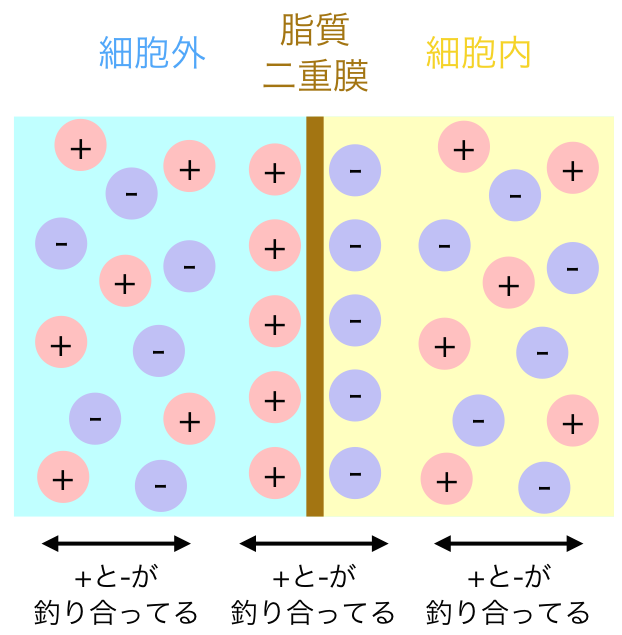
図1 神経細胞膜の内外でのイオンの分布
(カンデル本5ed. Figure6-1を元に作成)
巨視的に見て+イオンと-イオンが釣り合っていないなんて事態はよっぽどのことがないとおこらないということは化学を知っていれば理解しているはずだけれども、でもじゃあなんで電位差ができるのかということはネルンストの式をちゃんと理解しないと説明できない。
「これでわかるニューロンの電気現象」は神経生理学でつまづきやすい部分を対話形式で説明してある名著なのだけれども、この本の第1章はまるまるこの静止膜電位の問題に充てられている。
「キャンベル生物学」を見てみたけれど、たしかに細胞膜の内外のイオン濃度が代表的なものしか書かれていない。これがYahoo知恵袋での混乱のもとだった。じつはカンデル本5ed.(Table6-1)にも代表的なイオン濃度しか書かれていない(図2)。もちろん、Figure 6-1とその説明文章をちゃんと読めば電気的に釣り合っていることは書かれているのだが。
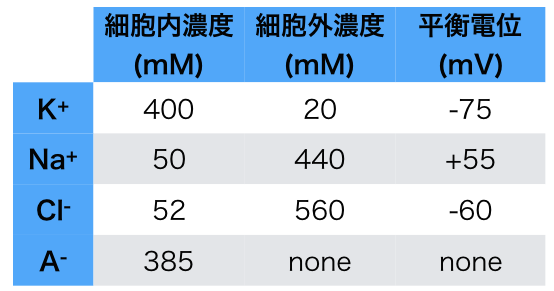
図2 イカ巨大軸索での主要なイオンの濃度
(カンデル本5ed. Table6-1を元に作成)
「これでわかるニューロンの電気現象」のコラム5.1 (p.158)では、陽イオンの濃度の総和と陰イオンの濃度の総和が細胞膜の内外ともに釣り合っている、つまり電気的に中性だということが図で示されている。(ちなみにこの図の引用元はAidley "The Physiology of Excitable Cells"とのことだが、google booksからは該当ページ見つからず。)
じゃあどうやって電位差ができているのかという説明なんだけど、教科書をいくつか読み比べて、どんな説明をしているか調べてみた。
「これでわかるニューロンの電気現象」では「濃度の違うKCl溶液がKイオンだけ透過する半透膜で遮られたとき」という例を使っている。ネルンストの式が濃度勾配による仕事と半透膜間の電位差による仕事が釣り合った平衡状態であることをわかりやすく説明している。
この本の第一章のネタ本はreferenceとして挙げられているとおり、Eckert and RandallのAnimal Physiologyの5章なのだけど、この図を見れば分極は膜の両端でだけ起きているということがよく分かる(図3)。
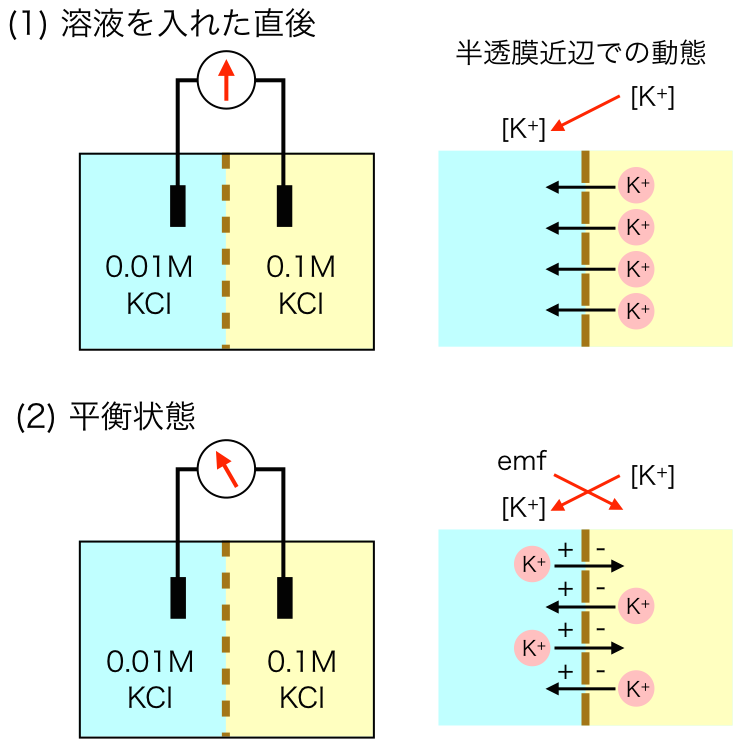
図3 K+イオンの電気化学的平衡
(Eckert and Randall 3rd ed. Figure 5-11を元に作成)
じゃあ、この分極している部分って細胞膜(厚さ5nm)の周りのどのあたりまでか、っていうと案外書いてない。少なくともカンデル本6章にはなかった。私も知らなかったけどWikipediaの膜電位の項目に書いてあった。膜の近傍の2-3nmだそうだ。
このあたりを深掘りするためには膜のチャネルをひとつの抵抗として捉えるような近似では追いつかなくて、膜の中を一定の電場として近似する定電場理論や、さらにチャネル内のエネルギー勾配をモデル化したりする必要がある。このあたりについては標準生理学8edのp.64-65やニューロンの生物物理 第2版のp.22-25に記述があるけどネルンスト・プランクの式(微分方程式)を導入する必要が出てくる。
じつは今回のブログ記事の個人的な伏線は、さいきんラボのOBの方たちと医学部の生理学教育においてネルンストの式をどこまで説明するか、どうやって説明するか、という話をしたことだった。あのあと、自分ならどうするかと考えていた。たぶん、図を多用して定性的な話をするべきで、それでさらに興味を持った者に対してはネルンスト・プランクの式とかを教える、という順番になるのではないかと思った。
(たとえば、ガイトン生理学ではネルンストの式は EK = 61(mV) * log10([Kout]/[Kin]) という表記を使っていて、RT/ZFを見えないようにしてある。しかもTは37度にしてあって、生理学実験でよく使う25度を使ってなかった。このへんはかなり実際的であることを意識していると思った。)
「これでわかるニューロンの電気現象」は著者が岡山大学の理学部生物学科の2,3年生を対象とした講義を元にしたものということなので、医学部の生理学教育を考えるのに良い題材であるといえる。「これでわかるニューロンの電気現象」およびEckert and RandallのAnimal Physiologyでは「どのくらい膜の近傍に限局してイオンが分極しているか」を説明するのに、膜の容量が1uF/cm2が上限であるということを使って、分極しているKイオンの数が細胞内の全体のKイオンと比べると1千万分の1程度であること、つまり膜を透過するKイオンによって細胞内のKイオンの濃度が変わったりはしないという説明をしている。これはわかりやすい。というわけで「これでわかるニューロンの電気現象」は素晴らしいのでおすすめ。(<-回し者)
(ちなみに、この説明を理解してもらうためには等価回路の概念の導入が必要であるので、正確に説明しようするとそれはそれでたいへんだと思う。脂質二重膜の実態に沿った説明と、等価回路を用いた説明をどのようにうまく使い分けて説明するかは、静止膜電位にかぎらず、神経生理学の教育において重要なコツとなるだろう。)
さきほどはカンデル本5edのTable 6-1に文句をつけたけど、どのように説明すると理解しやすいか、という眼でカンデル本を読んでみるとじつにうまく書かれていることがわかる。つか今回これ書くために読んでみて感服した。
カンデル本5edの6章では、Eckert and RandallのAnimal PhysiologyでのKイオンの半透膜の例を持ってくる代わりに、グリア細胞がほぼKイオンのleakチャネルだけである話を持ってきて、ネルンストの式の平衡状態を説明しておく。
そこからの差分として神経細胞の場合を説明している。神経細胞はKチャネルとNaチャネルからなるものと単純化しておく。グリア細胞ではKチャネルのみだった脂質二重膜にNaイオンのチャネルを付加すると、Naイオンが流入するようになってやがて膜間の電流の流れは0になるけど、ゆっくりとKイオンの流出とNaイオンの流入が起こる定常状態(平衡ではない)になってしまう。つまりそのまま放っておくとイオンの濃度勾配はなくなってしまう。それをせき止めるためにエネルギー使ってNa-Kポンプを回す話が導入されている。なるほどこれはステップバイステップでうまく説明してある。
今回この辺りの教科書を読みながら自分だったらどう説明するか考えていたんだけど、まったくゼロから説明するためには、「まず濃度勾配のない海水に脂質二重膜で壁を作って、Na-Kポンプを作って濃度勾配を作って、それからKチャネルを付加して」みたいなことを考えてた。でもこれはややこしくなりそう。カンデル方式のほうがすっきりしてる。
カンデル本ではその後でNa-Kポンプが静止膜電位を数mVほどnegative方向に押し下げるということも説明してある。これもKチャネルだけでネルンストの式を説明した後ならばすぐに理解できることなのでわかりやすい。ということでカンデル本おすすめ。と書くのも烏滸(うーろん)がましいが、あらためてその素晴らしさを実感したのだった。
(じつはこれらの教科書では触れられていなかったが、ニューロンの生物物理 第2版では、細胞内には有機イオン(A-)があってこれは膜の外には出られないのでドナン平衡を考慮する必要があって、これによってドナン浸透圧が説明できるということが書いてある。これは心臓の生理学に関わってくる。)
2016年05月22日
■ わたモテとSAD(つづき)
以前「わたモテとSAD」というブログ記事を書いたことがあるのだけど、そこで言及したKOTAKUの記事を再読してみた。
Kotakuの記事は「わたモテはSADを嘲笑していて許せない」という論調だが、それには同調できなかった。なぜなら著者の二人がSADもしくはそれに近い立場であっただろうと想像できるからだ(じっさい、何巻かのあとがきにそのような記述がある。)。そういうわけで私にはSimone86のコメントがいちばんしっくりきた。
つまり「SAD当事者にとっては『こういう経験は自分だけのものじゃないんだ』といったセラピー的効果がある。」「一方で、この番組を評価している人がみなこの苦痛を経験していると考えたとしたらそれは間違いであることも確か」(超訳的まとめ)
じっさいSADと診断された人でなくても、日常でそういった経験をすることは(私を含む)多くの人にもあるわけで、だからこそそれを誇張した感じで描かれるのを、どこか自分の一部として読み、どこか突き放して、苦いユーモアとしてこの作品を読むことができるのだと思う。
たぶん、そういう経験を共有していない人には「この作品のどこが面白いのか、さっぱりわからない」ということになるんではないかと思う。(逆に感情移入しすぎて、突き放して見れないというのもある。後述。)
じつのところ、環境次第でだれでもそういう状況は起こるのではないだろうか? 私の例を挙げると、海外のラボに行くと言語の壁で雑談とか全部は聞き取れないから、周りが笑うのに釣られて笑ったり、いきなり自分に話題を振られて(「マサトシ、日本ではどうなんだい?」)、キョドった応答をしてしまったりとか。
海外での経験に関してはこちらにも書いた:「ある闘いの記録」
最近は、次男のソフトボールチームの関係でお母さん方と会って話する機会が増えているのだけれども、共通の話題がソフトボールのことしかないから、これがしんどい。ソフトボールチームの忘年会に行ったら3時間、ほとんど面識のない人たちと飲むことになって、はじめの一時間は頑張って話題探して会話したが、後半はもう誰にも見られないところでスマホいじってた。あれは辛かった。あのとき俺はまごうことなきボッチだった。
大学生くらいの頃に弁慶かどこかのラーメン屋に行ったときに、注文を飛ばされたらしく30分くらい(<-誇張?)ずっとラーメンが来ないで放っておかれて、泣きたい気持ちになってそっと席を立った経験がある。先日栄の吉野家では5分近く放置されたので店員に声をかけて注文した。俺、成長したよ。
わたモテに関しては私はどちらかというと、主人公に感情移入しすぎて読んでいて辛くなる、という意味で楽しめないというところはある。それはTogetterの「"恥をかくシーン"が苦手な人たち」で話題になった「あの感覚」とたぶん同じことだと思う。
この感覚を指し示す言葉として、vicarious embarrassmentという概念があるそうだ。こちらの記事が詳しい:「あの恥ずかしい気持ちには名前があった」 さらに脳機能イメージングの研究もあったりする:Your Flaws Are My Pain: Linking Empathy To Vicarious Embarrassment
2015年10月30日
■ 研究発表などのプレゼンについて雑記
学会とかでの研究発表の場面の話だけど、レーザーポインタにしろマウスにしろ多用し過ぎると、体が観客に向かわずに画面に向いてしまう。だから複数の図をひとつのスライドに入れる必要がある場合は、アニメーションで順番に情報を追加するようにしている。こんなかんじ:
でも、複数の図をひとつのスライドに入れずに済むのなら、なるたけ分けたほうが良い。ジャーナルクラブでNatureとかScienceの論文を取り上げるとコンポジットの図が多用されるので、必要ない限り別のスライドで出すように分けて、その分図を拡大して出すように指導している。グラフの軸の数字が読めないとかそういうのはよくない。
複数の図をひとつのスライドに入れる必要があるのは図を並べて比較したいときだけ。たとえば上のスライドの場合、サリエンシーの高いところを元画像と比べたいという目的があるので並べて提示している。
もしアニメーションの使用を徹底するとこんなかんじになるだろう。
さすがにくどいんでこれは後者は本番のときには使ってない。あんまアニメーション多用しすぎるのもたぶんよくない。オーディエンスの注意をコントロールするのは良いのだけれど、ずっとスライドに注意を向けていないと話に追いつけなくなる。そしてわれわれは人の話を聞くとき、ずっと100%集中しては聞いていない。
以上はプレゼンを作る側としての考え方だけど、じゃあオーディエンスとしてはどうかというと、よく知った分野のことだったら、さっさと全部の図を出してもらって、それをざっと見回してスピーカーがこれから何をしゃべるつもりなのか予想した上で、スピーカーの説明を聞いているかもしれない。
たぶんオーディエンスにそういう自由というか余裕を与えておくことは重要で、100%聞いてなくても理解してもらえるようにするために途中でまとめを入れるとか、そういうことが必要になる。それもあんまやるとくどくなるのだけど、指導の場面ではくどい方に寄せておくようにしている。
P.S. ちなみにここで私が言っている「アニメーション効果」とは「出現」と「消去」のことだけを指している。あくまでオーディエンスに見せる情報をコントロールする手段として使っている。スライドの切り替えでなんか面白い効果を使ったりすることには興味が無い。端的に時間の無駄だと思ってる。ただし、瞬間的なスライドの切り替えだと、瞬きによるchange blindnessが作用する可能性があるので、同じようなスライドが続くときにはスライドの切り替えに気づかなくなる可能性はある。そういうときには切り替えの効果も役に立つのかも。
2015年08月10日
■ 脳と非線形力学系について勉強してますって話
Mark ChurchlandのNature 2012が出て、リーチング運動時の運動皮質ニューロンの集団での発火のダイナミクスが、非定常的なものではなくて周期的なものとして捉えることができる(がゆえに力学系的解析が応用可能になる)というのを読んですごくこの辺りに興味をもつようになった。
Vreeswijk and SompolinskyのScience 1996 (PDF)での「興奮性ニューロンと抑制性ニューロンがバランスしたネットワークでカオスが発生する」という論文では、かなり単純なニューロンモデルが扱われていた。
それに対して、ここ最近ではRenart et alのScience 2010やTan et alのNature 2014にあるようにEIバランスによるasynchronous stateがげっ歯類のin vivoでの仕事で報告されるようになってきていて、もはやこのあたりは流行りと言ってもよいかと思う。
そういうわけで非線形力学的な道具を使って脳活動を解析するというのに興味を持って、遅ればせながら勉強をしている。
埋め込み定理を使って脳波とかの時系列データを解析するってのは合原先生の1990年代の仕事とかで見たことがあったけどそれ以来あんまり見ないから、なんらか実際の運用上で難点があって一般化しなかったのかなと推測していた。でも前述のChurchand論文とかを見ていると再訪する価値があるかもとか思ってた。
そしたら最近、Pouget研の田嶋達裕さんが藤井さんのneurotychoのデータ(ECoGでの多点同時記録)を使って、脳活動の埋め込み関係から因果構造を捉えるという仕事に取り組んでいる。「生体の科学」のレビュー(PDF)
これはSugihara et alのScience 2012を踏まえての仕事らしい。こちらも読まなくて、と思ってた。
そうしたら先日の「因果フェス」でコメンテーターとして参加した伊庭さん@ibaibabaibai の8/7の連続ツイートでSugihara et al(のCCMに関連した講演)が批判的に取り上げられてた。
このあたりの状況に追い付きたいのだけれども、勉強不足でまだそのずっと手前の位置にいる。(イズケビッチ本のFHNニューロンの相空間解析とかさいきん翻訳が出たストロガッツの「非線形ダイナミクスとカオス」とか読んでる。つまり、まだ学部レベル。)
こんなことを「自分で」する意義があるのだろうかと自問自答しつつも、でも自分が次の段階に行くにはたぶんこれが必要なのではないか、とか考えているところ。自分語り乙。
2015年07月25日
■ 研究関連20140930
「エモーショナル・デザイン」ってのも借りてきた。けっきょく「感性」がなんだかよく分からないのは感受するものとしての感性と同時にデザインするものとしての感性工学とが混ざり合っているからだろうか。この「情動に訴えかける」デザインってのは意味が分かる気がする。
でも今度は「情動」って言葉がよく分からない。とくにemotionの訳に情動を使われるとさっぱり分からない。「感情」以外に「欲求」とかそういう要素が入るってことは分かるんだけど。"emotion"って語にはそんな含意はないよね? と思ってみてみると、ウィキペで"feeling"のことを"the conscious subjective experience of emotion"(APA Dictionary of Psychology)と定義してあって、だんだんわからなくなる。
emotion http://en.wikipedia.org/wiki/Emotion はfeelingとmoodとaffectと区別している。 affectって言葉がいちばん「情動」によく対応しているように思う。
"Affect is an encompassing term, used to describe the topics of emotion, feelings, and moods together, even though it is commonly used interchangeably with emotion."
TPJアナログとしてのSTG, STP, TPOあたりについて調べていた。このへんは場所的に視覚、聴覚、体性感覚が収束するという意味で重要なのだけれども、前庭系(PIVC)がここにあることをずっと気にかけていなかった。やっぱなにもかもここに集まってるな。
マカクでのPIVCの場所についてはDora E. Angelakiの論文で議論されている。
半側空間無視への介入としてカロリック刺激ってのがあって、まさに前庭系に刺激を加えているわけで、前庭系の話はとうぜん重要な要素。
解剖学やってた経験がfMRIの活動とか見るのに役立ってる。やっぱ全脳切ってニッスルで染めて、coronal, axial, sagital全部見てみるってのを導入に使うとよさそうだ。
「脳腫瘍摘出手術後の視覚障害の実際(視野障害・半側空間無視)」 ご自身の経験をブログでくわしく綴っていて、これはものすごく重要な資料だと思う。
「ちなみに、見えていない部分は「真っ黒」になっているわけではありません。単に見えないだけです。みなさんも頭の真後ろは真っ黒になっているわけではありませんよね。それと同じで、単に視界の外になってしまいます。」私が講義で使う説明と同じだ!
「脳には「見えない部分を勝手に補完する」という機能があるため(錯視等で有名ですね)、見えていない左下の領域も、それ以外の見えている領域(左上や右下)の映像から類推して、勝手に背景映像を作り出してしまいます。」この事実は盲視が見つかりにくい理由かも。
追ってブログ読んでみたら、グリオーマの摘出手術のあとで白血病・悪性リンパ腫で現在闘病中ということを知った。
盲視の患者で視覚的補完が起こることについてはMarcelのBrain 1998(PDF)が詳しい。実験5(図5)が視覚的補完の実験で、左右丸だとうまく補完できるけど、ゲシュタルト的に揃ってないものは補完できない。
来年のASSC19はパリでSid Kouiderがchairだけど、サテライトでKevin O’Reganがサテライトをやることを知った:The Sensorimotor Theory of Consciousness
The Sensorimotor Theoryに関してはわたしはproponentだけど、Kevin O’Reganの考えは極論に走っているので同意しない。盲視での研究経験から、「なにかあるかんじ」はSMTで説明できるかもしれないけど、それがfull-fledgedな意識経験になるにはまだ何かが足りなくて、それが腹側経路にある、という考え。
背側経路はprimitiveな意識経験の鋳型を作るのには必要で、生まれてから知覚運動連関の経験がないと意識は作れない。でも夢や植物状態でも意識経験はあるように、いったんその鋳型ができてさえしまえばオンラインでの行動は不要。ここで言っている鋳型というのが腹側経路で言うpriorのことであり、top down predictionのことになる。そしてそれはいきなり腹側経路で作られるのではなくって、発達期に上丘を経由した短絡路で作られる(発達期の顔応答の知見 see MH Johnson)。
だから、Deep Learningの話で、ここのレイヤーを学習させるときに短絡路があると効率が良いという話を聞いて、これだ!と思った。まだ勉強が足りないのでアイデアだけだが、subcortical face processingの経路でやっていることが学習理論にうまく組み込めてそれの実験的証拠みたいなのが作れたらインパクトがあるんではないかと思ってる。
なにかあるかんじ、と鋳型のあたりはもっといろんなことが今の段階ですでに推測できる。Visual agnosiaの人は色のモザイクが見えて、形が見えなくて、でも「空間」は持ってる。空間ができることと一人称的視点の形成と知覚運動連関の形成とはたぶん同じものだろう。
2015年04月21日
■ 低血糖無自覚 (hypoglycemia unawareness)
生理学実習関連で教科書とか読んでるんだけど、どうにも気になったのが「低血糖無自覚」って言葉。
糖尿病でインスリン療法をしている人ではインスリンが効き過ぎると低血糖発作(昏睡、痙攣)が出てしまう。ふつうだったら発作が起こる前の段階で低血糖特有の認知的応答(飢餓感、動悸、発汗)が起こるんだけど、この認知的応答になんらかの理由(インスリン投与に対する脱感作とか)で気づかず(unawareness)に低血糖発作まで行ってしまう。これが「低血糖無自覚」。だったら「無自覚性低血糖」という方が正しいだろ!と思って検索してみたら「無自覚性低血糖症」って言葉もよく使われるらしい。となるとけっきょく元の英語の表現("hypoglycemia unawareness")自体が悪いのだなということが分かった。
そもそもいったいなんで名詞が二つ並んでるんだよっていう。とりあえず元論文を遡ってみた。PubMedによると1982年に使用例があるが、メジャーっぽい雑誌ではこれあたりか:Endocr Rev. 1991 Nov;12(4):356-71 案外新しい。あとHAAF(hypoglycemia-associated autonomic failure)という表現も知った。
脱線したけど、メカニズムはなにかというと、これ見てなるほどいろいろ複雑そうだと思った。Defective counterregulation and hypoglycemia unawareness in diabetes
けっきょく繰り返し起こる低血糖のエピソードによってさらにそれに体が対応して、ということが起こっているので、こういう来歴のネットワークにどう対処するかというのが問題なわけで、対症療法では厳しそうだ。
以前合原先生の講義のときにちょっと読んだ、前立腺ガンのホルモン療法のモデリングとかを思い出した。こういうのがすごい重要だと思うのだけど、本気で医療の現場で使うのはたいへんそうだ。
Varelaの論文とかでてんかん発作の予測の話も読んだけど、あのへんは進歩しているのだろうか?
そんなことをつらつら考えた。
2015年03月28日
■ 研究関連20140731
「claustrumを電気刺激したら意識を失った」の報告のソースはこれか:"Electrical stimulation of a small brain area reversibly disrupts consciousness"00201-7/abstract?cc=y)
claustrumは非常に平べったい領域で、外包と最外包に囲まれていてすぐ外側がinsulaという場所なので、どのくらい正確にclaustrumだと言えているか要確認。要旨でも電極は左claustrumとanterior-dorsalのinsulaの間って書いてあるし。
延期しまくってたthe EyeTribeのMac用SDKだけど、ダウンロードできるようになってた。しかしメールとかでアナウンスなし。とにかく動作確認した。とりあえずはマウスの代わりに使ってProcessingで動かせばなんとかなる。
次の問題はもうすこしインタラクティブなことをするのにどうするかだが、GitHubにいくつかサンプルがある。Unityをいじるかどうか思案中。Processingと通信するためにはJSON使えばいいのだなとか単純に思っていたが、「コマンドラインとブラウザで JSON API を手軽に試す」を参考にして、ためしに curlでhttp://localhost:6555 とかやれば読めるのかと思ってやってみたら、なんにも返ってこなかった。
なんかストリーミングしているデータを読むときの流儀ってもんがあるらしい。そういうわけで、けっきょくUnityかC#のサンプルファイル読んだほうが近道っぽいということが判明。
Humanのvon Economo neuronちょっくら探してみるかとか思って、Allen Brain Atlasとか見てるけどそんなに簡単に見つからないな。ちょうどいい切片がないってのもある。側頭葉と切り離されたくらいにanteriorで前障が曲がっているあたりのはずだが。
例のclaustrumで意識のon-offスイッチの話だけど、解剖学的にはextreme capsuleを越えるとまさにVENのあるところだってことが気になる。こっちのほうが本命なのでは?
これの"Nearest Reference Image"で見つけた。やっぱventral surface側でgranularなところというのがミソか。あと右側に多いというのも読んだ。
claustrum自体は前後に広いので隣はかならずしもfront-insular cortexではないけど、VENのあるところには必ずdorsalにclaustrumがある。それはhumanでもchimpanzeeでもmacaqueでも。
marmoset-brain.orgでNADPHdを見たら、claustrumにすごく大きいニューロンがたくさん見えるので心躍った。Humanで論文が出てる:Topographical distribution and morphology of NADPH-diaphorase-stained neurons in the human claustrum
"Biological insights from 108 schizophrenia-associated genetic loci" このへんそろそろ勉強しないといけない気がしてきた。ドパミン説とグルタメート説とをどう繋ぐかとか。
Geneticな解析で出てくるのはたいがいNMDAだったり、Caチャネルだったりして、素因の部分はgluなんだろう。海馬とかWMとか前頭葉の萎縮とかあのあたりから攻めるのは得策ではないだろうと考えていて、症状発言の段階でのサリエンスとかあのあたりに取っ掛かりを感じたのだった。
だから両者をなんとかつなぐようなことを考えて、PPIとかMMNとかの現象論ではなくって、素因から症状発現までの経過のメカニズムを説明するようなことを生理学的に回路の方から攻めることができる宛てがあるのなら、このへんの参入する意義はあるのではないかとか思ってた。
10%が「機能する」のか、10%が「活動する」のか、「最大能力の」10%を出力するのか、というので大違い。そしてそのへんを意図的に混同させて使うところがミソ。活動していないニューロンも活動していないことがシグナルとなり、機能に寄与している。
10%しか「使われて」いない、という場合はどうだろう。「使う」というのがread-outされるという意味なら、持っている情報のほんの一部分だけが行動に現れていて、もっと多くの情報は無意識下で処理されている、という事態を圧縮して「10%しか使われていない」と表現…やっぱダメか。
「10%神話」はそれをとっかかりに脳の自発活動の話とか、脳の情報をどう評価すればよいのかとか、意識的なアクセスとは何かとか、運動の強度によってどのくらいのニューロン活動がリクルートされるかとか、そういう話題を授業で振るマクラにできるので、ネタとしてストックしとく価値はありそう。
「仰向け」になってください、と言われるとどっちのことかピンと来ないことがある。「仰向け」<->「俯せ」で対になっていると覚えればよいのだろうけど、「腹這い」の反対語は?とかなる。
supine <-> prone も取り違えるので、回外と回内のどっちがsup/proだっけとなる。この記事を見て納得行った。前へならえの姿勢から手のひらを上に向ける(回外)がsupinationで下に向ける(回内)がpronation。
眼球の運動ではtorsion(回旋)という向きがあるけど、これをやる上斜筋は筋肉を滑車みたいなのに引っ掛けて回旋の向きを作ってる。これ見るたびに、デザインした人(大いなる自然?)すげーと思う。なんJ的に言えば、おかしなことやっとる。
{kind=link}
同時確率P(x,y)を周辺化(marginalize)してP(x)を計算するときには積分をしているけど、経済学の限界費用 marginal costでは変化量を見るために微分している。marginalって同じ言葉を使うからには共通の要素があるんだろうけど逆に見える。よくわからん。
Marginal distribution ここ見たら、同時確率のテーブルを作ったときにそのマージンの部分に行、または列の総和での確率を書くから、と書いてあった。なぜだろう、がっかり。
「「脳だけになって生きている」モチーフの元祖ネタってなんでしょうか?」 これはなんかの機会にネタとして使えそう。メモっておく。
関連してwikipedia「斬首後に意識はあるか」
2015年02月26日
■ 「状態空間モデル入門」講義に出てきた
明日総研大の大学院講義で「状態空間モデル入門」小山慎介(統計科学専攻)というのがあるので勉強してくる。島崎さんのPLoS Comput Biolとか理解できるようになることを期待して。
予習中。「神経科学と統計科学の対話3」State Space Methods in Neuronal Data Analysis (Z Chen) パート1及びスライド。動画はパート2およびパート3まであり。
「状態空間モデル入門」講義参加してきた。あいにく出席者が少なくて残念なかんじだったが、おかげでバンバン質問してマンツーマン的に教わることができた。講義としてはまず確率的因果推論とマルコフ過程の概論から。
確率的因果推論っていうけど、以前ブログで「ラットの因果推論」について採り上げたときに、原因の確率を手計算していたけど、あれがまさに確率的因果推論であって、PRML8章の「グラフィカルモデル」というやつのこと。たとえば遺伝の話で言えば、メンデルの法則っていうforwardモデルがあるから、先祖から子孫で表現型がどのような確率で伝わるかわかる。ベイズの法則を使えば逆向きの推定ができて、ある子孫の表現型から先祖の表現型の確率を推定できる。
マルコフ過程のほうはPRML13章の「系列データ」に出てくるあれ。状態量x(t)が直前のx(t-1)だけで決まる。隠れマルコフモデルでは、計測値y(t)と見えない状態値x(t)とに変換式P(y(t)|x(t))があって、x(t)がマルコフ過程になっている状態空間モデルの特殊例(取り扱いしやすい例)だということですっきりまとまった。
ベイジアンのグラフィカルモデルでは有向でループのないツリー状構造が扱われるのに対して、マルコフ過程ではx(t-1)だけでx(t)が決まるという、枝のない特殊例であるということも理解した。
カルマンフィルターも状態空間モデルの特殊例で、隠れマルコフモデルが離散的であるのに対して、カルマンフィルターは連続的であり、ノイズがガウシアンであるという仮定が入ってる。
隠れマルコフモデルでは、いったん計測値y(1:t-1)が入手できれば、現在の状態値x(t)の確率密度分布p(x(t)|y(1:t-1))を計算するのにチャップマン=コルモゴロフ方程式を使ってやればよい。
M1ニューロン活動から運動の方向をデコードするような例の場合、ニューロン活動も運動も自己相関が高いので運動の方向は急には変わらない、つまり自己相関が高いのでマルコフ過程の仮定を置くことができて、デコーディングがうまくいく。
実際にチャップマン=コルモゴロフ方程式を使う際には
p(x(t)|y(1:t)) ∝ p(y(t)|x(t)) * p(x(t)|y(1:t-1))
でベイズの公式を使う。規格化するために分母を計算しないといけないのでパーティクルフィルタとか使ったりとかいろいろテクがある。講義では省略されてたけど、ここが難しいし、時間がかかる部分であることはわかる。
けっきょく状態空間モデルでは見えない状態x(t)を計測値y(t)から推測するためにベイズ推定を使うので、x(t)からy(t)を生成するモデルがうまく作れないとダメな推定を行ってしまうことになる。
後半の講義では、小山さんのJ Comput Neurosci. 2010 Comparison of brain-computer interface decoding algorithms in open-loop and closed-loop controlを元にした話をしてた。
ここでは、デコーディングをする際に、1) population vector => 2) ordinary least squares => 3) nonlinear least squares => 4) loglinear least squares => 5) state space model とモデルを変えるごとに拘束条件をゆるめてより自由なモデリングができること、そしてこれらのなかでM1ニューロンのデコーディング(Andy Schwartzのデータを利用)にはどの拘束条件が効いているかを検討した話を聞いてきた。
結論としてはopen loopのデコーディングでは「サンプルしたニューロン集団のなかでpreferred directionが一様に分布している」という1)での縛りが聞いているのだけれど、closed loopでは動物が学習してくれるのでこの縛りは重大でなくなる。それでも5)でadaptiveのノイズのスムージングをしてくれることがデコーディングの性能に効いているという点ではclosed loopでもopen loopでも変わらん、というものだった。
これはモデル推定のパラダイムだから、たとえば状態空間のモデルとして複数のものを作って、それらからより良いモデルを選択するということも可能。
状態空間モデルは自由度が高いので、島崎さんの論文のように高次相関を状態空間モデルに組み込んでやれば、時々刻々と高次相関が出たり消えたりする(セルアセンブリの形成)のを推定することができる。多細胞同時記録神経スパイク時系列データの状態空間モデルおよび動的スパイク相関の状態空間モデル やっとこのへんわかってきた。
状態空間モデルでは、データの追加にともなってそのつどベイズ更新をしてゆくから、たとえば電極埋め込んで長期間デコーディングをしてゆくときとだんだん記録が悪くなっていくのだけど、そのときにデコーダーを逐次アップデートしてゆくことができるわけで、それはよさそう。ざっとググってみたかんじタイトル的にこの論文が該当するか:Neural Comput. 2011 "Adaptive decoding for brain-machine interfaces through Bayesian parameter updates."
状態空間モデルがうまくいけば、状態量 x(t-1) -> x(t) -> x(t+1) といった推移を推定できるわけで、究極的にはそれがスパイキングネットワークモデルでの推移則を決めるための拘束条件にできればいい。こうすると力学系的アプローチにつながる。夢見過ぎだろうか?
それはそれとして、時系列の解析で見えない過程から測定値への変換のモデルを作って、測定値から見えない過程を推定する、というのはまさに生理学者がやりたいことそのものなので、いろんなところに使えるということがわかる。
ニューロン活動だけでなくって、サリエンシーに基づいた視覚探索の場面でも使えるだろう。サリエンシーにもとづいてどのように目を動かすか、IORとかattention apertureとかいくつかパラメータを作って、モデルを作ってパラメータフィッティングとか最近やってるんだけど、これってまさに状態空間じゃね?
とか思ってふと考えてみたら、NTT木村さんがやってたのってまさにベイジアンな隠れマルコフモデルによるアプローチだった。A stochastic model of human visual attention with a dynamic Bayesian network いまのいままで繋がってなかったomg!!! ということでなにすればいいか判明した!
- / ツイートする
- / 投稿日: 2015年02月26日
- / カテゴリー: [Saliencyと眼球運動] [ニューロンの情報コーディング] [脳科学メモ]
- / Edit(管理者用)
2015年02月25日
■ 研究関連20140630
多重共線性が出たときの対処法って結局どうすればよいのだろう。Y = b1*X1 + b2*X2 + e でX1とX2が相関高いときにX1もしくはX2だけに変数をdropしてregressするのは正しくないと思う。(X1+X2)と(X1-X2)という二つでregressするほうがよくないか?
Kindle入りするまで待つつもりだったちくま新書の哲学入門(戸田山和久著)を買ってしまった。まだ2章だけど、買って正解、ものすごく役に立つ。ミリカンの議論に基づいて表象とは何かという話から始まるのだけれども、「因果的に決まる限り表象は間違え得ないけどそれではおかしい」というところで、下條先生の「「意識」とは何だろうか―脳の来歴、知覚の錯誤」でのNewsomeのperceptual decisionの議論を思い出した。こういうことを知りたかったんだった。
あと、「意味を作るためには中国語の部屋だけではダメで行動が必要」って議論があって、分析哲学でも行動の意義は考慮されてるのであって、enaction説とそんなに断絶してるわけでもないなと思った。さらに読み進めてみることにしよう。
薬学部の頃に「プログラム学習シリーズ」の赤い本、たとえば「有機合成化学」とかがあって、あのシリーズにすごくお世話になったのだけれど、ああいうかんじで神経解剖学を大づかみなところから詳しいところまで教えることはできないもんだろうか。
いきなり「Gray's Anatomy」最初っから読めってのが正しいとは思わない。学部だったら薄めの教科書を一冊とにかく最後まで行くのが精一杯だと思うけど、どうしたものか。和訳本「カンデル神経科学」が出たタイミングなので考えてみた。
大学院の勉強会やるとしたら、カンデル本やってから"The New Visual Neurosciences"と"The Neurology of Eye Movements"だろうか。
数年前に二分冊の"The Visual Neurosciences"からいくつか章を選んで輪読したのだけれども、合冊の新版を見たらけっこう内容が変わってる。もう一回新版でやってもいいのかも。人も入れ替わっているし。
messy mindについてだけど、@fronoriさん、@kazuhisさんが言及しているとおり、これは「機能というものについての進化的な見方」というかなりgeneralな観点なのであって、私自身もこの概念を知ったとき、下條先生の「来歴」やVarelaの"lineage"を思い出した。
だから論点はそれ自体というよりも「脳波や脳波のcross-frequency couplingみたいな直接byproductっぽいものが機能を持ちうる」という気づきを得たことを記しておきたかった。
そしてこれは計算論的脳科学におけるデビッド・マーの「脳活動と行動から計算論的原理を抽出する」という考えに対して、否定はしないけれども、機能とimplementationの関係が一対一のような関係ではなくてもっとmessyでありうることを示唆しているわけで、意義は大きいと思う。
いろんな人に会って考えていたが、左右の脳で半視野ずつ分担しているのに、なんで左右が切れ目無く繋がっているかということは、連合線維と交連線維とが同等であるわけでもなかろうし、けっこう難問なのではないかと思った。
「非線形な世界」の補足ページより。こことかシビれる:「「複雑系」はダーウィン過程によって生成する系あるいはそのような系によって構成された系であり,短時間の過程で生成することは一般にあり得ない.したがって,伝統は枢要である.反動ならざる保守だけが複雑な系を維持しうる.」
脳とSOCとかそういう話題についての記事なのだけど、砂山モデルのPer Bak氏はすでに亡くなっていることを知った。nytimesの記事:obiuaries
「バクはデンマークのもっとも偉大なアメリカ人だった。…普通のデンマーク人は対立を避けるものなんだが、彼ときたら尊大で同僚と論争をすることを好んでいた。バクと合ったことのある人達ならみんなバクとの初対面のときに彼からどんなふうにひどい放言と侮辱をされたかを話すことができるよ。」ウィキペより。
いろんな動物の脳のサイズを比較している図がツイートされていて興味をもったが、出典が書いてない。Google画像検索で調べてみたらFrontierジャーナルのレビューであることが判明した。"The evolution of the brain, the human nature of cortical circuits, and intellectual creativity" ここのFig.7。リンク こういうのをストックしておくとトークの時とかに役に立つ。
{kind=link}
この図を見ると、ヒヒやマンドリルの脳が思ってた以上にマカクにそっくりだったので見方を新たにした。ちょっと彫りは深いかもしれないが、IPS, ArcS, LunSとかほとんど同じだからブロードマンの領野とか即推測できそう。
あと、イヌやネコに中心溝(ローランド溝)がないのってあらためて不思議なことな気がしてきた。それよりも外側溝(シルビウス溝)のほうがより深く、発達の段階でも一番最初に形成される脳溝らしい。胎生10-15週だそうな。(The Development of Gyrification in Childhood and Adolescenceのfig.1より)
autismでのSensory overloadとNed Blockのoverflowって繋げることができるんではないかなとか考えた。
Rodolfo R. Llinásの"I of the vortex"にホヤの話があることを知った。つまり、動いているときは原始的な脳を持っているのだけれども、いったん定住してしまうとホヤは自分の脳を消化してしまうと。「動きが脳を作る」系の話でこのネタは使えるな。
2015年02月06日
■ 駒場講義の準備メモ(2014年版)
毎年6月恒例となってきた、駒場講義について少し考える。このあいだのclinical neuroscienceの総説原稿を元にして再構成する。「意識のcontent」だけではなくてstateとしての意識、みたいなことを考えるのに「前反省的自己意識」の概念が使えることがわかったのが去年からの進歩だけど、いきなり現象学入門ができるほどこなれているわけでもないので、ダマジオの中核自己みたいなもの、いやそれでもまだ抽象的すぎるので、「自己」概念についての経験的な知見に言及する方向か。大きくは変わらないがいろいろこなれた流れを目指したい。
来月の駒場オムニバス講義では、先日書いた日本語総説のストーリーに沿って、これまで作ったスライドを再構成しようと思う。NCCという研究プロジェクトがあって、contrastive methodで強い表象主義なのだけれども、内的な意味を持ったカテゴリーを作るためには行動をして、その主観にとっての意味を作らなければならなくて、ゆえに知覚と行動ととのループがどう作られるかを力学系的に捉える必要がある。いったんそのような意味での表象ができてしまえば、その脳の活動はふたたび同じ知覚刺激が与えられたときだけでなく、夢や想像でその神経ネットワークが活動した時にも知覚経験に対応した夢、想像の経験を作ることができるだろう。こういった考え方でまとめようとすると、Varela-Noeというよりも、Andy Clarkの穏当な表象主義あたりに落ち着くだろう。
じゃあってんで、「現れる存在」"Visual Experience and Motor Action: Are the Bonds Too Tight? (2001)"あたりでまとめるということになると、まあ池上さんはみんな知ってることだろうから面白みはないかも。
あと神経科学側から足せることは、ここ数年でのMark Churchlandの運動皮質ニューロンの仕事やMante et alのPFCの仕事のように、多数のニューロンの活動から状態空間を作ってその中で軌道を描く、みたいな仕事が実験側からも出てきたことを紹介するとかか。
つまり、革命家ではなくて改革主義者でいくと、(単純な)NCCを拡張して、行動からの意味付けによって形成したものにして(たぶんここにベイズ脳が入る)、(単純な)contrastive methodを拡張して、isomorphismからhomeomorphismになる。
"neural correlates"の歴史を辿るひとつのやり方として、「運動野ニューロンが何を表象しているか?」を追うと、個々の筋肉の活動に対応しているという説と、ポピュレーションコーディングをしているという説があったのだが、Steve Scottによってそれらがoptimal controlをしている内部モデルの一部として捉えればよいということで統一された。これは知覚での予想コーディングへの流れと似ているというか先取りしている。(というか川人先生の説ってのは知覚も運動も両方それで説明する。)
駒場講義でIITについてちょっと喋れないかなあと思って大泉さんのPLoS Comb Biol論文 "From the Phenomenology to the Mechanisms of Consciousness: Integrated Information Theory 3.0"をぴらぴらとめくってた。IITがver.3になってて、現在と過去だけでなく未来への影響まで考慮するようになった。
あと、大泉さんから教えてもらったブログ記事も読んでる。これはあまり意味のないネットワークでもphiが大きくなってしまうという問題を指摘している。これ自体は前から議論になっていることで、indexの作り方しだいで対処できそう。
あともうひとつこのブログ記事では、"Pretty-Hard Problem of Consciousness"という言い方をしていて、これはIITの目指していることを端的に示していてわかりやすい。つまり、あるシステムがconsciousであるかをシステムのある時点(+直前直後)の状態だけから決めるという試みだというわけだ。行動とか来歴とか力学系的な軌道とかそういうものをprojectして、ある時点でのスナップショット的に捉えるという点ではNCCと発想は近い。だからChristof Kochがこれに肩入れするのかなとか思った。
京都のシンポジウムの時にも話したことだけど、IITに行動と環境との相互作用は明示的には入ってないんだけれども、高いphiを持つネットワークを作るためには進化と発達とをやり直すような経験が必要になるであろうという意味では全く入っていないわけではないはず。
以前の論文でGAで進化させてfittnessを上げるとIITも上がったというシミュレーションのPLoS Comb Biol論文 "Integrated Information Increases with Fitness in the Evolution of Animats"があった。たぶんphiを挙げてゆくためには、そんなかんじでagentを鍛えてゆく必要があって、ある時点でそのまま与えて作れるようではないはず。というか作れたらたぶん間違ってる。
"Pretty-Hard Problem of Consciousness"に関してはコメント#39がNed Blockの“Harder problem of consciousness”と同様であるとコメントしている。
あと、Chalmersがコメントしている。詳しいことはGiulio Tononiの反論を待ってからとのことだが、Pretty-hard Problem (PHP)をPHP1,2,3,4に分類している。つまり、consciousであるかどうかに関する我々の直感(石には意識はないだろう)に合致しているかどうか(PHP1)とあるシステムに意識があるかどうかを教えることができる(PHP2)。この違いは機能的なものからだけでは意識があるかどうか決められないから直感を使うよって話で、それではまたまたハードプロブレムに逆戻りなんだと思うんだけど。
ともあれ私の理解ではPHP2は、vegitative stateの人のように、直感的には意識があるとは思えないが、呼びかけに対してfMRIで反応が見られる例(「テニスをしているところを思い浮かべてください」)のことを想定しているのかと思った。
大泉さんから教わったけど、以前のIIT批判のブログにGiulio Tononiが反論(後述ブログ記事内のリンクのwordドキュメント)を書いてて、さらにそれへの応答が書かれてる。Giulio Tononi and Me: A Phi-nal Exchange コメントではChristof Kochも応答している。
駒場講義の方は昨年のそのまんまなら今すぐにでも出来るがそういうわけにもいかない。合評会、基礎論学会を取り入れて再構成、それなら今日中に作れる。それをバックアップとしておいて、日本語総説で作った流れで再構成する。これを火曜の夜まで時間が許すかぎりで、レジメをアップする。
意識を科学的に研究するためには一般的定義で充分 -> NCCとはなにか -> Logothetis and Schall 1990 -> Kreiman 2001 -> Wilke PNAS -> セマンティックプライミング -> 盲視 -> 可塑性(Sugita + Sur)-> 環境との相互作用 -> active inference -> 理論の必要性 IIT みたいな流れ。たぶんぜんぶは無理。
途中に「awarenessとattentionを区別する」、「awarenessとdecision biasを区別する」-> メタ認知、という流れもあったが、これは無理か。冬講義のときにこのへんやってみたけどもう少し工夫してうまくやる必要がある。
うーむ「力学系」についてしゃべろうとしたらけっきょく多賀さんの本とかから抜書きすることになって、これは私である必要はないなあという気がしてきた。だったら「精神・神経疾患におけるさまざまの意識経験」みたいな方向について集めてきた事例について話して当事者研究につなげるほうがよいか。
ただし、それならそれで「当事者研究について話してください」ではなくて「意識研究の中で当事者研究がどうしても必要になる」というふうに理屈を持ってゆく必要がある。うーむ。
そういうわけで、これまでにツイートしたのとかまとめて、半側空間無視や統合失調症の前駆期の意識経験などについてスライドを作ってる。
「日常的になにかを探しているとき、見つけにくいなあということが多くなることにまず気づきます。周りにいる人に尋ねると、ほらそこにあるよ…と簡単に言われて見渡すけれどもぜんぜん見えたものじゃないので、教えてくれた人がいらだって、ほらここ、と手にとって目の前に突きつけられるまで、ああ、あったとは思えないほど、ものが魔法のように消えてしまいます。」「目玉がどんなに一生懸命あたりを探っても、目的とするものの上に画像を結ばないのです」「高次脳機能障害者の世界―私の思うリハビリや暮らしのこと」山田 規畝子
べつの部分については以前抜き書きしてブログにまとめたことがある:研究関連メモ20140316
あと、"Psychedelic" Experiences in Acute Psychosesに関してもブログに抜き書きを作った:Aberrant salience仮説と潜在制止と主観的経験
超訳気味に日本語にしてみよう:「「いちばん最初に起きたのは、私の脳の眠っていた部分の一部が目覚めて、さまざなま人、出来事、場所、考えに対して興味が惹かれるようになったことです。それらは普段だったらなんの印象も憶えないようなものでした。」
「全てのことになにか圧倒的なまでに意味深いものがあるように思えるのです…知らない人が道を歩いているのを見ると、そこに私が解釈しなければならないなにか徴(sign)があるように思えました」
「私が入院をした頃には、窓枠の光や空の青さがあまりに重要な意味を持ちすぎて叫びたくなるような、そんな「覚醒状態」の段階にまで到達していました」」
それから中安信夫 (1999) 初期分裂病 日医雑誌より、
- 自生思考 「自分で意識して考えていることと無関係な考えが,急に発作的にどんどん押し寄せてくる」
- 気付き亢進 「他人の声や不意の音,たとえば戸を開閉する音や近くを走る電車の音などを聞くとビクッとして落ち着かなくなる」
- 緊迫困惑気分 「何かが差し迫っているようで緊張を要するものの,なぜそんな気持ちになるのか分からなくて戸惑っている」
- 即時的認知の障害 - 即時理解の障害 「他人の話の内容,テレビの内容などが理解しにくくて,なかなか頭に入らない」
前駆期という概念自体には問題があって、これらの症状は最終的に統合失調症を発症する人でなくても一時的には起こりうることなので、前駆期に投薬などの処置をすることは多量のfalse alarmの例を含んでしまうことだろう。ただし、発症過程のメカニズム理解の意味では重要な資料だと思う。
当事者研究が自己と経験の構造について重要な知見を持っていること、でも人それぞれでのその経験をどうやって共有するかという問題までたどり着くと、じつは以前から興味を持っていたテンプル・グランディンの自身の自閉症としての経験と動物の行動理解の話というのがまさにこの間主観性の問題であり、そして私の仕事そのものの、動物での経験とヒトでの経験とをどう繋いで考えるか、という問題にぶち当たる。
これこそがわたしのライフワークと言ってよいものになるのではないだろうか。まだ骨格しかないのだけれども、かなり核心っぽいものに手を伸ばしている感触がある。
この歳でそんなこと言ってていいのかというのはあるが、あくまでこれはモノの言い方であって、これまで問うてきたことを拡張していった先の方向性が見えたというか、そういう意味で。
駒場講義の構成、いろいろやっていたらけっきょく統合失調症のサリエンス仮説を入れるスペースがない。11月の当事者研究の話を入れて話の流れを作るとそれだけで昨年とはずいぶん違った感じにはなっているのだが。Goodale縮めて半側空間無視を入れて3*2の腹側・背側経路を入れるか。
新幹線の中でだいたい講義スライドはできあがった。でも、広域科学の授業で「前反省的自己意識」とは「当事者研究」とか言い出すのは無茶な気もしてきた。ギリギリまで手を加えて、前提を吹っ飛ばさないようにする予定。両眼視野闘争に関しても今回はその手前のおばあさん細胞あたりから話をすることでもうちょっとはマシになったはず。そういう作業を繰り返して、極力置いてけぼりにしないようにする所存。でも眠い。
明日のスライド用に「せっかくだから、俺はこの赤の扉(神経現象学)を選ぶぜ!」っていうネタを考えついたが、元ネタ(デスクリムゾン)を知らないのに引用するのは正しくないなと思ったのでボツにした。
両眼視野闘争の説明の図で、ノートを丸めて望遠鏡みたいに覗きながら手の平を横に添えると、手の平に穴が空いて見えるという図がOlivia Caterだったかだれかが作っていたはずなのだが、探してみたら見つからない。こういうのは気がついたときにストックしておかないとダメだな。
該当するブログの記事をひとつ見つけた。
ブログ更新しました。「駒場講義2014「意識の神経科学を目指して」配付資料」
寝る!
目覚めた。あと4時間か…
Jakob Hohwyの両眼視野闘争の話をスライドに足した。part 1のスライドは97枚、 part 2のスライドは89枚。実際には"Any questions?"だけのスライドとかもあるから実質はもっと少ない。あとは丁寧に説明していければよいのだけれど。
corollary dischargeとmicrosaccadeについては削った。これで寄り道が減って、かなり一本道になってきたと思う。
駒場講義終了! 諸々重要な行事も終了で、新幹線で岡崎まで移動中。東京駅へ行くためにひさびさに満員電車に乗って、やっぱあんなの正気の沙汰ではないなあと思った。
講義の方は、前半の「両眼視野闘争、二つの視覚経路、盲視」は質問もたくさん出て、興味持ってもらえたようでよかった。以前に藤井さんがブチ切れたようなので、講義参加者の皆さんがへんに空気を読んでたくさん質問したとかでなければよいのだけれど(<-気にしすぎ)。
後半の「ベイジアンサプライズ -> 予想コード -> Active inference -> sensorimotor contingency -> ヘテロ現象学 -> 神経現象学 -> 当事者研究」の方は、ヘテロ現象学のあたりで参加者の集中力が落ちてしまったのがよく分かった。
もっと話の流れをこなれるようにすべきあったかもしれないし、「ヘテロ現象学(=現状の意識研究)を超えるものを探そう」というかなり無茶な問題意識を共有してもらうことに失敗したというのが最大の問題だったか。(まさに「せっかくだから俺はこの赤い扉を選ぶぜ!」としか言いようがなかった。)
てんかん患者さんの研究とかも入れてそれなりに具体的にはしたのだけれども。もし次の機会があるならば、元々考えていた、力学系と状態空間(Mark Churchland)とSOC (avalanchesとup-dawon state)とIITというテーマについて掘り下げるという方向でいこうかと考えている。
そうそう、IITが高い状態というのはSOCになっているだろうか、という興味がある。excitation-inhibitionのバランスでcritical stateになっていること自体は意識の充分条件ではないだろうけど、SOCになってないような回路はpathlogicalであるか、もともと考慮に入れる必要のないtrivialな回路(デジタルカメラの例のような)を排除できると思う。では、criticalであることじたいは内側から見て分かる特性か?って問題にもなるけど。
つかそういうことをこそ池上さんと議論できたらいいなあと。池上さんのMDFというのがIITやSOCから見てどういう性質を持っているのかとか。数理的なことが私の手に負えることとは思わないので、どうやれば寄与できるかとか考える。
大学院講義でcovert attentionを説明するときには、このレビュー論文の図3bを見てもらってから、「皆さんも夜コンビニに行くときに入り口でたむろしてるヤンキーと眼を会わさないようにしてるでしょ? でも注意はそちらに向きまくってる。あれがcovert attention。」と説明するようにしている。
今にして思えば、Covert attentionの話をするためには、その前に「そもそも我々の視野で視力が高いところはせいぜい1degくらいしかない」という話(腕を伸ばした先の親指の爪の幅が1deg)って話をしてからにすればよいのだな。だからこそfoveateする必要があるのだし、orientしたりfoveateしたりせずに、attentionだけを向けるという高度な技がヒトやサルでは可能となる。このあたりをちゃんとまとめると、上丘がそれぞれの動物でなにに関わっているかという比較認知的な話が出来そう。ウサギやカエルの上丘、視蓋には方向選択性がある話とか。
ゴプニックの赤ちゃんはランタン型の注意って話も、赤ちゃんは中心視野の視力が低いということと対応付けて考える必要がありそう。
2014年11月07日
■ PsychoPyで試行錯誤中(2014年4月版)
Ubuntu 14.04が利用可能になったのでVMWareに仮想環境作って、PsychoPyをインストール。SciPyとか一色全部apt-get installで入れて、python-setuptoolsを入れる。これでeasy_install psychopyできる。
これでGUIなしでimport psychopyできるようになる。GUIは立ち上がらないので、python-wxgtk2.8 python-wxtools wx2.8-i18nをapt-get installすると/usr/local/bin/psychopyApp.pyが動く。
でもこのままだと画像(gabor.py)が出せないのでeasy_install pygletすると1.14が入る。でもこのままだと動画(MovieStim.py)が動かないのでavbinを入れる。エラーは出なくなったが動画は出ない。(<-イマココ)
(apt-get install psychopyすればいいだけなのだけど、一つ一つ確認しながらインストールしてみたかったという意味)
仮想環境だとディスプレードライバ周りで不都合あるし、かといってデュアルブート環境作るほど気合も入ってない。どうしたものかと思っていたが、USBメモリに環境作って起動させるというのが一番正しいのではないかと思い至った。以前はクソ遅かったが、USB3対応のものを使えば実用になるはず。
.@kosukesa 十河さんの例題18-1を見て、easy_installで構築してみようと思った次第です。
ただし、USB起動するときにはブートローダの書き込み先を間違えてOSに指定すると大変なことが起こるので気をつけないといけない。仮想OS上のGpartedからでやってみようと思うのだけど、トリッキーすぎるだろうか?
ここまでやっておいて、tobii analytic SDKがLinuxに対応していないことを思い出す。けっきょくWindowsでNBS presentationというのが一番はやいのか?
ついでにトビーのサイトを見てみたら、Tobii EyeX Dev Kit 139$というのを見つけた。これはeye tribeへの対抗か。こちらもWindowsだけ。はやくeye tribeがMac対応してくれるといいんだけど。
2/7の段階では"there are no plans for multi-platform support for the EyeX Controller and Engine yet"と言ってる。
2014年11月02日
■ わたモテとSAD
わたモテの黒木智子がSAD(social anxiety disorder)と捉えられるはずなのに日本ではそうならない状況について このコミック、チラ見した限り辛くて読めなかったのだけど、今度読んでみようと思う。
"The Most Mean-Spirited Anime I Have Ever Watched"
わたモテ1巻読んでるけど、いじめではなく「空気系ぼっち」だったので、その意味では予想していたほど陰惨なトーン(が裏を流れているというかんじ)ではなかった。
「動物に「うつ」はあるのか」を読んでいたらdisease mongeringの一例としてSADが挙げられていて、やっぱそうかと思った。正確な表現はp.98:「たしかに、それまではたんに「人前で緊張する」ととらえられていた人たちに対し、「社交不安障害」という病気であるとして、抗うつ薬による治療を過度にすすめる動きなどは、「病気喧伝」(…)といわれてもしかたがない面もあると思います。」
ここの切り分けは難しいけれども「人前で恥をかくのが恥ずかしい」という当たり前の感覚から、「身体症状が出て本人が苦しんでいて人前に出られなくなる」までの幅がある。"Disease mongering and drug marketing"には"One such example is social anxiety disorder, better known as shyness … "とか"shyness is a new disease invented by Glaxo"とかそれはそれで極端な表現がある。
読むべきはこれか:PLoS Medicine Disease Mongering Collection
SAD(社会不安障害・社交不安障害)とは? この記事は「SAD」と「恥ずかしがりの性格」との違いを明確に分けているので納得がいく。
わたモテは5巻まで読んだ。やっぱ自虐辛くて笑えない。救いはないが、聖者が理不尽にいじめられているという描写でもない。まさに空気系ぼっち。4,5巻辺りでちょっとトーンが変わったか? 出てくる教師がどれもクズで、作者の恨みの深さを強く感じた。
2014年10月24日
■ JackIn面白いな!
JackIn面白いな! SLAMもOBEもorienting responseも入っているので、なんかものすごく関連高いし。なんか出来ることはないかなと思う。
JackInにおけるBodyとGhostの関係は、[サッカードをすることで視線を移しながら獲得している複数の狭い映像]から[あるひとつのシーンを再構成して自分は経験している]という関係と対応している。もしくは感覚と知覚の関係、もしくは現象学で言う現出と意識、と対応している。
工学的にはSLAMを使って画像を縫い合わせて、頭の動きに依存しないひとつの大きなシーンを作ることはできるだろうけど、脳はそのようなひとつのシーンを表象してはいるのではなくて、探索で形成した空間マップと今見ている視野との関係として持っているのだろう。
ここまで書いてみて気づいたけど、頭と体を動かして空間マップを作るという概念はplace cellとかあのへんで進んでいると思うけれども、目を動かしてシーンをマップするという方についてはそんなにないように思う。そしてどちらにしても、どうやってその中からinvariantな構造を取り出すのかというのが共通の問題になっている。
さらにいえば自分の体という制限を越えた自分の体という制限を越えた不変項の抽出という意味では、物体認識についても同様だ。空間、シーン、形、くらいまではつながった。色だってそうだろう。そういう全体像でいったい自分には何ができるだろう?
ふたたび話をJackInに戻すならば、これはSLAMと画像のスティッチがなければ、BODYに付けたたくさんのカメラからの映像を流しっぱなしにして、それをGHOSTが見て操作するみたいな人力方式(株とかやってるヒトのディスプレーみたいに)になるわけで、骨格はそれと同じなんだけど、GHOSTが人力で頭の中にひとつの空間マップを作るのではなくって、SLAMと画像縫い合わせで明示的に作ってあることによって、それは一人のGHOSTだけに専有されずに複数の人間が共有できることになる。うーむ、インターサブジェクティビティー!
2014年10月14日
■ 研究関連メモ20140316
高次脳機能障害者の世界~私の思うリハビリや暮らしのこと 山田 規畝子を読んでた。「壊れた脳 生存する知」は体験談的な記述だったけど、こちらの本では半側空間無視における経験についての記述(p.85-93)がわかりやすい。
(左は存在しないという自覚を持っているというよりは)「これまで私にとって左側が存在しなかったがゆえにやってしまった失敗とか…を通して、実はそうしたことの「理由」として、間接的に、これは私にはそもそも左側が存在しないがゆえの失敗なのだ、指摘なのだという結論を出している」
「もし左側があるのならこんな失敗や不都合はあるはずがないという思考方法で、それはある程度以上、意識して考えようとすればそういうことかなと考えられるということで、肩の力を抜いてしまうとすぐに消えてしまう」
「たとえば左の壁際を歩いてみなさいと言われると、左のことがわからない人にとっては、わかっていない部分で何かしてくれと言われるのに等しく、それがものすごく怖くなるのです。…そこへ寄り過ぎると壁にぶつかるという予想もできる…何もわかっていないから右へ寄っていくのではないのです。」
もちろん、人によって経験はそれぞれ違っているだろうし、山田 規畝子氏の場合には脳内出血を三度経験していて、記憶障害や失認もあるとのことだ。
「それでも脳は学習する~高次脳機能障害者の世界を語る~」 これも参考になった。
以前失語症についての一般書を読んでいて、失語症について「いきなり外国に行った状態」と例えていて(しかも英語ではなくドイツ語にしてあるところが絶妙だった)、音は聞こえているが意味は為さない、反応が遅れるから会話に加わることが億劫になる、など自分の意識経験と地続きで深く納得した。
ということとまさに同じような理由によって、自分がLAに行ったときには日本にいるときよりも「コミュ障」になってしまうという経験について書こうとしたのだけれども、「コミュ障」という言葉が政治的に正しくないので使わなかった。
"Learning to Read Improves the Structure of the Arcuate Fasciculus"つか、FAとの相関からAFの構造が「改善した」って言うのはどうよ?太けりゃいいってもんじゃないだろう?
狭いところに入り込むネコの話 と、ミニチュアの滑り台を滑ろうとする2歳児の話と視覚の腹側経路、背側経路くらいでなんかの三題噺を考えられないだろうか。
Front. Hum. Neurosci. 2014 Perceptual decision making: drift-diffusion model is equivalent to a Bayesian model
Front. Comput. Neurosci. 2012 "Perception and self-organized instability"自由エネルギー、カオス遍歴、自己組織化臨界現象と重要そうなの全部入ってるので読みたい。
脳の活動パターンがself-organized criticalityにあることを考えるならば、脳の活動パターンの場合の数は組み合わせ爆発などはしなくてずっと小さいはずなので、そういう範囲の中で探索したほうがいい。IITはそんな、元からあり得ないような状態を排除することにじつのところ機能しているのであって、拘束条件が緩すぎるのではないかと想像する。
つまり、N個のニューロンがあって、発火するかしないかの二つの状態だけがあったとして、ある時点での状態は2^Nあるということになっているけれども、それは過大評価であって、実際にはニューロン全体での発火頻度の周りでしか分布していないし、状態空間のなかで歴史に強く拘束されている。
つかそれって統計力学だよな。それはわかってる。
多次元レクチャーとかそのへんの準備をしていて、神経解剖学の基礎についてのよい図を探していたのだけれども、けっきょくのところ「カールソン神経科学テキスト 脳と行動」でほとんど用が足りることを知った。だいたい必要なところにカラーのわかりやすい図があって本当に素晴らしい。
たとえば基礎の基礎でdorsal-ventral, rostral-caudalの位置関係について、ヒトだと神経管が脊髄から脳に入ったところで曲がっているため直感に反した表現になるのだけれども、この教科書ではワニとヒトとを並べて両方表示している。なんかファニーでよい。
Heschl's gyriを表示している図も探していたのだが、外側溝を開かないと見えないので、脳解剖の実際の図とかになってしまってかえってわかりにくい。これもこの教科書だとoperculumを開いてinsulaとHeschl's gyriが見えるような図がある。
Rolandic operculumって言葉は知らなかった。「弁蓋」とかいうくらいなので、とにかくペロっとめくったらその下にべつの脳表が見えるような構造が弁蓋で、マカクのV1の表面と折りたたまれて盲点に対応する部分も弁蓋。
マカクのV1はややこしく折りたたまれているので弁蓋があるけれど、ヒトのV1はcalcarine sulcusに沿っているだけなので、たぶんヒトのV1周りにはopercular regionみたいな言い方するところはないんではないだろうか。要確認。
2014年06月06日
■ 「自閉症の現象学」
身体論研究会のページを見て「自閉症の現象学」を読むべきだなと思った。岡崎図書館には入ってなかったけど「治癒の現象学」のほうは入ってる。
エヌ氏の成長・円錐この記事を見ていても、社会性がどうのこうのという話よりも感覚・知覚の統合能力という議論になっていて、この方が自分的には納得がいくし、自分にもできることがあるように思う。そしてこれは意識の研究になると思う。
「治癒の現象学」借りてきた。「結語」の部分の「現象学者は…対象に巻き込まれる。彼は客観的な視点を取らない。それゆえに自然科学的なエビデンスは持たないが、巻き込みの中での追体験が別種の確かさを生み出す。これをフッサールは明証性と呼んだ」とかこのへんにピンときた。
「現象学者が行うのは、経験そのものの追体験ではなく、経験を「創りだす構造」の再作動である。」こことかもすごく合点がいった。いわゆる現象学的な心理学とか精神分析とかって精緻化された内観報告にしか思えなかったのだけど、もし構造を取り出すのならそれは現象学的だろう。
でもって本文を読み進めてみたけど、これはあまりに現代哲学的で読み進められなかった。「生身の身体のもつ運動感覚とはフッサールが考えていたような単一の純粋な現象ではなく空想身体と生理学的な身体との複合現象、交差する地点のことなのかもしれない」とかすごく興味あるけど進行が速すぎる。
「幸いなことに現象学は一種の数学を使わない「科学」である…たえず間違いは訂正され、知が伝承される可能性を持つ。たとえ新しい概念を創りだしたとしても、それは分析者の個性の発露ではなくて、新しく発見された現象や構造への目印である」とか惹かれるけど、事実というよりは目標では?と思う。
「行為の方は空想身体に埋め込まれる」「触発する出来事や人間関係は、それ自体は目に見えない。現実と近くの背後から空想身体を触発する。そしてその触発において空想身体は「意味」を産出する」こういう部分はすごく惹かれる。惹かれるんだけど、そういうところで断定的に進むのには着いてゆけない。
ここで書かれているような「科学とは違った意味での明証性evidenz」が本当に確立していて、現象学による経験の構造についての研究プログラムというものが概念の拡散をせずに意見の一致を見る形で深化していけるのだとしたら、現象学の自然化なんてものはそもそも必要ないのではないだろうか。
現代思想-2013年8月号-看護のチカラ-“未来-にかかわるケアのかたち これに惹かれたが、岡崎市立中央図書館では「現代思想」をとってない。ありえないことだが、とってない。
現象学的心理学、みたいな方向を調べていくと「質的研究法」についてちゃんと理解しなければならないことがわかる。うーむ、そこまではまだ(私の中で)時が熟していない感じ。
Dan Zahavi: "Empathy and mirroring: Husserl and Gallese"(pdf)ってのを見つけた。フッサール現象学でのempathyとか間主観性とミラーニューロンの話。
「自閉症の現象学」取り寄せてぴらぴらめくってる。とても面白いし、書いてあることがすごく納得がいくのだけど、話の展開の仕方が決めつけ的なのが非常に気になる。
「感覚刺激に没頭する自閉症児の世界は、このような感性野がひとりでに組織化する現象が、純粋な姿で実現している状態である」(p.11)ということでフッサールの「受動的綜合」のより純粋なものが自閉症児の世界にあるのだという話で、すごく魅力的なのだけど、こんな言い方でよいのだろうか?
「受動的綜合が見られるのではないだろうか? その論拠を示す。」みたいな議論でないのがすごく気になる。読み続けていけば違うのかもしれないけど。
- / ツイートする
- / 投稿日: 2014年06月06日
- / カテゴリー: [オートポイエーシス、神経現象学、エナクティヴィズム] [脳科学メモ]
- / Edit(管理者用)
2014年04月29日
■ "Messy Mind"の威力
アンディ・クラークの「現れる存在」の付録部分(駒場での連続講義)読んでたんだけど、"Messy Mind"という概念を知って衝撃を受けた。つまり、ニューラルネットワークを使って外から問題与えて学習させるようなパラダイムを、FPGAのような物理的な実現の制約の中で行うと、ちょっとありえないような雑な('messy')な解決方法が出てくると。ここで出てきたのは論理演算をするデジタルな回路を学習するはずが、隣接した回路からの影響(回路をデザインするときには極力避けたい部類)というアナログな影響を積極的に利用しているという例だった。アンディ・クラークによるスライド資料:Messy Minds: Embodiment, Action, and Explanation in 21st Century Cognitive Science
Messy MindとFristonがどうつながるのかがわからない。Messy Mindは問題解決をするのは必ずしも脳でなくても良いし、身体の機構を使っても良いし、あるものなんでも使うって話である一方で、フリストンだと予測誤差はなんか脳内でもいいし、行動でもいい。というあたりでつながるのだろうか。Messy Mindで「漏れだす」という言い方をする一方で、active inferenceではいちばん予測誤差を大きく減らせるところで減らしてゆく、みたいな最適化の思想かと思っていたのだが、messy mindもイメージはGAだろうからそんなに変わらないのか。
Messy mindの話を読んでいて、たしかにこのようにして漏れ出すものをいくらでも使っていけるのだとしたら、ニューロンのスパイクだけでなくて、局所電場電位のような、もともとはもしかしたら脳の層構造が生み出したbyproductに過ぎない(かもしれない)ものをなんだかんだと利用してシステムの一部に組み込まれていて、そこには整然としたハイアラーキーとかは必ずしもない、っていうような理解でこのへんはすごく納得がいく。
脳波のcross frequency couplingとかも意義が正直よくわからなかったけど、とにかく起こってしまったものが使われて、システムに組み込まれてしまうのだったらそれもありかなと。自分が思った以上にsingle-unit至上主義者であったことに気づいた。
こうなると、脳と身体と環境、そして脳の中でもミクロとマクロの階層性みたいなものをもっと断念する必要がある。もともとそれはrecursiveなものなのだからシリアルではないっていうことはわかっていたつもりだったし、神経回路が短絡路だらけであることもわかっているつもりだったけど、messyであることの威力はそれ以上だった。
もしmessyであるのならば、物理的な遠隔作用であれ、シンボリックなものであれ、成り立ちの必然性から読み取ろうとするのは不充分であり、無謀であるようにも思えてきた。もちろん反動はあって、じつのところどの程度までmessyであるか、決定的な影響はあるのか、という線形 vs. 非線形、みたいなことを検証してゆく必要があるのだろう。
messyといえば、盲視そのものが、同名半盲からの機能回復の方策としてはmessyな解決方法であるといえるのではないだろうか、ということに思い至った。
"messy mind"が「猥雑な」ってなんかピンと来ないよなあと思ってmessyでgoogle画像検索してみたら、汚部屋の画像がぞろぞろと出てきたので納得がいった。似非関西弁で言うならば「むちゃくちゃですがな」みたいなかんじ。
2014年04月19日
■ NINS Colloquiumで話題提供しました(2013年12月)
NINS Colloquiumが近づいてきたけどどうしたものか。「セッション4 時間の流れに沿ったエポックの発生と「揺らぎ」」というところで話題提供をすることになっている。
自分の立場からすると、意識が脳と身体と環境の相互作用によってさまざまな意識下の情報の流れを背景に、意識的なものが生まれては消えということを繰り返す。Transientなセルアセンブリが生まれては消える、みたいなVarela的な意識観を紹介するとして、そのまえに自己紹介的に盲視の話をして、意識的な視覚と「なにかあるかんじ」との対比の話をその話の導入に使う。
「意識と時間」みたいな話では面白いトピックはいろいろある。たとえばLibetのtime-on仮説もRPもそうだし、color-phiとかbackward maskingとかsaccadic suppressionとかpostdictionとかそういった、「脳はじつは切れ切れになった情報処理を、必ずしもシリアルではないやり方で並べて、ある主観的な時間の流れを作っている」みたいなかんじのお話をすることはできる。
でもそれだけよりかは、力学系的脳観と神経現象学と、意識の統合を目指すトノーニのIITのような試みとかを紹介する方向のほうが楽しいと思う。
30分で収めるとしたらこうか:1) 盲視が明らかにした「視覚意識」と「なにかある感じ」 2) 脳は後づけで意識の流れを作る(止まる秒針の話) 3) 不安定な構造を作っては壊すという脳観 4) 力学系的な脳と一人称的な経験の流れ(精神現象学) って無理か。
NINSコロキウムの内容をもうちょっと考えてみたけど、あんま意識がどうのって話をするよりは、脳の活動のゆらぎとかそういう実験事実を中心にしたほうがよさそうだ。たとえば、同じ分科会には物理経済学の高安秀樹氏がトークするから、意思決定のdiffusionモデル(=ランダムウォーク)とかなんかはかなり共通性を強調して議論できるだろう。
以前書いたことだけど、diffusionモデルは一定の閾値を想定しているけど、実際には砂山が崩れるように臨界状態でのバーストを考えたほうがいいのだが、まだそこまで進んだモデルが役に立つレベルであるわけではない。
同様に、サリエンシーモデルもちゃんとやれば視覚刺激を処理するフィルターから、spiking neuron networkのダイナミクスへと移行できるはずだ。やっぱ話の順番を変えて、脳と意識の概論から、盲視の研究を紹介して、より力学的な方向性のアプローチを提案する、くらいまでか。
うーむ、いっそのこと、「意識」も取っ払ってしまって、ChrchlandのNatureとかNat NeurosciとかNewsomeのNatureとかあのへんを持ってきて、意志決定、運動開始、知覚、さまざまなところでinstabilityが起こっている証拠が集まりつつある、みたいな話をもってくるほうが(VarelaとかKelsoとかのEEG関係のデータよりも)強くて良いのかも。学会じゃあないんだから堅さよりは、面白いと思ってもらえればよい。他人の仕事の紹介になっちゃうから、あんま広がりは期待できないのだけれども。
脳と力学系という意味ではEEGによるseizureの予測というのが事例としてよく使われるのだが、じつのところMARTINERIE et al (VarelaとLe Van Quyen)とかも含めて再現性に難があって問題は解決してない。Brain. 2007 "Seizure prediction: the long and winding road"
NINS colloquium の話題提供終了。ランダムウォークの話に絡めるためにサッカードの応答潜時のdiffusion modelを持っていったが、話が細かいところに入りすぎてしまってこれはうまくなかった。力学系の話まで出来たので目的は達した。ただ、物理学者の前で力学系の話をしても当たり前すぎてインパクトがなかったので、ここからどう議論を展開するかが課題。
ブレインストーミングも終了。物理学者はみんなズケズケ物を言うのでけっこうしんどかったが、最終的には頭の中が整理されて、すごく有益だった。最後にはフリストンの自由エネルギーの話までした。と入っても詳細を理解してもらったわけではないのだけれども、
宇宙や分子とか、本当に物理法則で力学系をやっている人たちと、経済とか脳とかアナロジーとしての力学系をやっている人がいて、経済物理学でやっているようなことは力学系モデルとして良いプロトタイプになるのではないかと。要は力学系であるからには拘束条件となる多様体があって、多様体とかカッコつけたけど、とにかく曲がった平面があって(脳が取りうる状態は限られている)、そのうえでゆらぎを持って動くときの物理法則、例えばポテンシャルに対応するものを見つけることができるならばそれを充分捉えられるだけのデータ量とミクロマクロの記述レベルが決まると。
そういう話を伺って、なるほどそれはまさに自由エネルギー最小仮説だなあと思った。私自身はたくさんニューロン記録して、状態空間作って、その推移則の微分方程式みたいなものが作れればいいんだろうと、間違ってはいないけどどうとりかかったらいいかわからないイメージを持っていたのだけれど、高安先生の話にあったような、ポテンシャル関数が推移則の中に入っていて比較的低自由度で動態を記述できるというやり方なら、脳全体からの同時記録とかができるようになる前でも、空間的に限りはあってもより長時間の記録で高次の空間までありうる状態を記録してゆくことでできることはありそうだ。
力学系、力学系とさんざん言ってきたが、実のところ脳に応用する限りそれは単なるモデルであって、本当の物理ではない。だから、うまくいけばよかったねだし、うまく行かなかったらモデルが悪いか評価関数が悪いかという問題。
もちろん、ニューロンの発火自体は物理であって、HHモデルは微分方程式なわけで、スパイキングネットワークモデルはHHを簡略化してはいるけれども物理だけでニューロンのネットワークの発火までやってしまおうということ。
行動分析や連合学習理論はそういう意味では行動の推移則を作るだけのデータとモデルを持っているので、いま書いているような考えとは実はかなり近い位置にあると思う。
昨日書いてたことでひとつ抜けていたのが、ブラウン運動とランジュバン方程式の話か。ってググってみたら酒井裕さんのテキストが出てきた。
Diffusion modelってもっと力学系にできないもんかと思って調べてみたら今回見つけた論文:Roxin A, Ledberg A (2008) PLoS Comput Biol Neurobiological Models of Two-Choice Decision Making Can Be Reduced to a One-Dimensional Nonlinear Diffusion Equation
2014年03月19日
■ ここ一年分のブログエントリを電子書籍化したもの作った
Kindle Paperwhiteを買っていろいろ入れて重宝してる。ふつうにamazonから電子書籍を購入するだけではなくって、Kindleにはパーソナル・ドキュメントという仕組みがあるので、電子書籍を自分で作ってメールとかUSBとかを介してKindle本体に送ってやることができる。
どうやって電子書籍を作るかというと、いちばん手っ取り早いのは、htmlファイルを用意して、それをKindleGenというコマンドラインで実行するプログラムを使って、Kindleで読めるmobi形式に変換してやる。(以下全部Mac上で作業してます。)
たとえばPLoS Biologyのサイトに行って論文のXMLをダウンロードする。たとえばPetrides and Pandyaだとjournal.pbio.1000170.xmlをダウンロードして、これをxhtmlにリネームして(xmlのままだとwarningが出て変換してくれない)、kindlegenを動かせば journal.pbio.1000170.mobi が出来上がる。
cp journal.pbio.1000170.xml journal.pbio.1000170.xhtml ./kindlegen journal.pbio.1000170.xhtml
xmlファイルの中身をまったくいじらずに変換すると、要旨無し、画像なし、本文だけのmobiファイルができる。要旨や図を入れたかったら、xmlファイルを編集してやればよい。一個なら手作業で、複数ならなんかスクリプト書けばよい。今回は省略。
mobiファイルをMacからプレビューして、正しく作成できているかを見るためにはKindleプレビューツールを使えばよい。機種ごとにどのように見えるかを確認できる。ただし、このアプリを使ってみたところ、動作がけっこう不安定なので注意。
ここまででしばらく満足してたんだけど、せっかくだからこのブログの一部をまとめて電子書籍化してみようと思い立った。といってもKDPで売るとかそういうのではない。
ともあれ必要なデータは以下のとおり:
- a.opf (電子書籍の定義ファイル。ここで本体のhtmlや目次、表紙のファイルを指定する)
- a.html (本文をhtmlで作成。スタイルシート(CSS)も利用可能。)
- toc.ncx (目次の定義ファイル。)
- a.jpg (表紙の画像ファイル。)
a.opfとtoc.ncxの形式については、最後に置いた参考サイトの情報を見てほしい。基本xml形式なので、必要な情報だけ入れ替えて使えばよい。
それで今度はopfファイルをkindlegenで指定してやると a.mobi という電子書籍ができる。
./kindlegen a.opf

こんなかんじで私のブログの最近1年分くらいのエントリから研究っぽいものだけ編集したものを作ってみました。さらに元のhtmlファイルからPDFを作成したものも置いておきます。ご自由にどうぞ。
- よりぬきプーニールさん mobi版 790KB
- よりぬきプーニールさん pdf版 9.5MB 107ページ
CCライセンスとかちゃんと考えておきたいのだけど、まだちゃんと理解してないので今回はなんも付けてない。
表紙のためのフォントを探していて、ちどりフォント(仮)ってのを見つけたんだけど、非売品だった。
今回のエントリの作成には、以下のサイトを参考にしました:
2014年03月11日
■ 「方位」と「方向」ってどう違うの?
JNS2013 Emergent Properties of the Optic Tectum Revealed by Population Analysis of Direction and Orientation Selectivity メダカの場合は網膜と上丘の両方で方向選択性が作り上げられる。それにしてもなぜ霊長類の上丘では方向選択性がないのだろう?
必要なphotoreceptorとかが足りないわけでもないので、二色性、三色性色覚の話とも違う。だから、大脳皮質で損傷が起こると、皮質下の方向選択性が再生したりとかしないだろうかと考えてる。でも直線的な進化を反映しているようにも思うし。まだわからん。
Lettvin and MaturanaによるFrogのbug detectorはきっとprimateにもあって、それがsaliency detectorとして働いているんだろうと思う。そういう「お話」をどう持っていったら意味のあるものにできるだろうか?
V1ニューロンの選択性とかで、orientationのことを「方位」って訳して、directionのことを「方向」って訳して別物として取り扱っているけど、元々の「方位」の語自体は必ずしも0-180degの範囲ではない(方位磁石とか)。どういう経緯でこういう訳になったんだろう?
こっちのフォーラムでのorientationとdirectionの違いを見るとさらに分かってきたようなわからないような。
たとえば"direct our gaze"という言い方はするがorientは使わない。gaze(見ている位置)は動く。一方でorienting of attentionというときは私を中心にしてそこから注意を向けている。orientって自分は動かずにrotateするかんじか。
方位=orientationには基準があって、方向=directionには基準がないという説明をいくつか見つけたけど、方位というのは(自分からの)向きといったエゴセントリックな含意があるということを反映してのことなのだろう。
追記:「方位」の話はけっきょく、orientationという言葉自体には(たとえば)90度と270度が等価になるような意味はないのだから、正確には「傾きtilt」とでも言うべきなのではないだろうか?こっちなら、水平(0度)から右縦(-90度)までと左縦(90度)までで定義できるし。
2013年12月20日
■ Eyelink 1000セットアップ中。
Eyelink 1000セットアップ中。課題はMacでPsychoPyで動かすということで、そのまえにSR Researchが提供しているappで稼働させて、つぎにPTB3で稼働させて、PsychoPyにやっとこさ辿りついた。
PsychoPyでは、SR researchが提供しているpylinkを使う。デモプログラム(gaze contingent display)があって、カクカクするがとにかく動いた。さてこれをこれまで使ってたプログラムとmergeするというのが次の課題。
そこでpygameまわりでハマった。いろいろやっていたがけっきょく、pygameはなんとデュアルモニタに対応してないということが判明。pygameを一旦開いて、閉じて、それからwin = visual.window()をするというきったない方法で対処。
そしたらPsychoPyは1.77からioHubというプラグインで外部入力を統一的に扱うという形式に移行していることを知る。EyelinkでもTobiiでもSMIでもぜんぶ同じスクリプトで、設定ファイルはアイトラッカーごとにyamlに別々に書かれている。これは魅力的。
だがかといってここでこれまでの環境(1.73)をアップデートして、せっかくこれまで稼働していたのをぶっ壊してしまうわけにもいかない。べつべつにインストールすることはできるけど、もともとそういうことを想定しているわけではないから、なんか予想外の挙動とか示されると困るのでやらない。
というわけで、mbp2011のほうはアップデートせずに、mba2013のほうで新しいバージョンを試してみることにする。Tobii X2-60がもうすぐ届くので、こっちと両方一挙にセットアップ出来れば手っ取り早くてよいのだけれど。
さてさてあとはディプレイに貼りつけたフォトダイオードの信号をどうやって取り込むかだけど、前回TX300を使ったときは急ごしらえだったのでwindowsでNI USB-6008への入力を取り込むC#プログラムを改変してなんとかしてた。
でもいま冷静になって見回してみれば、PyLibNIDAQmxなんてものがあることに気づいた。たぶんこれでMacでPsychoPy上で全部いけるんではないだろうか。PsychoPyからだったらLabjack U3という話もあるが。
ってよくよく見てみたら、開発は2009年で止まってるし、NI DAQmx baseには対応してないくさい。ダメじゃん。こっちが正解か:pydaq 日本語のブログ記事:NI 6008をpythonで
って話の発端は、十河さん@SogoHiroyuki の「PsychoPy/VisionEggからTobiiを使うTobiiController」を読んでいて、そこからいろいろ調べていたのであった。
後日談。結局Labjack U3を買ってテストしてみたら、win.flip()の時間とLabjack U3で測ったディスプレーの描画時間はきっかし33ms (60Hzで2frame分)で、繰り返しによるばらつきは<1msだったので、ちゃんと垂直同期とって誤差なく描画していることがわかった。そういうわけで、毎回フォトダイオード計測しなくても、この33msだけわかっていれば充分だということが判明。
あと、PsychoPyとEyeLinkの間の通信の遅延に関しても、tracker.sendMessage(msg)してやればイーサネットで1ms程度の遅延でタイムスタンプをedfファイルに送ることができる。こちらもこれで充分。ということでスレッドで平行してアイトラッカーとフォトダイオード計測するとかそういうややこしいことをする必要はなくなった。
2013年08月23日
■ タブレットをケージに貼り付ける作戦、について試行錯誤
タブレットをケージに貼り付ける作戦だけど、AndroidでProcessingでNexus 10あたりかと考えていたが、Windows 8タブレットがだんだん良くなってるみたいなので、NBSのPresentationを入れて動かすという以前の策を再考するタイミングかも。
いまのところ本命はASUS TransBook TX300CAで、Core i5 1.7GHzで13.3インチ。問題はメモリーが4GBしかないことで、Windows 8だと動作がもっさりしてるかもしれない。
タッチパネルが大きければ大きいほどよいのだけどとか思っていたら、ONKYO TW21A-B36C7 21.5インチっての見つけた。テーブルトップか。Core i7 メモリ8GBだし、ありかも。つかケージにどうやって入れるんだ。
ここまで考えた時点で、NBSのPresentationは更新してなくて使えなくなっていることを思い出した。毎年7万円払うくらいだったらProcessingでシステム構築する方が長い眼で見たら生産性高いかも。
Androidタブレットだとサイズが小さくなるのが難点。ならば、Raspberry Pi上にubuntu入れて、PsychoPy動かして、タッチスクリーン制御するというラインでどうだろうか?
つか、Raspberry Piほど小さくなくてもいいんだよな。
ググってみたら、Raspberry PiではOpenGL使えないからPsychoPyは使えないとのこと。残念。 Cognitive Sciences Stack Exchange
ただし、OpenSesameというのがあって、これのlegacy modeだとopenGL使わなくてもlinuxで心理実験動かすことが出来るとのこと。グラボ使わないということだから処理は遅いのでガボールパッチ動かすとかは難しい。
OpenSesameというのは知らなかったが調べてみたら、なんかすごくよさそうだ。しかも、Android版も出てる!Google Play 作者のSebastiaan MathôtはJan Theeuwesのところの人か。
これでProcessingでandroidという方向には行かなくてよさそうだ。Processingは心理実験のためのものではないからいろいろ整備しなければならなかったので、それなりに時間がかかりそうだったので。
Running psychological experiments on a Raspberry Pi with OpenSesame OpenSesameの作者がRaspberry Pi使って時間の精度を調べた。これを見たところだと、私の目的には充分だ。
OpenSesameじたいはPsychoPyでいうところのbuilderみたいな存在で、簡単な課題を作るの自体はExpyrimentで充分なのかね。さらに調べてみる。
TransAiO P1801 18.4型のAndroidタブレット/windows 8両方で使える。これにNBS presentation入れてケージに貼り付けて行動データ採集。金はかかるが人間の労力はかなり減らせる。
タッチモニタ + raspberry piでraspbian (Debian Linuxのヴァリアント)上でpsychoPyかopensesameというのがたぶんいちばん金額的には安い。人間の労力入れると分からんが。
京大野生動物研究センターの田中正之さんから「生まれ変わる動物園: その新しい役割と楽しみ方」をいただいたので読んでる。途中で、使ってるタッチモニタの品番まで書いてあったのでググってみた。
Raspberry Pi立ち上がるようになった。sshdをenableにして、sshで入れるようにした。さらにtightvncserverをインストールして、「画面共有」でMacから入れるようになったところ。

ひきつづき、opensesameを入れようとしているのだけど、NeuroDebianが対応していないようで、sudo apt-get install opensesameしてもcandidateがないと言われる。本家では動作確認あり。
2013年08月07日
■ 統計的因果推論
統計的因果推論ノート:「正しいセカイの切り取り方」 これ楽しみ。「ラットの因果推論」読んだときにこのへんは参照した:相関と因果について考える:統計的因果推論、その(不)可能性の中心および確率と因果を革命的に架橋する:Judea Pearlのdo演算子
その文脈でさっきのfMRI is not an inherently correlational methodを見るならば、ここで書かれている「二つの観察変数の間の相関をみるのは相関的方法であり、片方の変数を操作して(独立変数)、もう片方の変数を観察(従属変数)することによって見つかった相関は実験操作が因果的に引き起こしたものであると言える」という言明は正しい。
でも統計的因果推論の立場からは、実際には[グループA: 操作あり(実データ)-操作なし(反実仮想)] [グループB: 操作あり(反実仮想)-操作なし(実データ)]という欠測値のある2*2を見ているということがさらに付け加わるらしい。前者はp(y|x=30)であり、後者はp(y|do(x=30))となり、実験計画法的にconfouding factorがないようにしてcommon causeとかをなくすことで、「バックドア基準」を満たすようにする。
とかそういうことを前回調べたときに理解したのだけれども、正直まだよく分からなかったのが、いつでも変数をsetできるわけではないんじゃなかろうかということ。
因果推論の論文を読んだときに「注射をしてXの血中濃度をyy mg/mlにする」なんて書いてあったけど、血中濃度なんてホメオスタティックに変わってしまうし、これはあんまいい例ではないのではないかと考えた。
話を戻すと、統計的因果推論の立場から考えるならば、視覚刺激A提示と視覚刺激提示無しという実験操作を加えてfMRIで活動を見ることと、TMS刺激ありとTMS刺激無しという実験操作を加えてfMRIで活動を見ることとは、因果推論のレベルから見ると形式的にまったく同じであると言える。
だから、TMS(nhpならelectrical microstimulation)が「因果的である」と言う場合は、端的に「脳内への直接的な操作、介入」が「行動に影響を及ぼす」ということを意味している。
視覚刺激だって行動に影響を及ぼすのだけれども、そういうときの関係は「因果的」というよりは「contingencyを変える」というように捉えられるため、「因果的に行動を引き起こした」という言い方はされない。
でも実のところ、TMSだってelectrical microstimulationだって、力学的因果で行動を引き起こしているのではなくて、contingencyを操作しているに過ぎない。ただし、筋肉を刺激する > M1を刺激する > 連合野を刺激する > 視覚野を刺激する、と因果的関係の強さにグラディエントがあるために、これらの違いが曖昧になっている。
2013年08月04日
■ スマートピル関連
ADHD治療薬コンサータ錠18歳以上の成人期への適応追加承認申請 ん?以前リタリン乱用が問題になって、適応症が狭められて大人のADHDでは使えなくなってたんだけど、製剤を工夫して再申請したということか? (<-これでも薬剤師)
Natureだったかなんだったかのサーヴェイで、研究者とか大学院生の結構な割合がその種の中枢刺激薬使ってるみたいな話があったと思うんだけど、いざ探そうとすると見つからない。
"Studies have documented usage of drugs for cognitive enhancement by 5–15% of students"
20%って数字の根拠はこれ。 でもオンラインのサーヴェイだからいろいろ偏りがある。もっと最近のこちらで調査の呼びかけをしている。
Smart drugs: would you try them? 英guardian誌がなんかしれっとerowidにリンクしているから吹いた。あそこってもっとアングラなところかと思ってた。LSDの合成法とかあるし。
で、いろいろ調べてて、Martha Farah ("Visual agnosia"の人)がいまではneuroethicsの人となっていることを知った。
Martha Farahの関与についてまとめると、Nature. 2008 "Towards responsible use"のラストオーサーになっている。
そのほぼ同時期に「Adderallはじつのところ認知能力を上げるわけではないのだが、本人は認知能力が上がった気がしている」というpreliminaryな調査を発表している。
これはTime誌とかにも取り上げられて評判になったらしいが、それと同時にたぶん反論がたくさんついたのだろうと思うけど、「実際に認知能力を上げる」とする結果がたくさんあることがこの記事の"UPDATE"に追記されている。
それへの応答だろうか、Psychol Bull. 2011においてメタアナリシスを行い、「スマートピルがノーマルな人々に対して認知能力を上げることはたしかに認められるが限定的である」というようなことが要旨に書かれている。これにもふたつコメントがついているので、いろいろ反論はあったのだろう。
前述のPsychopharmacology 2009は予備的研究だったが、さいきんNeuropharmacology 2013というのが出て、やはり「Adderallは(ほとんどの指標で)認知能力は上げないが本人は上がった気がする」という結論を繰り返している。
ここまで書いておいてなんだが、じつのところ「認知能力」の向上にはあまり興味がない。「意識経験」の変容には興味があるが。
2013年07月04日
■ うーむなぜだか「意図」intentionの定義について調べたくなってきたぞ?
なぜだろうふしぎ。
神経生理学では「運動の意図」という文脈でintentionについて調べられてきた。たとえばRichard A. Andersenらはposterior parietal cortexのニューロンの性質として「運動の意図」を反映していると主張してきた。Annual Review of Neuroscience 2002ではintentionを定義して、「実際に運動を行う前に、どこに向けてどういう動きをするか計画する段階」のことを指している。これは実際の運動ではさらにどういう動きを、どの筋肉を使って動かすかといった過程が続くため、それらと区別した表現になっているというわけだ。
以前のブログで、Jeffrey D. Schallのレビューでchoice(行動選択)とdecision(意思決定)とintention(意図)の違いについて書いたものを採りあげたことがある。 Jeffrey D. Schall, Decision making, Current Biology, Volume 15, Issue 1, 11 January 2005, Pages R9-R11
「行動選択」とは、選択可能ないくつかの行動の中から一つを選び、実際に行動を起こすことを指す。一方で「意志決定」とは、選択可能ないくつかの行動を選ぶこと自体であり、行動に移る前に熟考する過程を指す。「意図」は、行動の準備そのものではない。ある行動をしたときに「なぜその行動をしたのかの理由」を答えるものでなければならない。Andersenらが見ているのは行動の準備でしかない。人間以外の動物が「どういう意図でその行動をしたのか」を報告できない限り、ヒト以外で意図性について議論するべきではない。
Haggard P, Clark S. Conscious Cogn. 2003では、「意図的な運動」と「非意図的な体の動き」を対比する際に、意識的な意図とは、「そのactionの原因はなにであるか」を体験することであるとしている。たとえばanarchic hand syndromeの症例では、患者は自分の腕のownership(自分の腕であること)は疑わないが、自分の意図が腕の動きの原因となっていることを否定する。
動物行動学(ethology)の文脈だと、行動の分類として「意図行動」(intention movements)というものがある。動物がある行動を行う際に、それに先だっていつも行われる運動が見られることがある。このような運動からその動物が何をしようとしているか(意図しているか)が分かるということから、このような運動のことをHeinroth 1910は「意図行動」と呼んでいる。たとえば、オオカミが「噛みつく」という行動をする前に「牙を剝いてみせる」という運動をする。これが意図行動。他のオオカミがこの意図行動を見て次に起こる行動を解釈する、これがコミュニケーションの分野でのシグナルの原型と考えることが出来る。(Perspectives on Animal Behavior by Judith Goodenough, Betty McGuire, Elizabeth Jakob, p.388)Google Books
辞書を調べてみた。Animal Behavior Desk Reference. A Dictionary of Animal Behavior, Ecology, and Evolution by Edward M . Barrows によるとGoogle Books、intentionalityとは日常的な意味としては「ある個体が主観的に抱いた目的に向かって行動するのを計画し、実際に取りかかること」となる。(哲学的な意味での「志向性」とは異なった意味を持つ。)
この項目自体の元ネタはImmelmann K, Beer C. 1989. A dictionary of ethology p.153で、該当部分をGoogle Booksで調べてみるともっと突っ込んだことが書いてあって、intentionalityの定義というのは「心理主義」的な含意が大きいので動物行動学者はintentionalityという言葉を使わなかった。しかし近年「認知動物行動学」が栄えることでこの語が使われるようになっている。たとえば、distraction displayのような行動では動物がdeliberatelyでだましの行動を行っている。このような行動を説明する際にはintentionalityという側面からの説明が有効となり得る。
intention / intentionalityという言葉にはanimal consciousnessに関わる問題がある。"Essential Animal Behavior"ではこういう記載がある。「intentionalという言葉を動物行動について使うのはいくぶんか問題がある。ここで動物には意識があるかどうかという問題に入り込むつもりはないし、動物がintentionを持つとして、それがわれわれ人間が持っていると思っているものと同じものと見なすつもりもない。"intent"という言葉には自発的な行動を指す「自発的意図」とはべつに「進化的意図」を考えることが出来る。後者では、個体がある行動を選択して実行するのではなくて、自然選択の結果としてその行動が行われる、というふうに捉えることが出来るので「非自発的な意図」と表現することが出来るかもしれない」Essential Animal Behavior by Graham Scott p.94 Google Books
というわけで、動物に自由意志とか主観的な経験とかを仮定しなくても、合目的な行動を行うための企画段階のことを指すものとして「意図」という言葉を使っても良いのではないかということらしい。
前述の通りintentionalityはコミュニケーションの場面で出てくる概念なのでtheory of mindにも関わってくる。Theory of mindを持つということは相手の意図を読めるかどうかなのだから。
Shared intentionalityという概念があって、Tomasello M, Carpenter M. Dev Sci. 2007 Shared intentionality.では以下のように定義されている。「(ある社会的な集まりの)参加者が心理的な状態を他の参加者と共有する際の協力的な相互作用」("Shared intentionality, sometimes called ‘we’ intentionality, refers to collaborative interactions in which participants share psychological states with one another (Gilbert, 1989; Searle, 1995; Tuomela, 1995).")
Tomaselloによれば、個体がお互いに他の個体の視線の先を追うこと(gaze following)じたいはshared intentionalityではない。そのときに「お互いに同じことをやっていると知っていること」があってはじめてjoint attentionが成立する。つまりshared intentionalityの有無がgaze followingとjoint attentionを区別する。
Intentional communicationのcriteriaとしてこの論文のはわかりやすかった。こちらはgaze followingのレベルと言ってよいと思う。 (a) そのシグナルの受け手がいる (b) 受け手の注意が影響を受ける (c) 受け手が送り手とそれが指す物体とのそれぞれにくりかえし眼や頭を向ける (d) 送り手は注意を惹きやすい行動を行う (e) もし注意を惹くことに失敗したときには、繰り返し、そしてもっと手の混んだ仕方で行う Leavens DA, Russell JL, Hopkins WD. Child Dev. 2005 Intentionality as measured in the persistence and elaboration of communication by chimpanzees (Pan troglodytes).
あとは、行動分析的側面からのintentionとintentionality。 Paul Neuman Behav Anal. 2004 Spring; 27 (1): 55–65. An intentional interpretive perspective Paul Neuman Behav Anal. 2007 Fall; 30(2): 211–216. [Some Comments on the Distinction Between Intention and Intentionality](http://www.ncbi.nlm.nih.gov/pmc/articles/PMC2203638/ これはむずかしい。わからん。John Searleが提案したprior intentionとintention in actionという概念を出して、前者がいわゆるintention (行動を引き起こす原因と見なされるような)であって、後者がintentionality (行動しているということを経験していること、Haggardとかの文脈だとawareness of intentionか)ということになるらしい。それにさらに行動分析的な解釈が加わるのだけど、このへんで諦めた。
そんなわけで、私が600字でまとめるとしたらこんなかんじだろうか。
意図(intention, intent)、意図性(intentionality)とは:
ある個体がなんらかの目的に向けた行動を計画すること。実際に行動を起こすために筋肉をどのように動かすか決めることなどとは区別される。「行動選択」では実際に行動が行われるが、「意図」では必ずしも実際に行動を遂行するとは限らない。
行為者にとっての「意図」とは、「自分がその行動を行った原因」のことであり、主観的な経験とその報告を前提とするため、この意味での意図はヒトでのみ議論することが可能となる。しかし行為の観察者から見た「意図」としては、自然選択の結果として非自発的な行動が引き起こされた際の原因として捉えることが出来る。
動物行動学では、ある行動を行う際にそれに先だっていつも行われる運動のことを「意図行動」と呼ぶ(Heinroth 1910)。たとえば、オオカミが「噛みつく」という行動をする前に「牙を剝いてみせる」という運動をする。この意図行動は他のオオカミが次に起こる行動を推測するシグナルとして働く。このようにして動物コミュニケーションの文脈では意図が動物でも議論されている。
また、心の理論の文脈では「意図性の共有」という言葉で、ある個体が協力的な相互作用の際に他の個体と心理的状態を共有することを指す(Tomasello & Carpenter 2007)。このような意味での「意図性の共有」が人間以外の動物で可能かどうかはまだ明らかでない。(619文字)
2013年06月05日
■ 行動科学での表象と神経生理での表象
11/12
シンポのときあらためて考えたけど、鮫島さんのScienceでaction valueをニューロンがコードしていると言うときは個々のニューロンが左右どちらかのQを表象していて、その情報を使ってさらに下流で意志決定しているというモデルになってる。
つまり、Qじたいは明示的に表象されていなくて、端的にそれを元に左右を選択するための閾値だけを上げ下げすることでも同じ問題は解けるのだろうけど、その可能性を神経活動からunlikelyだと議論できる。
専修シンポで鮫島さんに対して真ん中のあたりの席で質問してた人への答えとしてはそういうあたりが答えかなあとか考えた。
これはモデルベース云々とはたぶん違う話なんだろうけど、どの段階まで明示的な表象を持っているか、というあたりは行動科学でやってる人と神経生理でやってる人とでぶつけ合って議論するに足る話題かなとか考えてた。
つまり、連合学習理論でいろんな方法使ってやってることは、動物がどのレベルの表象を持っているかを行動レベルで示してやろうとすることなのかなと。このあたりがマーのアルゴリズムのレベルと実装のレベルって議論の実例なんではないだろうかとか考えた。
.@KazuSamejima ありがとうございます。「どのようなレベルであれとにかくデコードできるような情報を持っていること」と「明示的にスパースにneural correlateがあること」とはどう違うんだろうかとか考えてました。行動で見る際にはデコードさえ出来ればいいわけで。
2013年04月11日
■ レスコーラシンポのために予習
2012/11/8
レスコーラシンポのために予習としてRWモデルの理解(<-そこから?)。読んでて楽しいのは、salienceとかsurpriseとか、わたしがこれまで聞いてきたようなキーワードが出てくること。
すると、ここで言ってるsurpriseをIttiのベイジアン・サプライズで置き換えらんないか?なんてことを考える。連合学習理論の本質がなんなのかまだ私には分かっていないのだけれども、もし究極には「連合強度の上げ下げ」を扱うかどうかであるなら、ベイジアンな連合学習理論もありうる。
つまり、複数のCS1, CS2の出現頻度とそれぞれでのcontingency P(US|CS1), P(US|~CS1), P(US|CS2), P(US|~CS2)を逐一「推定」して行動判断をする過程があれば、そのときの「連合強度」を計算することが出来るだろう。
そのときの更新則がサプライズ(連合強度の次元での引き算)になっているのがRWモデルで、この更新則にベイジアン・サプライズを使えないかってこと。(ベイジアン・サプライズ = posterior beliefとprior beliefとのKL距離)
"associative learning" bayesianでググってみたら、John K. KruschKeの論文がみつかった。("Doing Bayesian Data Analysis"の人)
あと、Fristonはどうかというと、本人筆頭の論文ではRW1972をSutton & Bartoと並べて言及するだけでそんな深いことを言ってない。共著ではCerebral Cortex 2009、 あとスライドでこのあたりに出てくる。
ともあれ、ここまで極端に振ってみると、ベイズ的になんでも全部取り込んで確率分布として表象しちゃう方策と、必要最小限のことだけ表象する方策(そこで「連合強度」を使うのが連合学習理論)とでそれぞれに学習をして行動を最適化をすることが可能だと言える。これで毎度のベイズ脳批判に到達する。
あとはどこまでそういった表象を持っているかを調べるというempiricalな問題になる。
ここまで考えると、データドリヴンでの連合学習モデルの形成とかも思いつく。つまり、CS1, CS2, US1, US2くらいのパラメータがあって、連合学習で知られているあらゆるスケジュールは上記パラメータ間の時間的関係として記述される。
それで、これらのパラメータとスケジュールを振った上で、CR1, CR2のデータ(それぞれUS1, US2にたいするもの)を収集する。これらのパラメータとスケジュールとCRの関係をうまく説明するモデルを機械学習で作ってやる。(<-出たっ機械学習信仰)
そのとき「連合強度」とよべるような潜在変数が出てくるかを問う。もしくは既存のモデルでの連合強度をモデルに入れてやるとモデルの自由度が効率的に下げられるかどうか検証するとか。
CSもUSもなんらかの刺激だから本質的には差がないので(じっさいSOPではそうなっている)、もう少しパラメータ数を減らしてやることは出来るかもしれない。
一般化すれば、P(R1(t),R2(t),...) = f(P(S1(t),S1(t-1),...,S2(t),S2(t-1),...))となるわけだから。
こういうのもどっかにあるか。ググってみたが、data-driven "associative learning"では見つからなかった。
いろいろ外している気はするが、それでもとにかく書いてみた。
明日の予習用にと"Pavlovian Conditioning: It's Not What You Think It"を読んだ。よく構造化されているし、かなりかみ砕いて書いてあるので読みやすい。
まとめておくと、1988年の段階でも、教科書的なPavlovian Conditioningの説明というのは連合学習理論の進歩をちゃんと反映していない。
1) temporal contiguityはconditioningの必要条件でもなければ、充分条件でもない。(P(US|CS) vs. P(US|~CS)の関係やblockingの例を出して、動物含むわれわれ有機体はinformation seekerなのだと強調する)
2) 単純な刺激CSとUSの間を連合するものだけではない。RHというcompoundとR,H単体とを別々にconditioningする例を出して、この連合によって単純なものから複雑な概念を作ることが出来ることを強調している。(これがたぶんHumeに繋がる)
3) CSはUSが引き起こした応答だけを誘導するわけではない。たとえばCSがlocalizedであるかdiffuseであるかによって引き起こす応答は違いうる。ゆえに、あるCSによってUSで見える応答が起こさなかくても、べつの応答に反映する可能性がある。
ここまで。1)に関しては「学習心理学における古典的条件づけの理論」で読んでだいたい分かっていたが、それ以外にももっといろんな論点があって面白かった。たとえばorganismのexisting structureの重要性なんてのは、すごくエコロジカルだ。
なぜ急にこの分野に勉強しているかというと、連合学習理論はじつはすごく表象主義的なので、反表象主義的なsensorimotor理論とぶつけて考える意義があると思うから。一方で、徹底的に擬人化を避けるという意味では両者は共通していて、認知神経科学とはちょっと違ったところにいるようだ。
明日は"Contemporary Study of Pavlovian Conditioning"を読んでみることにしよう。
自分の興味の向き方を考えてみると、現象学的経験自体には興味があるのだけれども、心の理論とかミラーニューロンとかはどうも擬人的すぎてなじめない。身体性には興味があるが、社会性はよく分からない。あと、いつまで経っても「言語」に興味が向かない。良くも悪くもいまのところそんなかんじ。
つまり、意識経験は重要なんだけど、人間的であること(概念、言語、…)にはなぜかあまり惹かれない。非人間的な意識科学者。ってまさに自分がやってきたことか。
2012/11/9
.@kosukesa おつかれさまでした。新幹線の時間があったのでレスコーラの若手へのメッセージを聞いたところまでで失礼させていただきました。
レスコーラのトークは「compound stimuliを使うとあれも出来るこれも出来る」みたいな話で、神経科学者としては難易度高かった。昨日読んだAmerican Psychologistをもし読んでいなかったら、「そもそもABはAB->USなのかA->US, B->USなのか」ってところからつまずくところだった。
レスコーラのメッセージでいいなと思ったのは"Identify critical difference among accounts"(そのためにきっちり統制実験しとけよ)ってのと"Find the improbable prediction"(インパクトのある仕事になる)の二つ。
こんかいのしゅっちょうでは、アイパッドミニをじっさいにてにとってかくにんすることができたのがよかったことです(<-小学生の感想文)
2013年03月21日
■ 学会やセミナーどう質問したらうまくいくだろうか?
2012/9/16
質問をするときに、いかにクドくしないかは考える。放っておくといくらでもクドくなるから。「あなたかもしくは他の誰かがこれこれこういうことを確認していますか?」は「あなたはこれこれこういうことを確認していますか?」で充分。ほかの誰かがやってるなら後者でも答えてくれるから。
あと、前置きを言わないようにする。「ナイーブな質問かもしれませんが」とかは情報がないので単刀直入にいくべき。
「素晴らしい講演ありがとうございました。質問ですが…」もやらないことにしている。他人にやるなとまでは言わないが。どうやらこれは学会の文化に依るらしい。儀礼的なのは好きでない。そういうのは懇親会で個人的にやるべきだと思ってる。
質問の真意を説明する前に、具体的な問いに落とし込む。頭のいい人だと質問の真意を汲んでくれるし、そうでなかったら畳みかければよい。
今日はGrégoire Courtine (最近Scienceに論文出した)のセミナーだった。脊髄損傷ラットのFESによる機能回復の話。
素朴な疑問として「トレッドミル条件で尻尾を支えるのはなんで?」と聞こうと思ったのだけれども、もうちょっと質問を組み立てることにして、「尻尾はロコモーションにはどのくらい寄与するの? トレッドミル条件では支えているようだけど、overground条件では支えてない。両者の差は尻尾を活用してるかどうかなのでは?」としてみた。けっきょくクドいw
結果論だけど、Grégoire Courtineはむちゃくちゃよく分かっていて、一聞けば十返ってくる人だったので、この人に対しては「トレッドミル条件で尻尾を支えるのはなんで?」と聞けば充分なのであった。
素朴な疑問が浮かんだときにそれだけ質問すると話が広がらなくてつまらんので、ちょっと質問を組み立てることを考えるようにしている。高等研の研究会のときには「contagious yawningでは実験者があくびをするけど、それはvoluntaryなものだから、fake yawningだと思う。」とコメントした。
さらに、「どうしてfake yawningで充分contagious yawningが起こるのだろう?」と聞けば話が膨らんだかもしれないが、そうではなくて、つづきを「real yawningでcontagious yawningを調べたものはないのか?」というふうに問うてしまった。これはよくない。
後者だと「そんなものない」で終了してしまう。質問の意図は汲んでもらえず、議論も盛り上がらなかった。前者だったら、contagious yawningを起こすのに必要な要素は何か、みたいなもうすこし本質的な問題に届いたような気がしたんだけど。
クドくはしたくないが、つまらん質問にもしたくない。そんなことを考える。あともちろん、clarificationのための質問と、議論をするための質問は別で、今は後者の話をしている。あと、ラボセミナーとかで、講演者がちゃんと分かって喋ってるかどうかを確かめるための質問とかもある。
つまらん質問にならないように質問を組み立てて、「これに答えられないとお前の主張はひっくり返るよ」という風になるようにして、必死になって答えてもらえるように心がけているのだけれども、なかなかうまくいかない。学会でもラボセミナーでも。
2013年03月16日
■ スライド用に使える解剖学の図を探してみた
2012/11/1
いろんな動物での脳の大きさを比較した図を探していて、これがいいと思うんだけど、ソースがわからない。いろいろ探したらHubel (1979) Scientific American 241(3) 44-53であることが判明した。
{kind=link}
上丘(superior colliculus) = 視蓋(optic tectum)がほ乳類では小さいけど鳥、両生類、魚類では大きいって絵はこれがよさそう。 だがふたたびソースが不明。(google画像検索済み)
しかたないのでほかを当たったら、Integr. Comp. Biol. (2002) "Understanding Vertebrate Brain Evolution" これのfig.1を発見。これはソースが書いてある。これらだそうな:Braun, C. B., and R. G. Northcutt. 1999. Morphology: Brain and cranial nerves. In R. Singer (ed.), Encyclopedia of paleontology, pp. 185–192
"What is the thalamus in zebrafish?" これすごくいい。Fig.8見るとメダカでは視床のどこを介して感覚入力が入るのかだいたいわかる。メダカではtectumから新皮質への投射は無し。PGIを介して扁桃体に行く。
{kind=link}
Fig.8でのanterior thalamus (A, green)がたぶんLGNの先祖なんだろうってのは以前比較解剖学関連の教科書を読んでて考えた。弱い根拠だが、tectum -> Aの投射はヒトからサカナまで保存されてる。
だが本文読んでみたらこの考え方には否定的で、なぜかというとanterior thalamusからの投射はGABAergicなんだそうな。glutamatergicなのはもっとcaudal側のthalamusにある。これは説得的だ。著者の結論はサカナにLGNの先祖はないというもの。
同様にして、pulvinarの先祖もない。ただし、PGIって核を介してtectumからamygdala, hippocampusへの投射がある。これは面白い。
Pulvinarはヒト、サルにはあるけどラットではほとんど無い。ではその先祖は何かっていうとLGN含むdorsal thalamusがそもそも新しい核であって、それが分化したらしい。Puellesの"Thoughts on the development, structure and evolution of the mammalian and avian telencephalic pallium"のfig.5参照。
つまり、渡辺茂先生の二重乖離の実験にあるように、トリのN. rotundusはヒト、サルのPulvに対応すると考えられている。これは機能的にはそうなんだろうけど、系統発生的にはたぶん違ってるということが上記のPuellesのfig.5からわかる。
いま言及した比較解剖学者のLuis Puellesのwebサイトを探していたら、アレン・インスティチュートのAdvisory Councilsのページがヒットした。関係者どんどんinvolveされているってことがよく分かった。
2012/11/5
以前言及した"What is the thalamus in zebrafish?"で味わい深いのは、Fig.1の系統分岐図でzebrafishが中心になっているために、その近隣の種が拡大されていて、ふだん見ているのと違っていること。
オーストラリア中心の上下反転世界地図(リンク1、リンク2)を見たときのような、視点の変更による驚きと喜びがある。
系統樹による大脳、中脳、間脳、小脳の大きさの違いを図示したこの図もよく見るけどソースが見つからなくて(カラバイだって書いてあるが)難儀していたんだけど、いろいろ探してCurr Biol. 2005 15(23):R946-50 "Evolution of the avian brain and intelligence"であることを突きとめた。
{kind=link}
2012/11/7
The Retinotopic Organization of Striate Cortex Is Well Predicted by Surface Topology この図はイイ! V1損傷の話でretinotopic mapについて説明するときにちょうどいい図がなかったのだが、こんどからこれでいける。
ちなみにmacaqueの場合は古典的な論文でDANIEL and WHITTERIDGE (1961) J Physiol. 159:203-21というのがある。電気生理から再構成。fig.5,6では折り紙wを使って構造を説明。
もっと時代の新しいところでは、Tootellが物理的にunfoldしたV1で2DGを使って作った J Neurosci. (1988) 8(5):1531-68. "Functional anatomy of macaque striate cortex. II. Retinotopic organization"
マカクでHortonでいい図を選ぶとすると、J Neurosci. 1996 Nov 15;16(22):7228-39. "Intrinsic Variability of Ocular Dominance Column Periodicity in Normal Macaque Monkeys"だろうか。
マカクでどうやってV1をunfoldするかは先述のJNS1996 16(22):7228のfig.1にある。味わい深いのは、このテクを使った初期の論文JNS1996 16(5):1791ではnewbornでも綺麗に開けていないが、1998 18(14):5433ではほぼ完璧であること。だんだんうまくなってるっぽい。
盲視で二つの経路議論をするときにトリでも同様の二つの経路があるって話をするときには"Hemispheric asymmetries: the comparative view"これの図1がよさそう。
{kind=link}
2013年02月08日
■ 「無を表象する」ことと「表象がない」ことの違い
2012/10/18-19
なんにも言わずにいるというのは難しいことだ。「無を表象」しているつもりなんだけど、「表象がない」と理解されるだけだからなあ。
つまり、「無を表象」するためには、普段しゃべり続けている人がふと黙り込まないといけない。つまり時間的前後とのコントラストによって表象される。暗い場所でも周りが明るければsalientであるのはこれの空間バージョン。
そこまで考えた上でもう一度見直してみると、でも「無を表象する」のと「有を表象する」のとが非対称であるはずもない。
以前因果推論のあたりを勉強したときに、澤さん経由で逆行性ブロッキングもRWモデルで説明可能であるということを知った。つまり「Tが出ていないということが連合強度を下げる」というやりかたで。でもこれはまさに「無の表象」だ。
あらゆるほかのものが提示されていないのにそれらの連合強度を潜在的に持っているとでもいうのだろうか? なんかフレーム問題みたいなことが起こっているような気がする。
.@kosukesa おっしゃるとおり、RWモデルそのものではありませんでした。
.@kosukesa ブログのときの表記を使うと、トレーニング1で LT->F のあとで トレーニング2で L->F をすると、トレーニング1のときにL-Tの間の連合が活性化されているから、トレーニング2でTの不在がシグナルとなる、こういう理解で正しいですか?
.@kosukesa ありがとうございます。そうなると、不在のものはなんでもかんでも連合強度が負になる、みたいな変なことにはならないわけですね。
するといまの例も「しゃべり続けていた人がふと黙ることで無を表象する」の範疇に入ると言えそうだ。つまり、記憶を持たなくても(統計的性質を表象しなくても)、時間的変化をモニタするだけで可能。
.@kosukesa ありがとうございます。「フレーム問題的な話題」自体はあるのですね。
今日は”learned irrelevance”というキーワードをゲットした。
即行”learned irrelevance”でググって日本語のもの探したら、北口勝也さんによるものが多く見つかったが(たとえば動物心理学研究)、いちばん最初に出てきたのはまたもや村山航さんのレジメだった。
2013年01月19日
■ カーゴカルトサイエンス
「カーゴカルトサイエンス」の章を読みたくて「ご冗談でしょうファインマンさん」を再読してみたんだけど、記憶にあるのよりもっとまっとうだった。もっと物理帝国主義全開で動物心理のことをバカにしてるかと思ってた。でもそうではなくて、ラットの行動に影響を与える手がかりをきっちりなくすという話で、科学的にまっとうなだけでなく、けっこうフェアだった。
ググったらすぐ原文見つかった。ファインマンのほかのエッセイもこのカルテクのサイトからいくつかアクセスできる。
ファインマンの文章に出てくる、ラットの実験のYoung 1937 (ラットが迷路を解く実験で、ラット自身の足音が手がかりになっていることを、床に砂を撒いた条件で確認したとのこと)ってのを探してみたのだけど見つからない。ファインマンのこの文章だけが検索で引っかかってくる。
ただその過程で見つかってきたのが、"Think Like a Rat"(pdf)。この文章では、ファインマンのカーゴカルトの章を引用した上で、ラットのplace cellは部屋を真っ暗にしても活動するが、床を拭いてラット自身の臭いを除去したらplace fieldが不安定になったって仕事 Hippocampus 2000をまさにこのファインマンの話の実践だと書いてる。
2012年12月20日
■ 計算論的モデルを使うということについて実感に基づきつつ。
2012/9/11
いままで自分が関わってきた計算論モデルは、サッカードの応答潜時をモデル化するdiffusion modelと、saliency mapという計算論的、心理学的な仮構物を中間に置いてボトムアップ注意をモデル化するsaliency model。
どちらとも、biologicalにplausibleであることに重点を置いて、たくさんのパラメータを入れ込んで行動データを予言するのに成功していたものを後から利用する立場だったんだけど、post-hocにパラメータどんどん突っ込んでその予言性能を上げるというのに違和感を持った。
当時はまだベイズ的なものを十分理解していたわけではなかったけれど。それで、どちらのときも不完全ながらモデル選択的なアプローチをした。
Diffusion modelの場合には解釈可能なパラメータ以外をなるたけ減らした上で、二つの条件であるパラメータthetaが等しいモデルと異なっているモデルとでAICを計算して、パラメータ増えたことを考慮してもthetaをべつべつにしたほうが良いモデルであるということを示した。
Saliency mapのほうはモデルを簡略化することは状況的に不可能だったから、チャネルの寄与をleave-one-outを使って評価した。こっちはcross validationはしていないし、いろいろ足りなかったが、やりたかったのはそういう方向性。どちらも、なんでもパラメータ突っ込めばいいというわけではないというのが基本的なアイデア。
そんなわけで、この方向性を突き詰めて、モデルベースで行こうとしたら、 たぶんベイズ的になるんだろうと思ってた。いっぽうで、この種のモデルはモデルの中でしか話ができない。たとえばsaliency mapの話で行けば、あれはもともとV1を経由した皮質での視覚情報処理のモデルなんだから、V1 lesionの話をこれで説明しようってのが端っからズレてる。
つまり、もっとぜんぜん違うモデルを持ってくれば予測性能が上がる可能性はまったく否定できない。Diffusion modelのほうも同じで、パラメータをthetaとmuの二つに絞っちゃったので、二つのグループの違いはこのパラメータの違いでしか説明できない。
もちろんモデルがまったく間違っているのならパラメーターの変動でデータを説明することがそもそもできないけど、もっといいモデルがあった可能性はある。以前も書いたけど、diffusion modelを含むaccumulator modelというのはevidenceを蓄積する。
でも、蓄積しないである時点でのevidenceの大きさが閾値越えるかどうかでサッカードするってモデルでも良い。ふつうはこういうモデルは有効でないけど、V1 lesionしたときにそういうものが有効である可能性はある。
まとめると、モデルベースだとモデルの空間の中で一番ましなものを探索しているだけということになる。まあ、そういってしまえば当たり前な気がするが、パラメータ空間だけ見てると、今言ったような蓄積モードかその時々モードかとかそういうものが見えなくなる。
そういうわけで、モデルベースでやることの限界と帰結みたいなものを自分なりの経験に基づいてまとめてみた。
こういうことに目が向く前は、統計でANOVAかけたとか、ここは対応があるデータだとか、多重比較がどうのとか、多重共線性がどうのとかそういうことに多大な興味があったが、モデルベースで考えるようになったらなんかどうにも間接的すぎる気がしたものだった。
神経生理の方と話していて、基本はt検ですよ、と主張するのを聞いてて、気持ちは分かる、たしかにそういう美しい実験デザインを汲んでみたいものだと思うけど、でも自分はずいぶんと隔たったところへ来てしまったなと実感した。
そんな自分としては、いまは信号検出理論とか見てるとあまりに原始的でもちょっとなんとかならんのかと思う。すくなくともガウス分布は勘弁してほしい。
そんなこんなで、あらためてこう書いてみると、自分がこれほどまでモデルベース思想だったかと驚いた。私はどっち向いて進んだらいいんだろう、と途方に暮れた。
ぜんぜん違うモデル、なんてあり得るかって議論はあり得る。たとえばサリエンシーモデルはもともとTriesmanのfeature integration theoryから来ているので、featureのチャネルはそれぞれ独立して計算されて、後で足しあわされる。当然、このようなパラレルな処理がいつも厳密に行われているとは限らない。でもそういうのは、featureののinteractionのチャネルを付加してやればよい(たとえばmotion * color)。けっきょくモデルを拡大して、パラメータを増やしてやることになるだろう。
同様にして、diffusion modelも蓄積モードと瞬時モードの両方を計算しておいて、必要ならどっちか荷重見かけてやればよい。こうして、モデルの柔軟性を上げることは、モデルの肥大化とそれに伴うパラメータ数の増大によって実現される。そうなると問題は「はたしてすべてのあり得る可能性を取り込んでいるのだろうか」というなんかフレーム問題みたいなものに帰着する気はする。そこまで行ってしまう前に、そもそもモデルの膨大かでは対処できないような致命的なモデルの改変というのはあるだろうか?
たとえばIttiのサリエンシーモデルは片道通行なのが特色で、Tsutosとかのものと比べると違っている。それすら、あらたに逆向きの流れを付加してやれば対処可能な気はする。とか書いているうちにだんだん分かってきて、これを推し進めるとけっきょくパーセプトロンからバックプロパゲーションに至ったニューラルネットワークと同じ道なのだな。なんという車輪の再発明w
2012/9/12
そういう意味であれば、数時間前に書いたdiffusion modelとsaliency mapを題材に書いた話とずれてはいないか。モデルを作っているときはそういうこと考えたりするけど、でも、論文を書いて戦っているときに「贋作だよな」という感覚をレフェリーと共有した経験はない。
いや違うか。「これこれこういうモデルの中で最適化したパラメータはこういう挙動を示します」みたいな言い方に留めておいて、もっと違ったモデルがあること自体は否定しないように書いているか。いまのところ、モデルをゼロから作ったわけではないのでそういう場面にはまだ遭遇してないんだろう。
「サリエンシーモデルはV1があること前提なのに、V1損傷のモデルをチャネルのゲイン変えるだけで対応っておかしいだろ」て言われたらどうしようかと事前に考えたけど、いままでに指摘されたことはない。そう言われたとしても「もっと良い(正しい?)モデルがある可能性は否定しません」でよいか。
「原理drivenで微分方程式で動かすようなモデル」このフレーズを見て、それからさっきの「お話としての説明」「物理学的な説明」 http://pooneil.sakura.ne.jp/archives/permalink/001364.php と突き合わせてみると、さっき書いたことは転倒していることが分かる。
つまり、認知科学的なモジュールを置いた説明が「お話としての説明」であって、バイオロジカルモデルが物理的因果で起こす現象が「物理的説明」で、「正しいモデル」というならそれは後者。前者こそが「贋作」であることを分かった上で使うべき仮構物。
Diffusion modelの話でいけば、サッカード開始のために「閾値を越える」というが、そんな閾値は仮構物。実際に起こっているのは、ニューラルネットワークの中でSelf-organized criticalityが出来ていて、どっかから雪崩が始まるということ。
以前diffusion modelをマーの三段階に当てはめようとしたことがあったけど、 http://pooneil.sakura.ne.jp/archives/permalink/001282.php あそこで書いたのと同じ。つまり、閾値はimplementationのレベルではなくて、もう一段抽象度が上がってる。
ニューロン間の相互作用(プロセス)があって、それのSOCとしての状態(なんか温度みたいなもの)をあらわそうとすると閾値(表象)になる。かくして表象とプロセスとのやりとりが抽象度をレベルを上下する。ここが「ベイトソン流に解釈する」って書いたところ。
だから、データドリブンにデコーダ作ってベイズの法則でひっくり返して(<-簡単に言いやがる)生成モデルを作るのも、認知心理学的な知見からモジュールを想定して作るモデルも両方とも贋作。すべてのニューロンの発火とシナプス伝達からなる力学系が「真作」。それは記述の抽象度を反映している。
ネットワークの状態(プロセス)を閾値として表象するとき、それは他者による観察ではあるけれども、客観的なものであるとは言える。
つまり、閾値は温度みたいな統計物理的な量に対応するから、ミクロでは定義できない。1個のニューロンの発火の閾値イコールネットワークのバーストの閾値ではないということ。ともあれ、物理量だから、まだここは物理的因果が働く世界であって、認知モジュール間の文法の世界ではない。
閾値はそういった統計量である一方で、蓄積されるエビデンスの大きさは発火頻度だから1個のニューロンの量のようにも見える。でもそうすると抽象度の違うものの間でのモデルになるからなんかおかしい。エビデンスは一つのニューロンでも集団でも質的には同じように見えるからだまされるのか。
2012年12月03日
■ 甲状腺刺激ホルモン放出ホルモン(TRH)のスプレー式点鼻薬
甲状腺刺激ホルモン放出ホルモン(TRH)のスプレー式点鼻薬 こんなことしてだいじょうぶなのか?TRH->TSH->T3,T4->TRH,TSHへネガティブフィードバック、となっているんだから、点鼻しないとTRH出ない事態になりそうだけど。
元ネタ( ArmyTimesおよびIndiana University)読んだが、点鼻スプレーを開発するというプロポーザルにお金が下りた、ということか。まあ、オキシトシン点鼻薬とかと同じ発想ということかな。
TRHを脳内投与するとratではarousalが起こる。これは甲状腺への作用ではなくて、オレキシン産生ニューロンを刺激することによるらしい。( JNS2009および JP2009) (ほかの系へもいろいろと作用することが知られている。)
すると、バセドウ病で自己抗体がTSHの代わりに甲状腺を刺激してT3,T4を産生、これが発熱や頻脈などを起こす。一方でT3,T4のネガティブフィードバックによってTRHは下がる、これがオレキシン系経由でarousalを下げてちぐはぐな状態になる、というところなのか。
ああ、もっと基本的な知見として、TRH ->NAc -> DA放出 -> locomotor activityという経路があるのか。 Neuropharmacology. 1979 および Eur J Pharmacol. 1981
2012年11月10日
■ 妄想(delusion)と幻覚(hallucination)の違いについて
Keisuke Suzukiさんとのツイッターでのやりとり(20120717)。
ksk_S 誰と話してたときだったか忘れたけど、ラバーバンドで元の手の温度が下がるのも面白いけど、ヒスタミンレベルまで下がるのはすごいねという話がでた。認知における自他が、免疫における自他と一致する必要性はないのに。
ksk_S あと、ちょっと不思議なのが、身体所有感や、運動主体感、自分の声や、思考ですらも自分に属してる気がしなくなる「妄想」が起こりえるのに、視覚にはその手の誤帰属が起きるという話がないということ。Derealizationはそれなのか?ちょっと違う気もする。
pooneil @ksk_S たとえば「今見ている視覚像はわたしではなくて別人が見ている」という信念ってこと? たしかにあり得なさそう。>> 視覚にはその手の誤帰属が起きるという話がない
ksk_S @pooneil その通りです。視覚は自らが視野の中心であるという感覚に揺るぎがないような。metzingerのOBEの議論でも、弱い一人称性は常にカメラのある位置に立ち上がる。
pooneil @ksk_S 身体性にもとづくagency自体はTPJとかあのあたりで作られるのだろうけどとにかくそっちはいろいろ揺らいで、外界にそのcontentを投影しているだけっぽい視覚の方は揺らがないというのは、あたりまえのような、そうでないような、面白い事態な気がします。
pooneil 「我思う、ゆえに我あり」ではなくて、「我見る、ゆえに我あり」?
pooneil 視覚では視覚のcontentはいろんな錯覚で揺らぐのと同じように、agencyは体性感覚や固有感覚でのcontentなので揺らぐ、と整理できる? 視覚では見る側が揺らがないということではなくて、端的にそれは視覚でのcontentとなっていない(視覚にとって外部である)ということ。
pooneil でも変かな。われわれが眼や頭を動かして視野像を繋いで視覚世界を作るというときにはその自己の運動は視覚にとりこまれているわけで、ほかでもない「わたし」が眼や頭を動かしたのであって、誰か他人ではない(とまでいかなくても、世界が動いたのではない)、という情報は持っている。
ksk_S @pooneil 身体性の感覚はagencyにしろownershipにしろ揺らぎますね。外界の投影としての視覚は、常に「外」なので揺るぎなくて、主体としての自己と帰属された自己が入り交じる身体感覚は、混乱しやすいということなんですかね。
pooneil @ksk_S そっか、「主体としての自己と帰属された自己が入り交じる」ここが今の話の本質ですね。
ksk_S @pooneil 何がcontentかを考えると整理できそうな気がしますね。気になるのは視覚はhallucinationは起こるけど、delusionalにはならないという点。身体の帰属自体がある種のdelusionだと考えると、それも自然な気もしますが。
pooneil @ksk_S そもそもhallucinationとdelusionの違いが分かってないのだけれども、hallucinationは知覚のcontentのレベルで、delusionはその知覚を元に判断(表象)している表象部分? 妄「想」っていうくらいだし。仏教でいう受と想の違い。
pooneil いや、ちがう、端的に日常言語的に、hallucinationは知覚のレベルで、delusionは思考のレベルだと理解していたのだけれども。
ksk_S @pooneil hallucinationは本来ならそう知覚されるはずべきものとの「ずれ」、delusionは世界とはこうあるはずものとの「ずれ」。低次か高次かという違いな気もするし、delusionは何か暗黙的で大きな世界観とのずれが際立っている印象。
pooneil @ksk_S なるほど、そこでprediction errorの出番ですね。どのスケールでのエラーなのかの違いか。
ksk_S Antonio Damasio, et al "Persistence of Feelings and Sentience after Bilateral Damage of the Insula"
ksk_S insular失ってもfeelingは残るのか
2012年10月22日
■ 赤ちゃんであるとはどのようなかんじか?

(20120901)「哲学する赤ちゃん」(アリソン ゴプニック)を図書館行って借りてきた。因果推論の話はいちばん初めの章「子どもはなぜ「ごっこ遊び」をするか?」のところで、子どもが反実仮想をすることに重点を置いているようだ。
あと、第4章のタイトル「赤ちゃんであるとはどのようなことか?」は"What it is like to be a baby"の訳だと思うけど、毎度ながらわかりにくい。「どんなかんじか?」とかにしてくれないと意識経験という一人称的な話であることが伝わらないと思う。
Alison Gopnikは、Ned BlockのBBSの記事へのコメントで、「赤ちゃんは我々大人よりももっとconsciousである」なんて言ってる。
つまり、赤ちゃんはボトムアップ注意はあるけどトップダウンはないのでサーチライトのような注意を使わずに全方位の(たとえば視覚の)刺激を経験している、だから「よりconsciousなのだ」という議論。ほんとか?と思う。全体的に薄められる、周辺視みたいな経験になるんじゃん?と思う。
あと、ボトムアップでもサーチライト的に注意は向く。というかそれがサリエンシー・マップだから。というわけでいろいろつっこみたいところはあるが、ともあれ赤ちゃんでわかったことと動物でわかったことを並べて考えるというのは基本なので読みながらいろいろ考えてみることにする。
眼球運動を使って、解像度の高いfoveaの部分を動かしながら統一性のある視覚像を造るという過程があって、それを赤ちゃんがどうやっているのかというのには興味があるが、そういうことをやろうとしている人はいくらでも居るだろうから、なんか自分の強みで出来ることはないかと考える。
そもそも眼球運動によって別々の網膜像を「統合する」ってなんだろう? 統合して一つの絵を作るって考え方自体はカルテジアン劇場のヴァリエーションなので、そんなことしてないって話になるかもしれない。計算論的には眼球の位置の情報を持って解像度の良いところだけ貼り合わせるとかは可能だろう。でも、貼り合わせないで、とにかく行動に使えれる分だけread-outしていると考えた方が尤もらしいのではないだろうか。ともあれこれはempiricalな問題だ。
2012年09月30日
■ こまごまとメモ。(20120521まで)
あとからピントを合わせられるlight field cameraの話に興味を持つのは、これがAdelson & Bergen 1991 (PDF)のPlenoptic Functionを記述しようとするものだから。
Adelson & Bergen 1991 はそれまでのmotion energy modelとかbinocular disparityとかいろんなものを統一的に扱えるという意味で視覚科学/コンピュータビジョン的に重要だ。
でもそれだけではなくて、わたしが重要だと思っているのは、これがギブソンの包囲光配列(ambient optic array)を数学的にcompleteに記述したものだから。これはわたしの妄想ではなくて、Adelson & Bergen 1991の本文にも書かれている。
ギブソンの本を読むとさっぱり数学が出てこないので、かえってわかりにくくて困るのだけど、しかもそれは反表象主義的な態度から来ていることも推測されるのだけれども、でもPlenoptic Functionでちゃんと「不変項の抽出」とか扱ってやればいいんじゃないのか?とか思う。
なぜかはよくわからないけど、Adelsonは1992のplenoptic camera以降はplenopticという言葉が入った論文を出してないし、このへんの問題意識を後継しているものが見つからない。見逃しているのだろうか?
アフォーダンスが脱臭されてgraspingとかの行動選択の情報として扱われるのと同じように、包囲光配列も数学的に扱うだけで脱臭され、それの含意する「直接知覚」的な立場は危うくなってしまうのだろうか? もうちょっと考えてみることにしよう。
"The Cost of Accumulating Evidence in Perceptual Decision Making" JNS2012 こういうかんじにaccumulator modelで時間が経つごとに閾値を下げてゆくというのはわたしもV1 lesionでdiffusion model使ったときに考えたけど、複雑になるだけだから避けてた。
いまだったら、deliberateである=accumulationする、reflectiveである=accumulationさせない、という感じで論じることもできそう。
つまり、normalではLIPを使ってevidenceのaccumulationができるのだけれども、blindsightでは蓄積できない(SCで積分でなくシグナルそのもので閾値を切る)=>いつまでたっても確証がない=>reflexiveになる。
サルではスピードのコストが高いので応答潜時は早くなる。ヒトではスピードのコストが低いので応答潜時はかえって遅くなる。そして、この「確証を与える」という部分こそが意識の機能であり、メタ認知との関連性に関わるとか云々。
蓄積できるということは時間をまたぐということなので、そういう意味では意識に時間の流れが必要なのはたしかで、でもそれは作業記憶みたいなものとは違うだろう、って話の流れ。
この話(deliberate=accumulation)はAndy Jacksonと議論しているときに考えた。Andyはそのときたしか、thresholdが時間で一定だってのは妥当な過程か?と聞いてきたはず。そのときは鋭いなあと感心したのだが、Andyはシアトルにいたので、Shadlenのこの仕事の話を聞いていたのかもしれない。
"Effects of Visual Experience on the Human MT+ Functional Connectivity Networks" Front. Syst. Neurosci. 2010
congenital blindでもMTはtactileとかで活動する。だから、V1スキップしてMT活動したときの感覚はvisualというよりはもっと抽象的なものかもしれない。
眼球運動の定量的な取り扱いでは網膜の座標だとspherical coordinate (radial distance, elevation, azimuth)になって、筋肉の座標だと外眼筋で三軸あって、オイラー角でroll-pitch-yawに対応する。
オイラー角なじめなかったけど、飛行機の進む向きで考えればよいのか。進行方向で上げ下げするのがpitchで、進行方向の軸で回るのがローリングで、左右に曲がろうとするのがヨーイングか。おぼえた。
そうなると、上丘での網膜依存の座標(spherical coordinate)から外転筋の座標(オイラー角)への変換となる。ただし、torsionはあんまり使わないから、pitch-yawだけでコントロールする。
眼球運動のコントロール自体にはあまり深入りせずにきたけど、軌道の議論とかをちゃんとやろうとするとこのへんを避けるわけにはいかない。
ジンバルロックとかなにその名前、かっこよすぎる。(<-老いて学び、なんだったっけ?)
2012年09月16日
■ モティベーションのサリエンシー
Peter Redgrave以降の仕事でDAがvisual salienceに関わるという話(Nat Neurosci. 2003)があって、SNcのDAニューロンのshort-latency visual responeは上丘からのvisual salienceのシグナルを受けている。
最近の彦坂研の仕事(Neuron. 2010)でも、DAはprediction errorだけではなくて、"motivational salience"をコードしているんだという言い方をしている。
この"motivational salience"って表現が絶妙で、Kent C. Berridgeの言う"incentive salience"って言葉と微妙に変えてあって、同一視されないようにしてある。
ともあれ、visual salienceとincentive salienceを包括して考えられるんではないだろうかと考える。このことはさらに、saliencyとconsciousnessとの関係について考える際にも役に立つはずだ。
Incentive salienceの方から考えると、Liking(=pleasure)とwanting(=incentive)との分類であれば、wantingのほうは無意識のnovelty detectionだけで可能で、でもlikingのほうはかならずやconscious experienceを伴う?
Prediction errorは+と-の方向があって、salienceというのは+-極性無視して変化の大きさだけを評価している(元ネタ KatoTadafumiさんのツイート)。だからもともと両者はそんなに大きく違ったものではなくて、入力値をどう料理するかの違いでしかない。
Prediction errorは内部モデルの改変のために必要で、オフラインで使うためのもので、salienceはオンラインでの行動のトリガーとかに使うためのものでとか。
Salienceが極性気にしないということは、これはエネルギーの次元みたいな量を扱っているという可能性もある。そのことと、salienceがスピード重視で演算する必要があることとはなんか関係あるだろうか。
2012年09月13日
■ 連合学習理論の記述のレベルってどこだ?
(20020822あたりのツイートを元にして作成。)
いまブログの準備を兼ねて、澤幸祐 動物心理学研究2012(「連合学習理論は擬鼠主義の産物か -表現論としての連合理論-」pdf)を読んでいるところなんだけど、「中間言語としての連合学習理論」ってのになるほどと思った。でもって、そのレベルの文法体系それ自体が自立した整合的な(まさに文法的な)構造を持っているのかどうかという問題はさておき、これをデビット・マーの三段階の理論と(むりやり)つなげて考える。マー的にはImplementation-アルゴリズム-計算論という階層的な構造があって、あくまで工学的な問題解決から捉えているのだけれども、これの上に中間言語としての連合学習理論もマップできないだろうか?
たとえばそれはアルゴリズムの層であって(種差よりも共通性を重視する)、いっぽうでエソロジーというのは計算理論のレベル(=エコロジカルな意義のレベル、と読み替える)であると説明できないだろうか? ってこれは井口善生 動物心理学研究2008(「連合論的学習心理学と比較認知科学の断絶と接点」pdf)の方に関わる問題か。
このエッセイでは「比較認知科学に関わる者は、古典的エソロジーが最終的にメカニズム論を生理学に任せてしまったように、ヒトの認知心理学にメカニズム論を任せてしまうのか、それとも比較認知科学独自の予測性を持った理論屋モデルを必要としているのか、再検討してみるべきだろう」と書いている。
これはさっき書いた、そのレベルの文法体系それ自体が自立した整合的な(まさに文法的な)構造を持っているのかどうかという問題にもつながるだろう。
話を戻すけど、以前ブログに書いた話で、 マーの三段階の階層構造は分断されたものではなくて、ベイトソン的に表象とプロセスの連関でもって繋がっていると考えた方がいいのではないかと思う。
ならば、それは脳と体と環境の相互関係が作るバイオロジカルなレベルでの記述(バイオロジカルセオリー=たとえばSTDPで強化しましたとかそういう記述)と心理学的理論の文法構造とは階層的に繋がっていて、両者が拘束し合う形になるはずだと思う。
神経科学の研究はこのような意味で心理学的な理論に拘束条件を与えていくというのがマーが想定したプロジェクトだろう。これはバイオロジカルセオリー(たとえば上丘のニューロンの活動の空間的平均でサッカードのベクターの向きが決まる)だけで充分ですか、って問いになる。
今平行して読んでいるBBS2009 "The propositional nature of human associative learning"(pdf)では、現象としての連合とメカニズムとしての連合を分けるべきで、メカニズムとしての連合は自動的無意識的ななlinkと認知的かつ命題的ななものとがあって、後者を強調している。
とりあえずこのへんまでは読み進めた。とりあえず自分なりの問題意識は見えてきたけど、まだ考えはまとまってない。
書いたもの見直して思ったけど、BBS2009でのlinkのようなものはほとんどimplementation(バイオロジー)のレベルを置き換えて言っているのに近いのだろうな。すると、アルゴリズムの層よりももっとimplementationの層に近いような気がする。というか両者を同一視しているのだな。
2012年07月06日
■ 「確率論的世界と動力学系的世界」との相克?
「相関はダメで、因果を証明しなければならない」っていう議論についてだけど、けっきょく精度の問題ではないかという気がしてきた。天文学では星一個取り除いたりすることが出来るわけではないので、因果の実験ではなくて、その予測の精度によって厳密科学となっている。
相関ってのもけっきょくr=0.7みたいな相関だから文句を言われるのであって、もし相関がr=0.99999とかだったらそれは予測をしているのとけっきょく同じで、そこまでいけば厳密科学になる。
たぶんそういうことをデコーディングとかやっている人は考えているんではないだろうか。r=0.7をr=0.9にするのは工学的問題なんだろうと思うのだけど、r=0.9999にするにはどうすればいいか。なんかむちゃくちゃ正確なforward modelを作らないとそこはムリな気がする。
ちがうちがう、確率論的モデルを考えている限り、r=0.99999になることはありえなくて、動力学的モデルが出てこないとたぶんそこまでは行けない。池上さんが言ってた「厳密な数学」ってのはそのレベルのことだろうと思う。
地震が自己組織化臨界現象なのだとしたら、M9が起きる比率/M8が起きる比率 = M10が起きる比率/M9が起きる比率 というかんじで1000年に一度と言わず、10000年に一度の現象が起こりうる。だから、想定外はいくらでも起こりうるのであって、想定外をなくせなんて無理なのでは?
そういう意味では「ブラックスワン」なんてものはなくて、量が質に転換しただけであって、「ブラックスワン」を何か実体のあるもののように考えることはちと疑似科学に入ってるんだろう。もしM10の地震が起こったとしてもそれをブラックスワンと呼ぶ必要はない。社会科学の複雑さゆえか?
地震予知の不可能性の話はまさにロバート・ゲラー氏が唱えていることを知った(Nature asia翻訳記事)。そして、これもまた「確率論的世界と動力学系的世界」との相克なのだということに思い当たった。
確率論的世界では「次の30年以内にXXが起きる確率」以上のことは言えない。 そして、この構図は脳の計測と制御でも同じだ。短期間の制御と予測が出来たとして、その先は確率論的世界のものではないのだろう。かといって動力学系的世界のものと言えるわけではないが。
「海馬スライスでカオスを制御する」ってNature論文(PDF)が出たときはすげーと思ったが、あんまそこから先が進んだようすでもない。でも、その方向こそが制御と予測という側面からはファンダメンタルなものであるように思う。ちと忘れないようにしておく。
「確率論的世界と動力学系的世界」ってのは至る所にあるのだろう。疫学で統計モデルを作って病気の様々な要因について研究するのと、病気の要因間の相互作用を同時に計測しながらその時間的発展を研究する。
ん?これは正しいたとえだろうか?今の場合は前者から後者への自然な発展が想像できる。
いやいや同じか。脳も同時計測をしながらその時間的発展を研究することが出来ればかなり動力学系的な研究になる。いまはまだ疫学的なのだな。もちろん、疫学をおとしめているわけではない。
なにがファンダメンタルなことかって考えたりすることがあるけど、やっぱ「複雑系の中でどうやって制御を行うか」なんじゃないだろうか。神経科学、医療、政治、教育、どれでもローカルに擾乱を加えることはできるだろうけど、それによって揺り戻しが起こって、それを「構造的問題」とか呼んだりする。
成熟し、均衡状態になったシステムではどこでもこういう揺り戻しが見られる。なんにもないところに新しい土地を作る喜びってのは、そこはまだ均衡状態になっていないから、デザインしたとおりに物事が動く、そういう制御可能性の喜びなんだろう。それは革命思想。これが通常科学と科学革命の差異。
2012年06月18日
■ マカクでTET-on/TET-off --- 伊佐研からNature出ました!
私が助教を務めている、生理学研究所の認知行動発達研究部門(伊佐研)からNature出ました!
Kinoshita et.al., Nature 2012 "Genetic dissection of the circuit for hand dexterity in primates"
詳しいことはプレスリリースを見てもらうとして、要は、
- マカクでTET-on/TET-offシステムを使った経路選択的な機能遮断に成功した。
- マカクで、解剖学的に同定されたある経路だけに発現するようにベクターを二重感染させてやる。
- 順行性のベクターを領域Aへ、逆行性のベクターを領域Bへ注入すると、領域Aから領域Bへ投射しているニューロンだけで二重感染が起こる。
- でもって、DOX(抗生物質)入りの水を飲ませてやると、その時期だけその二重感染した経路が遮断される。
- これによって、損傷実験や薬理学的抑制よりもより選択的かつ可逆的に、ある経路がある機能に関連しているかどうかを切れ味よく示してやることが出来る。
- 損傷実験では機能回復や機能代償の効果が無視できない(だから私は機能代償の方に主眼を置いている)。
- ムシモル注入などの薬理学的抑制実験ではムシモルがどのくらい広がったかといったコントロールが難しいし、そもそもどこに注入できたかをあとで確認するすべがない。
こういう系をはじめてマカクで実現したということが重要。原理的には解剖学的結合が分かっていればどこにでも応用できる。マカクでこの方法が使えれば高次脳機能についてのこれまでの知見に応用が可能になる。よってそのインパクトは大きい。だからNatureに掲載された。
けれども、マカク用のベクターを開発する、といった各ステップがたいへんなので、このような仕事には大々的なコラボレーションが必要で、今回のこの仕事は脳プロ課題Cの枠組みを最大限に活用することによって実現した。そのへんに関しては文科省ライフサイエンス課の資料(PDF)が詳しい。
方法論の論文じゃんってのはその通り。今回の実験系は脊髄にチャンバーつけて行動中にムシモル入れるとかしないかぎりこれまでの方法では難しいし、ゆえに急性実験のレベルまででしか明らかになっていなかったことを明らかにしたという意味では、この方法ならではとは言える。しかし、ムシモル注入ではなくて、原理的にこの方法でないと見つけられないものを出せるかどうかというのがおそらくは次の課題となるのだろう。たとえば…まあ思いつくでしょ?
ともあれ、木下さん、おめでとう!(ラボの部屋の私の席の真後ろにいる。)
# やまだ
おめでとうございます。読売オンラインで知って、武井さんや大矢さんと盛り上がってました。すごいですね~Nature。
この方法はムシモルと比べてかなり画期的ですよね。ある二つの領域を繋いでいる細胞だけ機能を可逆的に落とせるんですから。
アポトーシスがどの程度起こるのか知りませんが、ほとんど完全に可逆的そうですね。この方法なら、あるネットワークの一部の経路を落とした時のネットワークの状態とか議論できますよね。最先端を切り開いていってるのは本当に尊敬します。意思決定のような曖昧さの高い研究にこそ、いろいろ使えるように思います。
どうもありがとうございます! まあわたしはオーサーではありませんが、このテクニックが今後いろんなところで使われていくといいなと思います。
どのくらい機能補償が起こるのかとか(図を見るとDOX飲ませているあいだにも成績は回復してる)、感染効率どのくらいだと認知的課題に影響及ぼせるかとか、いろんな場面でこれから検証されていくと思います。
2012年06月12日
■ 駒場広域システム講義の準備中。
6/20に駒場広域システムの学部講義(たぶんこれ:61066 システム科学特別講義II)で「意識と注意の脳内メカニズム」と題して講義します。池上さんから依頼を受けて、いいですね!ありがたく引き受けさせていただきます!なんて返答をしたら、90分 * 2コマ連続であることが判明。泣きそう。だがベストを尽くそう。そんなわけで、いろいろアイデア練ってた。
DFさんはtextureとか質感とかは関知できる。Humphrey et al 1994では懐中電灯を見せたときの例(レクチャーのPDFのp.25)がある:「台所用品。赤いパーツが付いてる。赤いところはプラスチックで他は金属。」手渡されると「懐中電灯か」
盲視ではこのような質感はない。だから、同じように腹側経路が損傷しているとはいえ、両者の視覚経験はまったく違っている。V1こそがそのような基礎的な視覚経験に必須であると言えるし、これを「感覚」と「知覚」の区別で言えば、sensation without perceptionと言っていいのかもしれない。
メロポンの入門書を読んでいたら、視覚はゲシュタルト的構成を元に一挙に与えられるのであって、知覚の前の感覚のような段階説は間違っているとするような書き方があって、どういう文脈で言ってるか分からないが、(メロポン的にはセンスデータ説批判ではなくて「行動の構造」以来の、ゲシュタルト心理学の含意の敷延のはずだから)、本人の文章ではどういう言い方をしているのか見てみることにしよう。
ニコラス・ハンフリーはトーマス・リードを引いて、このような感覚と知覚の違いに基づいて議論を進めるのだが、これは哲学者にはとても受けが悪いとこぼす(「赤を見る」)。たぶんこのときはセンスデータ説批判のほうから来ているのだろう。わたしも盲視から発想するので同じような考えに至る。
つまり、sensorimotor contingencyによって決まるような技能としての視覚(背側経路)とpredictive codingしてsurpriseをtop-downのawarenessによって消してく、ヘルムホルツ的視覚(腹側経路)との折衷、ってアイデアになる。
じつはこのようなアイデアはJoel Norman のBBS2002にあって、両者の範囲を正しく限定するという意味でよいと思うのだけど(Noeがcolorについてsensorimotoroの議論を応用しようとかするのは無理だろとか思う)、BBS2002自体の反応見てるとイマイチ。
なにより肝心のGoodale & Milnerが出てこないもんだから、Normanの話の前にGoodale & Milner説自体の妥当性とかの話になったりして。David Ingle (retired)がコメントしてたので期待して読んでみたら、昔話に終始して、使えない奴だった。
まだ全部読んでるわけではないけど、どうやらギブソン的視覚観とマー的視覚観とを統合したい、なんて動機がそもそも共有されていないんではないか、という印象を抱いた。
Goodale & Milnerの中でいちばんきっつい主張(dorsalはunconscious)にも与しない。腹側系は意識のcontentであって、それが配置され、他者と環境を含めた世界として経験されるためには背側系が必要。
進化の過程では、背側系の方が先立つと考えた方がよいのではないだろうか? つまり、Goodale & Milnerにハンフリー的な進化の視点を導入する。視覚への応答がvisuomotor processingそのものであった状態(背側系)から、表象の世界(腹側系)がどうできるか。
こんなことを今度の講義のまとめに持ってくるつもり。Goodale and Milner成分をいくつか付加して、通りいっぺんな説明ではなくてそれなりに血の通った話をして(DFさんの「視覚経験」)、盲視の話への導入とする。ついでにJCでも再利用。
前半は「注意」。サリエンシーマップと半側空間無視の話をして、前者ではpredictive codingまで、後者では空間と身体との関係まで言及する。これが後半の伏線になる。
後半は「意識」。両眼視野闘争とNCCとGoodale & Milnerの話をして、盲視を最後に持ってくる。盲視では質感はないけどサリエンシーはあるのだ、という話をする。脳とかSDTとかテクニカルな話をするか、それとも外在論とかenactionとかの話をするかのバランスを考える。
つまり、ニコラス・ハンフリーの話で出てくる原始的生物の話は、背側系(手で物体を操作し、目で定位する)という過程が先立って、その生態学的な拘束条件によって決まるアフォーダンスそのもの(たとえば手に届くものを届かないもの)が弁別の材料となる。
そのような弁別能力が長期記憶となり、カテゴリー化の源となる、といった腹側系の機能が出来る。このような表象自体が独り立ちして表象間で操作を行うようになると前頭葉が必要になる。ってこういうおとぎ話をえんえんと書く必要はないのだけど、アフォーダンスが表象に先立つ、というのはVarela-Noe系列のenactive viewとしても筋が通っていると思うし、enactive viewの適応範囲を正しく決めるのにも寄与しているんではないだろうか?
「その生態学的な拘束条件によって決まるアフォーダンスそのものが弁別の材料となる。」つまり、この時点では弁別そのものをしているのではなくて、行動として本当に手が届くか届かないかという事実だけがある。そこから行動しなくてもあれは届かない、という判断が出来ればこれは弁別したことになる。
つまり、行動をせずに、あれは届かないと判断するのが弁別であって、弁別は経験からの学習を前提としている。ってそりゃあたりまえだった。Perceptual decisionではこれがもっと具体的に確率密度分布で持つのか、それとも判断基準で持つのかとかそういう問題になったり。
OBEで「痛み」はどちらの「自己」に帰属するのだろうか? たぶん答えがあるはず。調べておこう。どちらに帰属するにせよ、それによって痛みを他人事にしてしまうことはできないのだろうか?
ksk_S @pooneil RHIでラバーハンドの方に痛みを感じるというのはあるようですね。素朴には、痛みのような内受容性の感覚はそれを感じてるところが「こちら側」になって、他人事にならないような気がしますが。
@ksk_S なるほど、rubber hand illusionのほうで考えればよいのですね。まさに「痛みのような内受容性の感覚」と視覚のような外界に投射する感覚とではいったい何が違い、どこに限界があるのか、みたいなことを考えてました。ではまた。
ksk_S @pooneil まさにそれについて僕も考えていました。RHIやOBEで問題にしている身体的自己感覚は外受容性なんですよね。内受容性の感覚は、身体のように帰属させる自己じゃなくて、もっと意識体験のフレームそのものに直接関与してるような気がします。
(4/21のを吉田がリツイート) ksk_S あともう一つ最近の疑問。形式システムと、力学系と、確率論的世界の上下関係。力学系は形式システムを内包してそうだけど、確率の世界は可能性を扱えるので力学系を含んでいるといえるのか? 含んでるけど目が粗くて捉えられないものがあるということなのか?
講義スライド用に今まで持っているマテリアルを並べてみたら、209枚になった。セクション用の見出しとかもあるから実質180枚。これだけあれば3時間の講義には充分だろう。どちらかというと、これを使ってちゃんとストーリーが流れるように構成することに注力するのがよさそうだ。
ブログ更新: 「脳の生物学的理論」からの話の展開: 20111227のtwitterでの池上さんと藤井さんとのやりとり。 pooneilの脳科学論文コメント 20120516
alltbl @pooneil ちなみに吉田さんは、脳や意識についての論文をかなりきちんとフォローされてると思うのですが、脳はどういうシステムだと思ってますか?Alan Turingの考えたチューリングマシーン的なものではないでしょう?
@alltbl むつかしいこと聞きますね。脳を実際に見ているものとして、脳はコネクショニズム的な分散表現を行っているというのが前提なので、古典計算主義的な脳観は持たない。ただし、そしたらニューロンの活動はニューラルネットの中間層みたいなことやっているのかというとそんなことはなくて、じつはスパース表現がなされていることが多い。つまり、おばあさん細胞のようなニューロン活動というものは偶然に出来ているのではなくて、どっかのレベルで最適化の結果であるらしい。そうなってくると、脳で表象をするということがまた違って見えてくる。
@alltbl あくまで仮説ですが、分散表象とかポピュレーションコーディングのような表象が背側系で行動を引き起こすのに使われて、腹側系でのスパース表現というのは表象の操作を含むような認知活動に関わっているかもしれない、とか考えます。
alltbl @pooneil なるほど。コーディングのような表象が背側系で行動を引き起こすのに使われて、腹側系でのスパース表現というのは表象の操作を含むような認知活動に、というのは面白いですね。ただ聞きたかったのは、何をしているかという時に、世界を写しとるというコピーマシーンみたいなもの?
@alltbl ちょっと寄り道しましたが、このようなニューロン活動のあり方というのが、先日の鈴木さんのツイートにもあったような、「形式システム」と「力学系」と「確率論的世界」のすべてに対して寄与しているんではないだろうか、とか考えたりします。
@alltbl ニューロン活動がポピュレーションコーディングで確率論的な振る舞いをすると同時に、スパース表現でばらつきのない確実なニューロン間通信を行う、みたいに考えたら、確率論的な脳と力学系としての脳が同時に説明できないかなとか考えました。
@alltbl 強い表象主義だと外界のコピーを内的に表象することになるけど、それは無いと思う。まず、背側系は技能として視覚を使うのでコピーをしない(昨日書いた、enactiveな脳)。腹側系は外界をinferする表象を作成するけど(昨日書いた、ヘルムホルツ的脳観)
@alltbl 、実のところ注意を向けたところしかinferしてない。これこそがchange blindnessからわかったことで、われわれは注意を向けていない部分についてはコピーを作っていない。(これはpredictive codingの観点から説明するのが良いと思う)
alltbl @pooneil コピーマシーンなんだけど、自分で世界を変えてコピーしやすくしようとする? 必要以上に脳の仕組みが複雑に見えるので、他に何かしてるんじゃないかと。
@alltbl うーん、これは池上さんの言葉が分からない。
alltbl @pooneil すいません。運河を見てましたw Andy の読みましたが、どうなんだろう。ぼくはこのpredictive codingに賛同できないですね。というのも、生命は予測を最適化するならば、暗い部屋にじっとしてるはずだけどそうではないし、遊びこそが大事、だと。
@alltbl predictive coding的にいうなら、コピーを作るんではなくて、予想外だったときのサプライズを脳内に表象を作ることでキャンセルアウトする、というかんじで。(Andy Clarkもなんかこのへんについて言っているけど、まだ読んでない)
@pooneil これまではミクロには力学系で、疎視化すると確率論、とか考えてたけど、こういう可能性もないかという思いつき。
@alltbl predictive codingにしろ、ベイズ脳にしろ、最適化と言いつつ最適化しようのないノイズというか揺らぎがたくさんあるのに抗しているという状態なのだから、最適化と相反する作用とのバランスという図式を描かないと、池上さんの言うとおりになると思います。
predictive codingだと最適化した行動を前提としているとかいうのはニューロンレベルと行動レベルとのカテゴリー錯誤がありそう。predictive codingの重要度はニューロンの表象の意味を一変するところにあり、おばあさん細胞はおばあさんを表象しているのではなくて誰もいないというpriorからおばあさんが現れたサプライズがニューロンの発火として表現されて、それが緩和される過程を我々は観察者としてみているだけだし、脳内では、上流の細胞が下流の細胞のサプライズを消すように活動することが結果として情報をデコードしてことになってるんだと思う。
- / ツイートする
- / 投稿日: 2012年06月12日
- / カテゴリー: [オートポイエーシス、神経現象学、エナクティヴィズム] [大学院講義「意識の神経科学」] [脳科学メモ] [腹側視覚路と背側視覚路] [視覚的意識 (visual awareness)]
- / Edit(管理者用)
2012年05月29日
■ 表象、光点、ベイトソン、オートポイエーシス
TMSや微小電気刺激によるphospheneはたしかに視覚なのだろう。でもそれは知覚そのものを作っていると言うよりは、すでに視野が用意されていて、そのcontentを書き換えてやったと考えた方がよいのではないかという気がしてきた。
盲視の人は、明らかにMTは活動しているのだろうけど、内観を聞いた感じどうみても視覚という感じはしない。アルヴァノエの本で似たような議論があったときには釈然としなかったけど、知覚のcontentの表象とか操作とかよりは、視野という場そのものを作ることの方を考える方が意識研究なんではないかという気がしてきた。
まだ充分飲み込めた気はしない。でも、表象がsurpriseでありprediction errorであるという構図を進めていくこと以外の道を探るとしたら、こういうことを考えるしかない。でもってこれってたぶん、現象学だな。どうすればよいんだ。
ベイトソンの問題圏というのは徹頭徹尾表象の世界なのだな。違いを生み出す違い、メタになるプロセスと表象。一方でオートポイエーシスは根本的には内部の視点の話で、そこからカップリングとかを使って表象とかを作ろうとしているわけだが、それは成功しているだろうか。
外側から内側を推測しようとするのと、内側から外部を構成しようとするのと、両者は向きの違いの問題であって、どちらも同じくらい原理的にムリがあるのではないか。
オートポイエーシスで「カップリング」という装置を使って表象の世界と行き来しようとしたのと同じように、ベイトソンの「プロセスと表象との間でジグザグとレベルの上下する」という装置を使って表象と内部のダイナミクスとを行き来しようとした。少なくとも私はそのように読んだ。
反表象主義なんて極端なことを言う必要はないのだけれど、ミラーニューロンとかPPCニューロンの捉えにくさってのは「生態学的な意味づけを持った行動」を表象の方から捉えようとするがゆえなんだと思う。って自分で書いててもいまいちピンと来ない。「行動」をどう捉えるかなんだと思うのだけど。
2012年05月16日
■ 「脳の生物学的理論」からの話の展開
20111227のtwitterでの池上さんと藤井さんとのやりとり。
pooneil 因果的関係は物理的に見れば非常に細かいステップの連鎖なので、その連鎖の部分をはしょって、2つの変数間の関係を取り扱うのが相関。だから相関とはあいだにブラックボックスを置いたままで現象を扱おうとする簡便法であり、表象-プロセスの関係で言えば、表象だけでなんとかしようとする方法。
pooneil そう考えると「脳の生物学的理論」、つまり、計算論の層や認知科学的概念を入れずに「どこどこのニューロンが活動してそれによってある領域のニューロンが活動して、その結果行動が起こった」みたいに説明しようとする考えは、物理的な因果を追うことを志向していて、そんなナイーブなものでもない。
alltbl @pooneil おー、それってナイーブではないと思いますよ。その物理的因果の連鎖は難しすぎる。
alltbl @pooneil 難しすぎる、ってことなのですね。前の方のツイートをみると。むしろ「因果律」というフレームがつくれないって、ことですね。
pooneil @alltbl 神経生理学者は「生物学的理論」を志向していて、認知的モジュールを厳密に考えるのは内心不毛だと思っているように思います。(たとえばこのニューロン活動は「注意」を表象しているのか「意図」を表象しているのかとか。) ある意味ニューロン中心主義なわけです。
pooneil @alltbl でも、生物学的理論を徹底すると因果の連鎖を本気で考えることになって、そうするとMarrのimpelementationの層を[表象-プロセス]まるごと捉えることが出来るようになる。ここまで来て初めて、アルゴリズムとか計算論の層とかまで考える意義があるかもしれない。
alltbl @pooneil なるほど。人工生命や複雑系は、「適度な疎視化」こそが、重要で、ぼくはそれを「中間層」とよんだけど、そこにこそ厳密な数学が成り立つと考えてます。
NaotakaFujii @alltbl @pooneil だからさ、理解するんじゃなくて、自分もそのネットワークの一部になるしか無いんだと思うんだよね。理解じゃなくて感じるしかないんだと思うな。世界の仕組みは分からなくても、世界の一部ではあるという感覚。
pooneil @NaotakaFujii 観察者ではなくって「内側から理解する」ためにどうすればいいかというと、たしかに藤井さんの言うやり方になるんではないかと思います。けっきょくそれは「説明」することではなくて、「了解」するってことだろうから。
alltbl @pooneil つまり、生物学的なニューロンをreplaceできるようなレイヤーの設定ということなんだけど。
alltbl @NaotakaFujii @pooneil うちのTom Froeseとかもその考えです。ぼくも、そう考えるけど、なんかまだその分かり方になじまない。
pooneil @alltbl それはコネクショニズムで言う中間層とかpopulation codingとかの概念とおなじなんでしょうか? ともあれ、個々のニューロン活動と個々の認知的概念や計算論があって、それを媒介する中間層を想定して、それの動力学を考えるというのは賛成です。
alltbl @pooneil うーんと、それとは違って、物理的にニューロンが下層にいなくてもいいことを積極的に証明するってことです。
pooneil @alltbl @NaotakaFujii 外側から説明しても内側から了解できてないじゃんってのと同じように、内側から了解できても外側から説明できてないじゃん、ってことが起こりえますかね? ともあれ内側からの方がもっと必要なのは確かだけど。
NaotakaFujii @alltbl @pooneil 外側からの理解に方法論的に絶望したから逃げてるだけのようにも思うけど、多様な了解可能性があるのが希望ではあります。
pooneil @alltbl なるほど、それは以前の話に繋がりますね。ニューロンという物理的実体がなくても心が作れるとしたらそれはニューロンそのものではなくてより抽象化したものが本質ってことですね。そこでの「厳密な数学」というのは情報処理の話だけでしょうか? もっと物理的なものでしょうか?
pooneil @NaotakaFujii @alltbl 「多様な了解可能性」ってところが面白いですね。「自転車に乗れるようになる」みたいな技能的なものと「赤い色とはどういうかんじがわかる」みたいな内観的なものとかがどう違っていてどう同じなのかとかそういう問題がもっと具体的になる気がします。
alltbl @pooneil いや、非常にタンジブルな感じとか、目の前にあるという「この感じ」が疑いえないもの、は、その意味で「厳密」なので。
alltbl @pooneil @NaotakaFujii コンピュータの内側のエージェントに、自分が内側だってことを了解させる謎のプロジェクトは、うまくいかなかったw
pooneil @alltbl @NaotakaFujii ありがとうございます。またそのうち今日のやりとりをブログにまとめてさせていただきます。
2012年04月23日
■ ブラックスワンと世界滅亡の夢
東北大通研でなんか気の利いたこと言えるように「心理学が描くリスクの世界」とブラックスワンのおまけ部分を単行本化した「強さと脆さ」を借りてきた。タレブの本は尊大すぎて嫌な気分になるので読み進めるのがつらい。
ヒトの不合理な楽観的思考 これ面白い。ガンになる確率を推定してもらって、あとで実際の確率を提示される。そこをfMRIで見る。ガントリの騒音の中で現実を見せられて、なんだかどんどん気が滅入っていく被験者を想像するとなんだか可笑しい。
そういえばなるほど私は生命保険に入っているにも関わらず自分が死ぬ確率を知らない。生命表で調べた。第20回生命表(男) いま43歳の私が60歳になるまでに死ぬ確率は7.2%。(= 1 - l_60 / l_43) やっぱ生き延びるだけですごいことかも。
計算してみた。五分五分の確率であと40年生きるのか。無限? 揺らいだ 有限つまりは。
「ブラックスワンと世界滅亡の夢」ってのを考えた。太るまでエサを与えられた七面鳥がある日終了するのと同じようにいつか世界も滅亡するが、その確率を我々は計算することが出来ない。それでもわれわれは「めったにあり得ない現象」を心の底で恐れ、その恐怖は「世界滅亡の噂」として繰り返し現れる。
2012年04月10日
■ MacaqueでのDTIの現状
MacaqueでのDTIの現状を調べておく。ただし、DTIとかDWIとかDSIとかいろいろあってなんか違うんだろうけど、今のところフォローできてない。
以前調べたときはSchmahmann et.al. 2007 "Association fibre pathways of the brain: parallel observations from diffusion spectrum imaging and autoradiography."でpost-mortemで25時間スキャンって書いてあって、そんな長時間動かしてスキャナーぶっ壊れないのかとか思った。
[Methods. 2010] "Optimization of in vivo high-resolution DTI of non-human primates on a 3T human scanner." 3Tでin vivoでやってて、1mm voxelで36-72minかかる。[Neuroimage. 2012] "A diffusion tensor brain template for rhesus macaques." こっちはぱっと見、取得にかかる時間は書いてない。
Humanのpost-mortem [Neuroimage. 2009] "High resolution diffusion-weighted imaging in fixed human brain using diffusion-weighted steady state free precession."のほうはどうかと見てみれば、50hだと。[Neuroimage. 2011] "Diffusion imaging of whole, post-mortem human brains on a clinical MRI scanner." こっちは100h。そういうものなのね。
同じ著者の[Magn Reson Imaging. 2009] "A practical approach to in vivo high-resolution diffusion tensor imaging of rhesus monkeys on a 3-T human scanner."
あとは読むだけ(<-PDF取っただけで満足するタイプ)
Monkey to human comparative anatomy of the frontal lobe association tract (PDF) ヒトとマカクで白質走行の相同をまとめたもの。AFとかSLFIIIとかの妥当性が気になる。
arcuate fasciculus(AF)は左脳ではウェルニッケとブローカを繋ぐところで、これの損傷で失語症が起こる。右だとこれが半側空間無視のventral networkだとたぶんHO-Karnathとかは考えているんだろう。Ventral pathwayはけっこう穴だ。
Nature Neuroscience 2008 The evolution of the arcuate fasciculus revealed with comparative DTI この論文だと、AFはマカクやチンプでは未発達なので、これがヒトの言語発達と関連してるって話になる。
2012年03月29日
■ 超解像とマッピング
複数フレームからの超解像画像みたいな話題の場合はわざと解像度を下げた画像を作ってそこから元画像を再現するみたいな例がよくある。そういうばあい、samplingがせいぜい2倍とか4倍とかそういうのしか見ない。もっとセンサーの数が少ない例はないのだろうか。
たとえば、4*4のセンサでちょっとずつずらしながら128*128の画像を再構成するような。っていうかそれだったら一個のセンサーをスキャンして128*128の画像を作ればよいのか。 要は電極でニューロンの記録するという状況でなんか超解像的なことが応用できないのかって話なんだけど。
テトロードってのは三角測量みたいなもんで、一個のセンサだとシグナルの大きさとシグナルからの距離が分離できないから、複数のセンサを使ってそれを分離するということに原理的にはなるのだけれども、テトロードの場合、センサの配列の仕方に制約があるから完全にそれが出来ているわけではない。そういうわけで、これはべつもん。
佐藤先生のforward modelを作ってVBでセンサの数よりももっと詳しく推定をするってのと同じ問題。でも電極は動かせないので、超解像みたいなことをそのまま応用することは出来ない。
一個のセンサーをスキャンして128*128の画像を作ればよい<< これかならずしも正しくなかった。そんな狭い部分を記録できるセンサがあればよいが、もし、受光する部分が広いとしたら、それを使ってどのくらい細かいものを再構成できるか。
具体的な問いとしては、128*128の元画像から4*4のモザイク画像を様々なshift(とりあえず平行移動だけ)で作ってやって、そいつらからどのくらい128*128の元画像が再現できるかをやってみればよいのだな。
layer 2-3はoptical imagingのほうがほとんどすべての意味で勝っているからそちらのことを考えるよりかは、layer 5のprojection neuronの活動の空間分布みたいなことを電極アレイで解決できないか考える。てかもう超解像関係ないし。
一般的なシリコンプローブみたいな直線的にコンタクトが配列しているものでもなく、剣山電極みたいな間隔が開きすぎているものではなくて、ある立方体の中で3D方向すべてで50um間隔でコンタクトが配置されているようなものから記録することが出来れば、本気でポアソン方程式とか使えるのだな。
なんかlayer 5あたりに電位感受性細胞をinjectionしておくと、そいつらが他のニューロンとはシナプスを作らずに脳表までprocessを延ばしてくれて、その部分をoptical imagingする。うーむ、SFすぎるか。
SFついでに考えると、そもそも細胞をinjectionする必要ないな。いまあるニューロンを電位感受性細胞として使えばよい。なんか感染させてやって、そのニューロンの出力とシナプス入力を停めてやって、そいつらの軸索側枝が光学測定できるくらいの浅さまで伸びてくれればよい。
なんか自分で言っててわけわからなくなってきたぞ。つまり、ニューロンはそれじたい前シナプスニューロン活動のセンサなのであって、べつに出力停める必要もない。シナプスの入力は全くのランダムというわけではないので空間的な統計があまり使えない。
だから電位感受性センサとしてだけ使うとしたら、dendrite全体にチャネルが分類してしまっていると再構成しなくてはいけなくなるので、細胞体のチャネルだけで、しかも超局所的ではない細胞外電位が記録できるようなことが可能であればよい? そうするとチャネルのコンダクタンス的には既存のものでは向かない?
あとで見たら何言っているか分けわからない。だが残しておく。
2012年03月22日
■ ヒトの脳にはニューロンがどのくらいあるか
ヒトの脳にニューロンがどのくらいあるかちゃんと測った論文がAzevedo et.al. 2009で、860億個だそうな。体積の80%を占める大脳皮質に19%のニューロンがあって、体積の10%を占める小脳に80%のニューロンがある。なにげに小脳すごい。この事実は小脳の重要さを強調するために引用されたりする。たとえばRoger LemonのLife without a cerebellum。
小脳の説明をするのに、大脳はCPUで、小脳はGPUで並列計算するのに向くアーキテクチュアになっていて、わざわざM1から長い線維伸ばして計算させてまた戻すとかコスト払っているので、大脳皮質の構造では出来ないようなことをやってんだろう、とか説明をしたりする。ほんとうのところはわからない。小脳に聞いてください。
Azevedo et.al. 2009でのもう一つのポイントは、ニューロン-グリアの比率は1:101:1くらいで(3/26訂正。Thanks to Gayaさん)、これはほかの霊長類でも同じということ。そういう意味ではヒトの脳はマカクとかと比べてとくに特別なわけではない。
The human brain in numbers: a linearly scaled-up primate brain 同じ著者が書いたレビュー。これの図3がよくて、いろんな齧歯類、霊長類の脳の重さとニューロン数が一覧できる。
{kind=link}
それによると、マウスは脳重が0.4gでニューロン数が7千万、ラットは1.8gで2億、マーモは7.8gで6.3億、マカクは87gで63億、ヒトは1500gで860億。こういうデータを持っておくと講義とかそういうときに捗るんでオススメ。
追記3/26: ニューロン:グリア比が1:1ってのはけっこう重要な話で、上述のFrontier誌を読んだところ、これまで信じられていた1:10というのはほとんど根拠がないうえに、神経神話「人間は脳の10%しか使えていない」の元となっている可能性があると。グリア比が多いことが分かっているのは視床だったり、ventral pallidumだったりというところでの報告だけで、大脳皮質ではグリア比が2を超えることはない。いちばんニューロン数の多い小脳では逆にニューロンの方が多くて、ニューロン:グリアの比は>=25:1 (at leastって書いてある)。 以上の数字はHerculano-Houzel本人の仕事ではなくて、それまでの文献から得られたものなので、カウント方法にはそんなに依存しなさそう。
# Gaya
あれ?この論文では「ニューロンとグリアの比率は、かつては1:10だと言われたこともあったが、実際には1:1だった」と主張していますよ。それがメインの発見だからこそ、論文のタイトルにもなっているのかと。
# pooneilあ、ほんとだ。斜め読みで完全に誤読してました。もうちょっと読んでから直しておきます。ご指摘ありがとうございました。
2012年03月09日
■ 「お話としての説明」と「科学としての説明」という対比
Per BakのPNAS1995 Complexity, contingency, and criticalityの初めのほうを読んでた。History vs Scienceという対比があってなるほどと思った。
History(=進化とかそういった一回性の現象)にはnarrative accountつまり物語的なイベントの連鎖としての説明が行われる。ここでは砂山モデルで戯画的に説明されているが、砂山のシミュレーションで大きな雪崩が起きるとき、それはnarrative accountでは、「いくつかの悪い偶然が重なって予想外に大きい雪崩が起きた」といった説明がされる。
しかし物理学的説明からすれば「それはSOCであって、そのような大きな雪崩はpower lawからすればなにか特別なところがあるわけではない」ということになる。(因果的な説明ではなくて、統計的な描写でしかないところに注意)
ここでもhindsight (後付け)という表現が使われているが、けっきょくブラックスワンの特徴として挙げられる「あとから後付けでいくらでも説明できてしまう」という部分はたとえ科学的な言葉を使った説明だったとしてもnarrative accountでしかないのだということが分かる。
おそらく認知神経科学のほとんどの部分はそういったnarrative accountでしかないのだろう。たとえば脳のある部分が損傷したから、XXの処理が出来なくて、その結果YYの課題の成績が低下する。しかし様々な要因(脳内及び行動面での代償とか)があって予想通りとはならない。
だから、論理の連鎖をあんま長く続けることは避けた方がよい。もしくは関係ありそうなパラメータを全部突っ込んでシステムバイオロジー的に行うか。
前者の方策は問題をなるたけ細分化するかまたは禁欲的に操作的に行うという方策。たぶん行動分析というのもなるたけ説明変数を減らそうという方策で前者の範疇に入るのだろう。前者はどこか「禁欲的」な側面を持つ。
意識とかエピソード記憶といった一回性の事象を科学的に説明するというときに、事象の方だけじゃなくて、説明の方にもnarrative accountと物理的説明という、違ったものがあることを考えることは重要だ。narrative accountに頼らないのなら、脳と身体と環境との物理的因果の動力学を描写することこそが一回性の事象の説明となる。
あともうひとつ面白いと思ったのは、恐竜の絶滅も様々なスケールの絶滅がある中で考えれば確率的に起こりうるものだという話で、なるほど、なんだかものすごく特別なイベントのように思えたけど、これもpower lawでいいじゃん(本文中ではこういう書き方はしてないが)とけっこう得心した。
まあこうやって、揺れながら、人混みをうまくすり抜けるつもりで左右に揺れ動きながら、かえって人にぶつかってしまって遅くなるような(じっさいよくあるんだこれが)、無駄な動きをたくさんするのがどうやら私の行動原理らしくて、たぶんなんかメタなパラメータを最適化しているつもりなんだろう。
sandpile modelをmatlabで書いてみたくなったが、禁欲してコードを探すところまでにしておく。
- sandpile (matlabcentral)
- Pattern Formation in Growing Sandpiles (pdf)
- Gradient Based Bak-Tang-Wiesenfeld Sandpile Model (pdf)
2012年02月26日
■ サルには利き腕ってあるの?
調べものをしてたらいつのまにか「マカクの利き腕」あたりの話題にぶつかったのでまとめておく。というかものすごくたくさんの研究があるので焦点を絞るのが難しい。ニホンザルに関しては幸島のサルで左17、右8、あいまい16という報告("On the handedness of Japanese monkeys" 徳田喜三郎著)がある。
べつの論文("Preferred hand use in the japanese macaque troop, arashiyama-R, during visually guided reaching for food pellets" 久保田競著)でもおなじかんじ。このへんのは、餌をやったときにどっちの手で取るかといったような調べ方をしている。学習効果を見るために、暗くして触覚弁別課題とかやったりすると左優位なのが減る。"Lateral Preferences in Monkeys"
とかやってるときりがないので、Primate laterality - Google ブックス この本の6章を読むと歴史的経緯からいろいろ書いてあるようだ。ただし、これは1993年の本。
そのあとでプログレスと考えられるのは、bimanual taskの使用をしているものがある。これではマカクは右優位だそうだが、同じ著者がinfantでは左優位 と書いてるんでいまいち信用できない。
メタアナリシスしているのもある。 Old world monkeyでは左優位ということになるらしい。
とはいえ、けっきょくのところstandardized testが必要だなと思うわけで、これはbaboonの論文だけど、ヒトのテストを応用して、trunkの向きとかそのへんまで考慮している。
予想以上に込み入った話だった。しかもこれはまだ脳機能イメージングによる脳活動のlateralizationの話を入れていない。
とりあえずメモっとく。
2011年10月21日
■ フィードバックとフィードフォワードにおける時間 (池上さんとのやりとりを含む)
ついったでいろいろ書いていたら、池上高志さんとのやりとりになって、考えていたことがまたもや確率論的世界と動力学系的世界の相克なのではないかなんてことに思い至った。池上さんの許可を得て、転載します。池上さんのツイートはalltblから始まるもの(リンク先はツイートのparmalink)。
つぎのJCで何を採りあげようか。いま考えているのはNature Neuroscience 2011の"Decoding the activity of neuronal populations in macaque primary visual cortex."で、Population codingによるprobabilistic codeというやつをちゃんと読んでおきたい。
話の流れ的には、Zemel et.al. Neural Comput. 1998でencoding-decodingの基本的なスキームがあって、Jazayeri M, Movshon JA. Nat Neurosci. 2006 でニューロンが独立でポアソン的ならば、Log likelihoodがチューニングカーブと発火頻度の掛け合わせになることを示した。
今回のNat NS 2011ではこのような単純なモデル(発火頻度を足し合わせるときの重みにチューニングカーブを使う)とニューロン間の相関を考慮に入れた場合の重み(重みをデータから学習させる)とを比較している。ぱっと見、correlationを考慮したときの重みがチューニングカーブと比べてどう変わるか、みたいな図がなかったので何が起こっているかよくわからん。
別のラインでさらにBayesian inference with probabilistic population codes [Nat Neurosci. 2006] とかも理解しておきたいが、そもそも「確率的コード」という概念自体が重要。おなじファーストのSignal detection theory, uncertainty, and Poisson-like population codes [Vision Res. 2010]も読んでおきたい。
著者が書いているとおり、SDTはpdfを使っているからこれも確率的コードの一種なわけで、そういう確率的表現をニューロン集団としてどのように持つことができるかということが、神経生理的には重要。単一ニューロンでuncertaintyをコードすることと確率的コードとの関係とか。
これは意志決定の問題でもあるし、メタ認知の問題でもある。単一ニューロン記録でuncertaintyやるのはKianiとかKepecsとかでもう行われたから、やるならニューロン集団での確率的コードみたいなことを考えるべき。でもRGCとかはもうデータがあるから、Pillow et.al. 2008みたいなかんじでどんどん進んでいくだろう。
高次の脳領域で、意識の問題と絡めてこういうことをやっていけるとしたら、私がやる意義はあるかもしれない。あんまShadlenの後追いみたいなことをやってもしょうがない。ただし、意識の問題をSDT側からいろいろ追っていたら、そういうところに突き当たったということだけど。(盲視でFC d' > YN d'となる問題)
確率表現の話も内部モデルの話も、私にとっての興味というか論点は同じで、単回でのフィードバック制御と、ヒストリーの分布を持った上でのフィードフォワード制御とは論理のレベルで別なのだというベイトソンの言い方を使って、なんかうまいこと整理できないかっていうこと。
どうも理系らしい言葉にならないけれども、フィードバック制御には過去がないから自己はない。フィードフォワード制御には過去があって、内部表現があるから一段高次。これが「なぜサーモスタットには自己がないと言えるか」の答えだと思う。
alltbl @pooneil すいません、FFとFBの解釈が、わからない。Recurrentは?
確率表現というやつもそういう意味で、現在のみに生きていない有機体でしか起こらないこと。だから尤度がなんか本質的なものなんだろうと思う。ベイズかどうかはpriorがあるかどうかだけの違いではあるけれども、渡辺澄夫氏の書いたものを読むかぎり、尤度-priorという対で一つのモデルということらしい。こういうことをもっともっと考えたい。たとえそれが望まれていなかったとしても。
石ころ単体に意識はない。意識があるとしたらそれはなんらか相互作用が必須で、石ころはほかの物体となんらか相互作用をして情報伝達をすることによってはじめて意識システムの一部となりうる。
alltbl @pooneil 電子1個の意識、というやつですか?
といいつつ、「情報」という言葉を不用意に使ってしまった。情報は読み、読まれるという意味で観察者が必要。内部的にはたんに物理法則に従っているだけなわけで。
@alltbl recurrentはFBではないですか。ただし、時間遅れがある。今の文脈で行くと、では時間遅れがあったら過去があると言えるか。無いんではないでしょうか。
alltbl @pooneil ぼくは、FBにはかならずtime delay があるし、FFは記憶はないと思いますが。
@alltbl 「電子1個の意識」ってことを言っている人がいるのですか? 今書いているのとはちょうど逆方向かと思いますが。
alltbl @pooneil そちらの文脈だと、電子1個の意識もあることになり、それと石の差は、かなり曖昧になる。
@alltbl いまの文脈からすると、電子一個でもほかのものと情報のやりとりをしているならば意識システムの一部となると言えるかもしれない。ただ、そうなると本質は電子や石ころではなくて、情報の方になる。すると「情報」自体をちゃんと考えないとこの話は意味がなくなる。
alltbl @pooneil ぜひ、研究室の Life Mind Seminar で話してください。歓迎します!
@alltbl 確率表現みたいな話と意識のオートポイエーシスの話がうまくつながったらぜひお話をしてみたいですが時期尚早かと。それとはべつにして一度Life Mind Seminar聞きに行かせてもらえませんか? 日時とか情報教えてください。
alltbl @pooneil 情報というのは、時間的な不可逆性が生まれること = 観測問題、では?
@alltbl なるほど、情報という概念を使うと観測問題になっちゃうのはわかっていたのですが、時間、過去という概念も使ったとたんにもういろんなものを先取りして入れてしまうことになるのですね。
alltbl @pooneil はい、不定期なのですが、是非。確率表現というのは、外からの視点で、Autopoiesisは、内部的な視点。ですよね? なぜなら、外から見たら、死ぬ確率が1/3という場合でも、本人にとっては0か1だから、つねに。
@alltbl なるほど、もっと精緻にして考える価値がありそうです。直接時間を扱っているのはFBで、FFは実行時は学習結果の重みを演算しているだけという意味では過去を直接利用しているわけではない。つまり、両者は時間への関わり方が違っている。
わたしが書いたのはFBはそのつど誤差を計算すればよいという意味で過去の蓄積を必要としない。FFは(実行時ではなくて)学習の段階で過去のヒストリーによって重み付けを変えるという意味で過去の蓄積を必要とする。ベイトソン本の例では、ライフルで照準を見ながら正しく標的を合わせるのがFBでライフルで複数撃った結果を元にライフルの癖とかを学習して狙うようになるのがFF。
alltbl @pooneil うーん、でも、いままさに生きている時間、というやつがなくなってしまうのでは? 時間は生きているから。
@alltbl ですよね、確率という概念は外からの視点ですよね。それで、この問題は本当は確率的表現と力学系との関係の話になる。津田先生の新学術に入れていただいていろいろ発表を聞いてきたのですが、神経科学で力学系ってどうやったらよいのだろうともやもやしていたのとつながってきました。
@alltbl 推測ですが、FBの時間遅れの方が、現在と重なりながら進んでいく、生きられた時間に近いものと捉えていたということでしょうか? そういう意味ならば納得がいきます。FBの時間は力学系的な時間で、FFの時間は確率表現の世界の時間、なんてまとめ方はどうでしょうか?
alltbl @pooneil FFはやっぱり力学系を走らせてるだけ、という感じですよね。FBは、おっしゃるとおりの解釈です。
@alltbl ありがとうございます。やりとり面白かったんであとでブログでまとめ直してみようと思います。転載させてください。お願いします。
alltbl @pooneil はい、光栄です。こういう話はtwitterだとむずかしいですね。はじめてそう思った。
池上さんとのやりとりを元に今日書いたことを見直してみる。内部モデル的なアイデアはMLE的、ベイズ的ひっくるめて確率表現の世界を前提としているようだ。となると力学系でのカオス的遍歴としての記憶というものが内部モデルのようなアイデアとものすごく離れていることがよくわかる。
これは本質的なことかどうかはわからないけれども、確率的表現の世界と力学系の世界とで時間の見え方が違って見えるだけでなく、お互いに隠蔽しあうような形になっているかもしれない。ある意味相補的なのかもしれないし、物事の捉え方が反転していることを反映しているのかもしれない。
@alltbl ありがとうございます。むずかしいですけど、twitterならではのやりとりだった気もします。Face-to-faceのスピード感だとこういう展開にはならなさそうで。
alltbl @pooneil なるほど。それはそうかな。twitterは研究の議論にも向いていますか? ちなみに僕の疑問は、ソフトウェア、ハードウェアという線引きは、どの程度、脳では明確だと思いますか?
@alltbl twitterで研究の議論するのは難しいですね。どうしても断片的になりますので、clarificationに終始してしまうでしょうし。詳細を突き詰めるよりはインスピレーションを得る方が実りがあるし、そういう意味では池上さんとのやりとりは楽しいです。
@alltbl 「ソフトウェア、ハードウェアという線引き」これの論点はいろいろありすぎてぱっとはわかりませんが、たとえば、あるニューロンの機能を決定づけるのは他のニューロンだという意味で関係主義的に捉えた方がよいというのは神経生理学的立場からも言えるのではないかと。
alltbl @pooneil コンピュータのメタファで脳が考えられるか、という意味です。TeXやCromeなどのアプリは、下のハードウェアを考えなくても乗せられるか、という問題が、脳にはあるか、ということです。
見直して考えるに「サーモスタットには自己がない」をもっとちゃんと言い直すと「サーモスタットには表象がない」となるだろう。つまり内部モデルとは表象のことであって、観察者を前提とする。「サーモスタットに生きられた時間がある」かどうかは観察者ではなくてサーモスタット自身の問題だ。
@alltbl それは「情報」の概念とかかわりますね。もし情報のやりとりだけが重要なのであったら、それの乗り物は脳である必然性はない。V1ではなくてA1が視覚を処理できるように、可塑性を使ってぜんぜん別のニューロンに機能を移し替えることはあるレベルまでは可能。
alltbl @pooneil そうすると、認知心理学的な現象は、ニューロンとかじゃなくても実装可能。だと思いますか? ロボットということだけれど。だとすると、逆に脳科学は、神経細胞で生じているから偉い、ということに?
ではこれはニューロン以外でも可能か。わたしは外在論的立場に親和性が高いので、環境との相互作用のループを保ちながら徐々に脳以外のものに機能?を移行してゆくことは可能だろうと予想して、BMI的研究がそれに寄与できるのではないかと考えてます。
@alltbl 機能は実装可能として、現象的側面が実装可能かどうかってのが論点かと。「実装」って言葉にかかわる問題ですが、オートポイエーシスは分岐させることは出来ても、丸コピーは出来ないのではないでしょうか。コンテキストがコピーできないから。機能と現象の違いはそこではないかと。
alltbl @pooneil 難しいこといいますね。個性はコピーできないけれど、新しい個性は作られるのでは? イーガンの黒い宝石、でしたっけ?それみたいだな。
@alltbl コピーは出来ないけど、新たにはじめることは出来る。池上さんがオートマタ動かしたり、油滴実験したりするのはまさに新たにはじめることが出来るからですね。となると論点は、コンテキストがないところではじめられたオートポイエーシスは新たな個性なのかそれとも生きてない偽物か。
alltbl @pooneil 生きていけたら、本物ですね。
ここまで書いて気づいたけど、私の書く「コンテキスト」にはちょっと生気論的な神秘が入っているようだ。オートポイエーシス的に言えば、維持されるかぎりは新たに生み出されたものでもなにかが生きていると捉えるのが正しいか。
伊庭幸人氏の「学習と階層」から:「階層、とくにあらかじめシステムに作り込まれた階層は、非線形動力学の陣営からは、旧弊な機構として厳しい批判をうけている(Kaneko and Tsuda(1994)) ..."表現とアルゴリズムが区別されていない動的な情報処理" のような理念のもとで研究を進めることで、階層の概念を脱却することが可能であろうか。それとも、( "確率"の場合と同じように)"階層"も、いつまでもどこかに潜んでいて、われわれがそこから脱却したと思った瞬間に再出現するような概念なのだろうか。」ここまで引用。
単回のイベント(FB)から複数回のイベント(FF)というイメージで、後者が現象的なものが出来るために必要なのではないかとぼんやり考えていたのだけれども、見直してみればこれもやっぱり表象だ。表象以前に現象はあるのだろうか。
でも、生成の過程をイメージするに、表象が出来るのと同時に現象が出来たと考える方がもっともらしい気がする。強い表象主義(現象というものはなくて、表象だけがある)には与しないけど、表象が出来る程度の複雑さが現象には必要といった弱い表象主義は知覚運動説となじみが良さそうに思う。
もちろん、このときの表象はカルテジアン劇場的なものではなくて、断片的なものでよくて、なんらか操作できさえすればよい。そしてそんなもんだったらわざわざ表象主義とか言わなくてもよさそう。
2011年07月27日
■ MacBook Pro 2011で120Hzできた
神経心理の実験の準備のために、刺激提示について検討してた。いろいろ道具を持って出張するということになるので、CRTではなくてLCDで刺激提示をすることにする。
刺激提示装置の選択は視覚科学の分野ではいつでも問題で、CRTモニタが年々入手が難しくなってゆくところでどうやって心理物理的実験を行うかというのはしばしばMLにもポストされる。
つまり、CRTでの時間応答と比べて、LCDでの時間応答は遅い。年々スペック表に出ている「応答速度」は速くなってゆくが、これは白->黒での値だったりして、中間色のあいだでの時間はもっとかかる。
そういうわけで、motion刺激を出すような場合にヘンに残像とか出たりすると困ったことになる。(静的な刺激に関しては事情は違っていて、液晶のIPS方式だと色再現や視角がよかったりして、医療での画像診断用のモニタは液晶だったりする。)
とはいえ、LCDでランダムドット刺激使って心理物理やった論文とかも出てはいる。"Speed and Eccentricity Tuning Reveal a Central Role for the Velocity-Based Cue to 3D Visual Motion" (JNP 2010) LCDを使うに当たってはMethodsのなかでそれなりに確認したことが書かれている。あと、続編では120HzのDLP projectorも使っている。"Motion processing with two eyes in three dimensions" JoV
そんなこんなで気にはなっていたのだけれど、今年のvisionlistのポストでLCDでもけっこう大丈夫ですよ、という論文の原稿が公開されてた("An LCD monitor with sufficiently precise timing for research in vision" (pdf))。この論文ではネイティブ120HzのSAMSUNG 2233RZとNVIDIA Quadro FX580の組み合わせを使っていた。
なるほど、たしかにだんだん120Hzのものが増えてきているようだし、これを使うことを検討してみた。
グラフィックボードに関してはNVIDIAはGeForceがDirectX特化(ゲーム用)で、QuadroがOpenGL特化(科学計算用)ということらしい。私が使うとしたらPsychPyにしろPTB3にしろOpenGLベースとなるのでやはりQuadroか。
液晶ディスプレーの方はネイティブで120Hz出るもので今いちばんよいのはBenQ XL2410Tか。これはTN方式なので欠点はあるが、応答速度も速い。(比較記事) 三菱とかはIPS方式で色再現や視野角はよいけど、これはネイティブ120Hzではないので私の用途には向かない。
ほかにもいろいろと調べていたのだが、この世界はFPSゲーマー向けがいちばん進んでいることがよくわかった。LCDはだめでCRTがいちばん、とか視覚科学者と同じこと言ってて親近感がわく。
たとえば、120Hzにしても人間の目が追いつくのか?なんて議論をしていて、ソニーの人の論文 とかに言及していて、240fpsあたりが限界だって話をしている。ちなみにこちらはpdfが取れる: Development of the High Frame Rate 3D System
平行して、macのラップトップでも刺激出せないかどうか検討した。MacBook Pro 2011にはMini DisplayPortからDual-Link DVIに信号を送ることができるようになっているのだけれど、外付けモニタで120Hzは出せないのだろうか? webのサポートページとかではシネマディスプレーとか大画面用には出せるようだが、リフレッシュレートについては記載がなかった。
もうちょっと古い型ではうまくいかなかったと書いてあった:120Hz monitor usage ただし、グラボの性能としては120Hz出るようだし、switchresxを使って120Hzが出せるという情報もあった。
以前書いた120Hzモニタの件。BenQ XL2401TをMacbook Pro (2011 Spring)とDual-Link DVIで繋いでみたら、ちゃんと120Hz出せることが確認できた。PsychoPyでtime by frameを計算すると、平均8.3ms (= 1000ms/120frames)。
ただし、スクリプトの立ち上がり(<100ms)のところで盛大にフレーム落ちすることがある。
Macbook ProはふたつのGPUを切り替えているので、それのせいかなと踏んで、「省エネルギー」のところで 「グラフィックの自動切り替え」をオフにする。これで常に高性能グラフィックの方が使われるようになる。これでうまくいったかなと思ったらまだframe落ちすることがある。
よくわからん。ただし、frame落ちするのは初めの方だけなので、どっちにしろ刺激を出す前に待ち時間をおくことになるし、私の用途ではとりあえずはよしとする。
参考までにLCDの応答特性とかを評価した論文いくつか:
2011年04月26日
■ 階層ベイズなdiffusion model
たぶんどっかで「信号検出理論と応答潜時のaccumulator modelとが統合されてconfidence levelとかの議論ができるようになるべき、俺の仕事じゃあないけど」みたいなことを書いた気がする(*)けど、これはけっこうそれっぽい: Two-Stage Dynamic Signal Detection: A Theory of Choice, Decision Time, and Confidence
(* あとでブログ調べたらこれのようだ:Confidence in LIP「"Log posterior odds = log-likelihood ratio + log prior odds"という式はaccumulator modelとSDTとを組み合わせるために有望なものです」もっといろいろ書いてた気がするのだけれど。)
Constraint-freeな状況でライフログ的に膨大なデータを獲得するというやつは階層ベイズモデル(とその仲間)と組み合わさることで威力を発揮すると思う。つまり、たくさんのデータがあるところで、構造を考えずにデータマイニングすると終わりが見つからない。いっぽうで、充分リアリスティックな階層ベイズモデルをつくるためにはそのパラメーター空間を充分埋めてくれるような大量な多次元データが必須だ。
このふたつの組み合わせによって生成モデルを作ることができる。生成モデルを作ること自体が理解するということであり、統計モデルでしかないという意味ではこれがまず第一歩。もちろん、本当は統計的モデルではなくて、動力学的なモデルに行きたいのだけれども、その前を固めることが重要。
そういう意味で以前「生態学的アプローチを見習ってGLMMとか使ったらいいんじゃん」とか書いた。実際、この論文(pdf)のfig.2とかシビれる。こんなかんじで、電気生理から解剖学まで全部ツッコみたい。
アイデアの発端は以前のJNS論文で使ったdiffusion modelなんだけど、あれはかなり大胆にモデルを簡略化してパラメータたった二つにしたことでなんとかなった。大元のdiffusion modelはもっとパラメータが多い。当時はこんなパラメータ多かったらなんでもfittingできちゃうでしょ!とか思ってたけど、逆に行くならば、たくさんのsubjectとたくさんのセッションとたくさんの条件でたくさんデータを取ればたぶんもっと複雑なものでもいけた。じっさい、nhpのデータのいいところは、データ数がsubjectあたりで万を超えるってところ。ヒト被験者ではこうはいかない。
Diffusion modelはそれでも脳の中の過程はぜんぜん考えてない。個人差とかいろんなrandom effectを取り込んでいけばよいでしょうなんて思ってたら、Ratcliffの共著者だったTuerlinckxによる"HIERARCHICAL DIFFUSION MODELS"(pdf)ってのが出てきた。まだ読んでないけど、こんなかんじでいきたい。
書いててわかったぞ、つまり、Ratcliffの論文読んでるといつもなんか歯がゆい感じがしてたんだけど、つまり、データがショボイくせに複雑なモデル立ててなんとかしようとしてたからなのだな。驚くべきことに、Ratcliff論文ではsubjectのデータをmergeしている。そうしないと充分なデータ数が集まらないからだ。つまり、データとモデルのバランスが悪いからなのだな。これがさっき書いたことと繋がる。
(ついったに書いたことを元にして編集して作成した)
2011年04月15日
■ Neurobehavioral Systemsのpresentationをいじり中。(2月に)
さて今日はNeurobehavioral Systemsのpresentationをいじり中。30日間使用できるので、とりあえずwindowsにインストールして、デモプログラムを動かしている。
デモプログラムを始めるのにリターンキーを押す必要があることに気づくのに30分かかるとかそんな調子。いまはマウスが日本語名であるためにプログラムから認識されない問題に対処中。娘は隣で宿題やってる。そんな休日。
日本語の外部機器(マウスとか)が見つからないって言われる問題はExperiment File(*.exp)のなかにその情報があるのか。いつも上書きしますか?と聞かれて嫌だと答えていたから同じこと何度も繰り返してた。
なんど日本語の「マウス」を指定してexpファイルを保存しても、読み込み時に文句言われて実際マウスが指定されていないので途方に暮れていたが、設定をよく見たらencodingがUTF-8でなくてsingle-byteであったことに気づいたときの徒労感は異常(<-なげえよ)。
Neurobehavioral systemsのpresentationいじり中。とっかかりになりそうな実験ファイルがあったのでそれを元に改変してゆく。だんだんわかってきた。
実験ファイルはおもに三つの部分で構成されていて、ヘッダとSDLとPCLがある。ヘッダに定数を置いて、SDLでクラスのインスタンスを作って、PCLでクラスのメソッドを使って動かしてゆくのだな。なるほどなと思ったのは、trial自体もクラスで、その中で提示時間とか刺激とかも定義する。
TEMPOのCライクな文法と比べるとこちらはオブジェクト指向な、もうちょいモダンな文法だ。10年前のわたしには無理だったかもしんないけど今なら使えそうだ。
たとえば赤い四角を定義するとしたらこんなふうになる。
box {
height = 100; width = 200;
color = 255,0,0;
} red_box;
これでboxクラスのred_boxっていうインスタンスというかなんかを作る。あとは同じ要領で、刺激のパーツ(box)、課題の画面(picture)、課題のスケジュール(trial)が階層的に定義されていて、そのインスタンスを作ることができる。
たとえば赤い四角を一秒間提示してもう一秒経つと終了、という課題を作るときにはこういうふうにする:
#----------------------------
begin;
box {
height = 100; width = 200;
color = 255,0,0;
} red_box;
picture {
background_color = 0,0,255;
box red_box;
x = 0; y = -100;
} red_pic;
trial {
trial_duration = 2000;
picture red_pic;
time = 0;
duration = 1000;
} red_trial;
#----------------------------
begin_pcl;
red_trial.present();
ぱっと見でだいたい何してるかはわかると思う。静的なところはbegin;以下の部分(SDL)に書いておいて、動的なところはbegin_pcl;以下の部分(PCL)に書いておく。たとえば提示する図をシャッフルするとかはPCLで。
クラスの定義のところ自体はもうありもんとしてreference manualに記載されているけど、たぶん頑張れば派生クラス作ったり、一からクラス作ったりすることはできるのだろう。
かなり進んだが、いまだにstimulus_dataとresponse_dataの違いとか分からない。stimulus_data.sceを読めばいいらしい。なにが気味悪いって、hit-missはstimulus-responseの関係なのにresponseの方に入っている点。
いまどきパラレルポートのあるPCなんてもうないから、デジタルの入出力をどうするかはけっこう問題らしい。 以下presentationのforumから:
- fMRI : Detecting a TTL pulse using USB
- fMRI : USB port for response during fMRI
- Hardware Interfacing : Output via USB port
- fMRI : Recommendation for a USB digital I/O device
Forumが充実しているってのは素晴らしい。
まあとりあえずTTL->USBコンバータ 買うのと、NIの一番安いUSB DAQユニット買うのの二本立てでなんとかしようと思う。
TEMPOだとひとそろい集めるので250万円くらいかかるので、presentationでPCひとつと5万円ってのは気楽だ。心理学的な課題(Visual searchとか)はたぶんこれでよい。視覚心理物理がこれでいけるかまだ分からない。PsychoPyとかいろいろ出てるし。
PCがどんどんレガシーなものがなくなっていって、CRTとかパラレルポートとかシリアルポートとかに依存しているものはどんどんやりにくくなっている。PsychoPyとかpsychotoolboxとかがどうやっているか分からないけど、OS上でやってるかぎり問題は変わらないはず。
たぶんもうソフトウェアだけでの解決ってのは無理なんだろう。なんかreal-time OSを走らせている外部boxを使ったシステムとして開発すればよいのではないかと思うのだけど。Matlabの場合でいえばxPC TargetBoxみたいなかんじ。
TEMPOは心臓部分をDOSのPCで動かしているのと形式的には同じだけど、TEMPOの場合はいまどきrealtime-OS部分にでかいPC一個あてがって(PCIボードを入れる必要があるから)、しかもフロッピーで起動とかクソ過ぎる。実験前に起動が引っかかったりして冷や冷やしたよ。
組み込み用のシステムだったらシリアルだってパラレルだって現役だろうし、なんか新しいことをする必要はないよね。中身ビーグルボードで1個50万で売ればじゅうぶん価格破壊で、年に50個売れて2000万円くらい儲かるんでないの? (<-まさに素人の床屋談義)
でもそれよかおしゃれなのはオープンソースハードウェアの流れで、外部boxとクライアントPCのプログラミング言語と全部込みでフリーってのがいいよな。たぶんそういうの支援する競争的研究費があるはずだから、業界への貢献としてそういうプロジェクトを立ち上げても良いんではないだろうか。
これも俺がやる仕事とは思えないけど、こういうのは使いたい人が作らないといいものにならないし(作ったけど動いてないシステムというのがあることは知っている)、オープンソース的思想でもってやります、ってだけで充分貢献だと思うけどなあ。
バカな質問なのかもしんないけど、そういう目的で科研費とって、オープン化前提で業者の人にデザイン・実装してもらうって可能ではないのだろうか。データ共有、解析関連の話にも繋がるけど、使いたい人が作るってところが大事だと思うんだ。
だがそんなことはどうでもよい。それより気がかりなのは、「今日の雨、傘がない」ということであって、枯れてたアジサイにこの冬いっぱいダメくさいと思いつつも水を与えてきたけれど、やはり芽は出ず、やっぱり無理だと決断して引っこ抜いたので、この植木鉢を今後どうするのかということだ。
(ついったに書いたことを元に編集して作成した)
2010年07月24日
■ 勝っている人たちは当然、私だって少しは。
「医療過誤訴訟を生き残る」 というエントリで"How to Survive a Medical Malpractice Lawsuit: The Physician’s Roadmap for Success"という本が紹介されていて、そこではたとえばこんなかんじでプロテクティブにやることを勧めている:
公判前の質問を受ける場面においては、病院の内部でしか通用しないような略語を、むしろ多用するように心がけたほうがいい。原告側弁護士は、しばしば言葉の説明を求めるかもしれないが、略語を多用することで、相手に伝わる情報を、より少なく保つことができる
裁判ってこうやっていかに失点を防ぐかみたいなことをガチガチにやることなんだなと思うと、そういうのが日常の人もいるのだろうということに思い当たって、そら恐ろしくなった。(なんて甘ちゃんなことを言ってる、と思うかい?)
ちなみに学術論文が無味乾燥なのも同じ理由から来ている。学術論文ってのは揚げ足取られないように細かい表現のレベルを調整してぎちぎちに書いてあるものだからだ。(以前、法律の文章のようだ、と書こうと思ったことがあったけど、じつのところ法律の文章がどんなもんだかわたしはわかってない。)
たとえば、ある機能のneural correlateを見つけたという論文を書くとしよう。この論文の結論として、そのニューロンが機能に関わっているとは絶対書けない。機能に因果的に関わっているかどうかは不明だし、同じニューロン活動は脳の別のところにもあるかもしれない。損傷実験やムシモル注入実験とかの関連する報告を踏まえたうえで、機能との関連はhighly likelyだ、とか言うのにとどめることになる。
Neural correlateの研究というのは結局、脳の中にこれこれこういう情報があるのを見つけましたよ、ということであって、本質的にはデコーディングと同一なんだと思う。だからこそ最近の気の利いた論文ではneural correlateの代わりにデコーディングの文脈でものを言ったりする。たとえばfMRIでワーキングメモリをV1からデコーディングとか。わたしはダッサいながらも戦略的にneural correlateについて論文を書いていこうと思っていたけれど、のろのろしているうちにあっという間に古くさくなってしまった。
脱線するけど、かといって、刺激実験をしたからって因果的に機能に関わる、みたいな結論にするのも全然足りない。刺激の下流が関わっている可能性もあるから。損傷とかにしてもそうだけど、そういうすごく単線的なロジックが求められているので嫌いだ。これは認知科学的な機能モジュールの発想を引きずっていて、コネクショニズムとかを通り過ぎた後の発想ではない。たとえば、ある部位を抑制したからってその下流が押さえられるとは限らない。
だから私としては、もっと計算論的モデルをいれて、あるところを刺激したらほかのところがどうなるかと動態を予測するという方向へ行くべきだと思うのだ。そっちの方向へ行けば、かえって予測の精度は下がるだろう。それでも良いと思う。そういう試みが並行して走れば、イノベーションはどこかで起こるだろう。
私が「イノベーションのジレンマ」で好きなのは、3.5インチHDが現れたときとか(さらにいまだったらSSDでも)、破壊的イノベーションが起こったときは、新興が手がけていて、スペックとしては先行するものと比べたら低かったということだ。しかし最終的にはその機能を上げて、先行者を追い越した。(これを胸に置くことは良いことだけれども、だからといって逆は真なりとは言えない。当然。だからこそ、そういう試みを並行して走らせればいいのだ。)
いまmulti-unitやLFPやECoGを使っている人は、single-unitだったらできたような個々のニューロンに対する課題条件の最適化を捨てて、その代わりに得られるものを求めているんだと思うし、私も次のプロジェクトは全面的にそちらに向かっている。
だからこそ、わたしが目を向けているのは、そういった新しい実験パラダイムでのデータのサンプル法(overtrainingと繰り返し前提の行動課題のデザインに留まらない)についてだ。つまり、大澤先生がやってるような刺激空間をいかにして網羅的かつ効率的にマップしてゆくかという問題だ。
しかもそれを知覚だけでなくて、知覚と行動の連関、つまり環境-個体のinteractionのレベルでマップするにはどうすればよいのかと考えている。はじめから網羅的にマップしなくても良いけど、それでも操作的脳科学になるようにぐるっとひとまわりできるようにしておく必要はあるだろう。
ぐるっと話を戻すと、それでも学術論文を無味乾燥でなくエキサイティングなものにすることは可能だし、読んでてこれは面白いと思ったことはいくらだってある。だからこそ論文コメントブログなんてやってるわけで。うまい人はホントにうまく書く。私もそうありたいと思う。自分の論文が毎度日本人的なこまっかな議論に落ち込んでいくのをなんとかしたいと思ってる。ぶっといメインのラインをいかに印象的に書くか(スティーブ・ジョブスの演説みたいにね)、あれこれひっくり返して考える。
それに揚げ足取られないようにきっちり書いてない論文ってのは、もう安全地帯にいる大御所か、逆に泡沫方補の演説みたいな論文にしかならない。揚げ足取られないようにきっちり書いている論文は、高校野球の試合で選手がきびきびしているのと同じようなさっぱり感というか、基本ができているかんじがある。ここをないがしろにして良いなんてわたしはぜったい言わない。
そもそも両者(エキサイティングであること/揚げ足を取られないこと)は両立する。みんなが苦労しているのは、NatureやScienceに載るほどエキサイティングな主張でありながら、揚げ足を取られないような完璧な論理構成を作る点にあるはずだ。
さらに話を戻すと、医療訴訟のように失言が命取りな世界で戦いを日常にしている人がいて、そういうのにわたしはビビったりするけれども、じつのところ研究の世界も同じなわけで、まさに戦いを日常とした世界にいて、いかに相手を黙らせるか論争的でありつづけている。
勝っている人たちは当然、私だって少しは。
追記。Twitterではこうやって話を閉じたけれど、わたしはいま自分がそういう戦いの世界にいるというイメージは持ってなくて、いかに想像力をぶっ飛ばしていけるかということに重きを置いている。その世界では、戦いの世界のように萎縮していたら、想像を広げることはできないと思うんだ。わたしが何度も繰り返して表明してきた「恫喝的コミュニケーションへの反発」とはそういった世界観の部分で繋がっている。
今日のエントリでは、A-B-C-B'-A'という変な構造を採用して、いつもどおり直線的なロジックを回避した。当然、A-A', B-B', Cと話を分けるべきだ。そうしなかったのは、A-B-Cの関連性にインスピレーションをかき立てられたからで、そういう意味では『脱線だけど』とか書いたけど、本人は全然脱線だと思ってない。それが論文で書くような直線的なロジックとの違いのはずで、そういうことが伝わると良いのだけれど。(<-今日いちばん言いたかったこと。)
「刺激空間をいかにして網羅的かつ効率的にマップしてゆくかという問題」についてはもう少し書いておきたい。と思ったがすでにエントリが長い。またにします。(<-こいつゼッテー続ける気がない)
2010年07月17日
■ 日経サイエンス記事を翻訳しました
どうもご無沙汰してます。LAから帰ってきて、アムスに行ってまた帰ってきて、肩まで伸びた髪を切って、西海岸風ヒッピー系ナードから、元のだらっとしたかんじに戻ってきました。さていろんなことが溜まっている。
で向こうに行っているあいだにひとつしたのが、日経サイエンス記事の翻訳です。ちょうどいま発売中の日経サイエンス2010年8月号に出てます。
タイトルは"Uncanny sight in the Blind" 「盲人の不思議な視覚」 タイトルからわかるように、Blindsightの話です。元記事はScientific American 5月号のdeGelderの記事です。けっこうタイムラグが少ないなって印象があった。翻訳作業は4月だったけどそのときはまだ元記事が出版されてなかくてゲラ状態だったし。
deGelderはオランダの人で、Weiskrantzのところでやっていたaffective blindsightで有名。つまり、emotionに関連する視覚情報(怒った顔とか)はV1を通らずに直で扁桃体まで行くって話。
「英語で読む日経サイエンス」っていうページで私が訳した記事の訳文と元の文章が並べられてます。恥ずかしい。これをみて受験生が勉強に使ったりするのだろうか。このページの中にある例だと、"sidling"を「カニ歩きする」とかちょっとおちゃらけぎみに訳してみたりした。"it also does not make sense"は「意味をなさない」とか堅めに訳したら、編集部が「支離滅裂である」という表現に直してた。いま思うに「あり得ない」くらいでも良かったかも。
視覚障害の話題なので言葉の選び方とかえらく注意したのだけれど、元記事のタイトルは"Uncanny sight in the blind"。Uncannyって例の「不気味の谷」の「不気味」ですよ。どうなん、それ。タイトルの写真は目隠しをしている男性の写真なのだけれど、その目隠しに目が描いてあって、明らかに不気味な感じを意図している。辞書の定義では"seeming to have a supernatural character or origin: eerie, mysterious" なんで「不思議な」と処理しておきました。とりあえず間違ってはいないと思うんだけれど。
ともあれ、これが私にとって商業出版で翻訳をした初めての経験だったのでなかなか難儀しました。これまで科学系の翻訳書を読んでは、これはひどい訳だ、私がやればもっとうまくやるのに、と思ったものだけど、自分でやってみるとやっぱり難しい。いかにかみ砕いて表現するかとかそのへんはたぶん、労力と時間をかければもっとマシにすることはできる。しかしやっているときりがない。たぶんこねくりまわさずに、ざっと行くスピード感が必要なんだろうなあ。
大学生くらいの頃はアメリカの現代小説(アーヴィングとか)の訳書とか読んだりして翻訳に興味を持って、バベル・プレスの「翻訳の世界」(いま調べたら名前が変わってる!)とか購読したり、安西徹雄の翻訳英文法(新装版)読んだり、関連するワークブック(「英日翻訳トレーニング・マニュアル」)とかやってたことを思い出した。今回も、そういえば名詞句が主語の時は副詞句的に訳すんだ、とか思い出した。訳例で言えばこことか:"These discoveries spurred further systematic investigations of animals lacking the primary visual cortex. " 「これらの発見に刺激されて,第一次視覚野を失った動物を使った系統的な研究が進められた。」
だらだらしてきたんでこのあたりで。ではまた。次回はASCONEおよび生理研研究会について。
P.S. 「翻訳の世界」についてネットで調べてたら見つけた:『翻訳の世界』元編集長 今野哲男さんにきく
2010年03月08日
■ シェイピングの十の法則
私たちの実験で非常に重要だけど体系化されていないものに、被験者のトレーニングがあります。
基本的にオペラント条件で教えてゆくので、スキナーの行動分析について知っておく必要があるのだけれど、トレーニングに特化したものというのがないんです。それで、みんな試行錯誤でそれぞれにやり方を確立してゆくというところがあって、体系化されていません。
一時期いい教科書がないか論文などをあさってみたことがあるのだけれど、ほぼ皆無です。(採血を自発的にやる方法を確立したViktor Reinhardtの仕事などはありますが、システム神経科学でcognitiveな課題遂行中のニューロン活動を記録するためのトレーニングというのはほとんど見たことがありません。)
私が唯一知っているのは、一般書ですが、「うまくやるための強化の原理」カレン・プライア著 というものです。
カレン・プライアはイルカのトレーニングにオペラント心理学に基づいた方法を導入し、犬でのクリッカートレーニングなどでも本を出してます。たしかに、犬のしつけ法とかのあたりも参考になりそうだ。
でもって、この本あんまり見かけないんで(わたしはbookoffで100円で買った)、このなかにある「シェイピングの十の法則」というのをメモっておくことにしましょう。
- 十分な数の強化が得られるように、基準を少しずつ上げよ。
- 一時に一つのことだけを訓練せよ。
- 基準を上げる前に、現在の段階の行動を変動強化で強化せよ。
- 新しい基準を導入するときには、古い基準を一時的にゆるめよ。
- 相手をたえず観察せよ。
- 一つの行動は一つのトレーナーが教えよ。
- 一つのシェイピング手続きをやっていて進歩しないときには、べつにやり方を見つけよ。
- 訓練をむやみに中断してはいけない。
- 一度できた行動でも、またできなくなることがある。そのときは、前の基準に戻れ。
- 一回の訓練は、できれば調子が出ているときに止めよ。
これはどれも自分の経験と照らし合わせて、だいたい納得がいきます。基本はなにより「相手をたえず観察せよ。」ですけどね。
このくらいであっさり終了とします。ではまた。
追記:こちらにも類似したことが書いてありました:ABA(応用行動分析)
# 桑原
僕もサルを使っているので最近ですが読んでみました。飼い犬から配偶者まで、っていう副題がなんともたまらないなと思いました。
3のところはどう思われますか?たまに休んでいるときとかモチベーションが低い時にBIG REWARDをあげるとしばらく頑張って正答率も上がるのは感じますが、長期的に見てどうなのかはまだまだ経験が浅くて分からないところです。
10番も難しいですよね?より高く跳んだら良い、みたいな状況だと、トレーニング終了自体大きな報酬ですしよく効きそうな感じがしますが、僕らの場合やっぱり長くタスクを続ける事が重要なことなので難しいとこじゃないですか?
どうもありがとうございます。
たぶん、イルカに芸を教えるような状況だとどんどん新しいことをやっていく必要があるけど、我々のトレーニングの場合、fixationの時間をじりじり延ばしてゆくとか、かなり漸次的にやってかなければならないところが多いのが違いかと思います。
新しい課題をできるようになること(たとえばvisually guided saccade taskからmemory-guided saccade taskへの移行)とタスクのパラメータの漸次的に変化させてゆくこと(recordingのためにtrial数を多くできるようにするとか、fixationの時間を伸ばしてゆくとか)とは別ものですが、ここでのシェイピングの法則は新しい課題をできるようにする状況で特に重要なこととなるかと思います。
3での変動強化はFRではなくてVRにするということですから、われわれの課題のようにtrialごとにrewardをあげる条件でrewardの上げ下げをするのとは別ものだと思います。
10に関してですが、たとえば、セッションの後半では新しいことをやらせずに同じパラメータで回数こなすことに専念すべき、というようなことに繋がるかと思います。
大変勉強になりました。ありがとうございました。やはりVRを使いこなせるトレーナーは数少ないともありましたし、我々の状況ではあまり使えないようですね。
# pooneilVRとかVIのような変動強化スケジュールはもっと活用する価値があると思ってます。これまで使われてきたのは、もっぱら強化学習関連に直結した仕事(設楽先生のとか、鮫島さんのとか)だったわけですけど、たぶんもっと普通のトレーニングでも活用できるんじゃない買って思ってます。
われわれはだいたいtrialごとにrewardをあげてますけど、被験者はそんなにtrialごとのフィードバックを使ってるわけでもなさそうです。私たちトレーナーの方が、大人の人間にものを教えるときの方法に縛られているだけなのかもしれません。
以下は神経科学者SNSに書いたことの転載:子供のしつけでも共通性を感じることがあります。5に関して言えば、相手をよく見ておくことは重要だけど、おもねってしまうのはよくない。こちらの基準がふらつくのは良くないので、どのタイミングで基準をいじるか、ここが腕の見せ所だったり、個人差が出るところだったりすると思ってます。
2009年12月12日
■ Pooneil Radar 2009
いろいろ書きたいネタが貯まっているんだけど、もうすこし調べておかないとしょうもないことしか言えないという中途半端な状態なので、ネタ帳的にいろいろ書き付けておきます。
タイトルの元ネタは"O'Reilly Radar"っていう、ティム・オライリーのレーダーにかかった今後面白くなると思われるネタ集みたいな話で(たとえばIT conversationのpodcastでOSCON2004のときのトーク)、それをもじって命名するとはどんだけオレも自我が肥大化しているかっていう(って書くと後輩が「吉田さん、どんだけの使い方間違ってます」って指摘してくれるという親切システム。)
-
以前からgeneralized linear mixed model(GLMM)について調べてる。fMRIの世界ではFristonのおかげでrandom effect modelがSPMに導入されて標準化されたので、複数の被験者のデータからのinferenceにrandom effect modelが使われるようになった。
一方でnhpのsingle-unit studyではこのへんが遅れている。fMRIと比べて、被験者の数が少なく、記録しているニューロンの数や、繰り返しの試行数が多い。だからたいがい被験者のデータはmergeするし(必要に応じて被験者ごとに検定したり)、試行数の繰り返しは平均して使ってしまう。これはよくない。
データ構造は一般的には、被験者数がn=2-3で、その中で複数のニューロンを記録して(n=100ずつとか)、さらにそれぞれのcellで各条件で最低限10trial以上の繰り返しを行っている。つまり、nested designのmixed modelになってる。Rのmodel式で書くと(lme4でのnotation)、Firing rate ~ condition + (1 | Subject) + (1 | Subject:Cell) + (1 | Subject:Cell:Trial)みたいな構造を考える必要がある。しかも応答変数Firing rateの実体はNumber of spikesだから、計数データとして"generalized model"にしとかないといけない。
-
このへんを勉強しようとすると、生態学がいちばん進んでいるのがわかる。生態学はフィールドでどの区画にどれだけの個体がいるかみたいな情報を扱うから。北大の久保拓弥氏のサイトとか見るとたくさん資料があって、超助かる。あと、GLMMが経験ベイズの一種であって、階層ベイズによって包括されるというこの図を見て感激した。こういうことをもっと知りたい! (いろいろ読んだけどここでは省略。)
階層ベイズをもっと体系的に組み込んで、実験系全体をモデル化するようなことに興味がある。たとえば"Hierarchical Modelling for the Environmental Sciences"の4章の図4.2見てほしいんだけど、こんなかんじで脳の解剖学的な拘束条件とか全部突っ込む。
- いろいろあってneuroshare関連の話に多少関わってるので、話が通じるくらいには勉強してる。要は自分の解析と一体化させることが出来たらいいんだけど、neuroshare形式を使った解析ソフトがそんなにあるわけでもないのですよね。FINDとかがmatlabが主体になっているのは実際的ではあると思うけど、オープンソースという観点ではRとかでできないっすかね。上記の問題意識からすると、Rベースでunit/LFP/ECoGにとってのSPMみたいなの作ればさ、Fristonになれるんじゃないですか。だれかやって!
-
nhpを使った実験をしてゆくにあたって、いかにして適切なトレーニングをしていくか考えなければならないとずっと思ってた。そのためにはpositive reinforcement techniqueに基づいたトレーニング法を採用するのが正しいはずだ。しかしこの分野は資料が少ない。J. Appl. Anim. Welf. Sci. 5(4)に特集があって役に立つ。単行本だけど「うまくやるための強化の原理―飼いネコから配偶者まで」カレン プライアが名著で、前述の特集号でも参考文献として引かれてる。
ちなみにこの人はイルカのトレーニングの創始者で、犬のクリッカートレーニングにも大きく寄与してる。しかも"The creative porpoise: training for novel behavior"のファーストオーサーだった! (これはベイトソンの本にも出てくる「それまでにしたことのない行動をすると強化される」というメタな学習の例)
あと、「動物感覚」も良いことが書いてあるのでまとめておきたい。スキナー的なものへの敵意が混ざっているので取扱い注意だけど。
ともあれ、微妙なissueなので、書くときはちゃんと書かないといけない。 - このブログはMovable typeのver.3.33で出来ていていい加減古いので、アップデートしようと思ってる。Movable type 5が出るって発表が出たあたりから、アップデートするか、wordpressに移るか決めるためにテストしてました。Ubuntuの9.04のserver editionが出てたんで、virtualboxに入れたり、vmwareに入れたりして。でも最終的な決断が出来ず。正直いまのままでもいいやって感じで、テンプレートいじったりしていろいろ工夫する情熱を失ってる。
-
以前のエントリでNIPS-SSCのregistrationとかabstract suibmission用にOpen Conference Systems (OCS)ってのをいじってるというのを書いたことがありました。じつはいろいろテストをして、いろいろ手を加えて(無駄な階層をどんどん削って)、とりあえず稼働できるところまでは来たんだけど、けっきょく扱えるのがわたし一人で、それだったら手動でやっても余り手間が変わらないということでけっきょく止めにしました。
というわけでその間の奮戦記とかいろいろあるんですが、まとめるのさえ面倒になってしまった。どうしてだれも日本語化とかしないですかね。でもこの方向で行くのが正しいと思うので、機会があったらまたチャレンジしたい。(ここ最近のわたしの行動を見てもらうとわかると思うけど、基本はDIYでいこうってわけ。) -
Facebookのアカウント取って細々と使っているのだけれど、せっかくだからもっと活用したい。とくにNIPS-SSCつながりでFriendが増えたので、英語でなんか書いておきたい。というわけで、ブログで英語のエントリの作成をして、それをFacebookに「ノート」としてimportしてる。
こっちは日本語だと書きにくいこととかを書こうと思う。たとえば、秋くらいに"The Electric Kool-Aid Acid Test"(の訳書)を読了していまは原書で読み直してるところなんだけど、へんに誤解されないようにこういう話題は英語に持ってゆく。 - なんかポストロックっぽくギターをかき鳴らして、ビデオに撮って、YouTubeに挙げて、埋め込みでエントリ作るってネタがずっとあるのだけれど、実行している暇がない。キャプションに「お遊戯的なことなら 外でやってくんない?」って入れるところまで決めてあったのに、つーかクラウザーさんネタだったのにもう古くて使えないぢゃないか。
- 将来のプロジェクト関連。これはいろいろ。パーツ自体よりは組み合わせが大事だと思うんだけど、こういうのはいろんな人のところに行って吹聴しては、反応見てる。
- 自分の健康についてはいつかきっちり書いておこうと思う。ってほんとうに単なるToDoリストになってしまいました。Get Things Done! 話を戻す!
- 以前に「細胞外電極はなにを見ているか 」というのと「細胞外電極はなにを見ているか リニューアル版」というエントリで、細胞膜でのイベントから細胞外電位の発生までの生物物理的な話をまとめたことがありました。あそこで足りなかった視点のひとつは電極の特性の件でして、電極のコンタクトの面積や抵抗でどのように波形が影響されるかみたいな話をちゃんとまとめておかなければと思ってます。つーかわたしである必要なんか無いんだけど。誰かやってほしい。ほかにも、microstimulationについてとか、電気生理用のアンプについてとか、わたしはそっち方面のバックグラウンドがないので、将来のため、教育者として、きっちり勉強しておきたいと思ってます。
- Saliency mapについては将来のプロジェクトのひとつではあるけど、もっといろんな繋がりが可能なので、もうすこし自分で使えるようになっておきたい。たとえば、Ittiはあくまでスタンドアロンでのneuromorphicなシステムとして捉えているようだけれど、OpenCVとかで使えるようなライブラリにしちゃえばいいんじゃん?とか思う。あと、あらかじめシステムとして構築するのではなくて、data-drivenにシステムを作る方向へ行きたいですよね。実際そういうことをやっている人はすでにいる。たとえばFelix A. Wichmannの"Non-linear System Identification: Visual Saliency Inferred from Eye-Movement Data"とか。(そういうわけでPRMLとか読んでるわけ。)
-
Overtrainした状況でのくりかえし行動での脳活動というパラダイムを越えてゆくためには、膨大な行動データから特定の行動を自動的につり上げてくるというようなことが必要であって、そういうアルゴリズムを開発している人はいないのだろうかと思ってた。わたしのPCには「行動のデータマイニング」というフォルダがあって、そこには去年の冬あたりにサーベイしたときのファイルが残っていて、「センサネットによる行動パターン識別システムの開発」とか「バイオロギング研究所」(リンク切れ)とかそういうのを集めたところまで行って、止まってた。
そしたらATRの神谷さんがいろいろ教えてくださって、一挙に道が開けた。そもそもわたしは「エソグラム」という言葉すら知らなかったのだけれど、ここに広大な領域があるのを知った。ということでここすごく勉強したい。
って書いてたらそれなりに分量はあるな。計画を書くとそれで満足してしまうというありがちな展開を経ながらも、このブログは続いていくよ。普通頻度が下がってゆくとどっかで静かに消えるというのがありがちなブログの終わり方なのだけれど、一ヶ月更新がなかったあとでいきなり週に3回書いたりするのがこのブログ。RSSに入れてチェックしておいてください。(7年目に突入したよ!!)
# viking
Human fMRIと違って、nhp single unit recordingは測定そのものに「ノイズが少ない」のですから、自ずとfMRIにおける統計とは考え方が変わってくるのではないでしょうか?
というのは、ヒトfMRI実験におけるBOLD信号が、blurring effectだの血管効果だの、はたまたin-flow effectとBOLD effectとのせめぎ合いだの、といった多くの未解明な複合要因のバランスとして得られるものであるという事実を考えると・・・あくまでもtaskなりeventなりの効果だけを見るためにfMRI(のSPM解析)でrandom effectsに着目するのはnoisyな測定系であることを考えれば当然の帰結でしょう。
これに対して、それほどnoisyではないnhp single unit recordingではそういう問題を考える必要性は薄くて、なればこそ個体数n = 2,3でも許容されるということだと思うのですが。
ですから、ここでGLMMなり別の統計を提案するということであれば、考慮すべきは何よりもまず「どの測定量がばらつくのか」ということを事前に仮定するということだと思います。それが個体間であれば個体数nを増やしてSPMのrandom effectsライクにいくしかないわけですし、ニューロン群間であればそこに着目した統計値を採用する、という流れになるのではないでしょうか。
# pooneilコメントどうもありがとうございます。
論点はどこでしょうか? Single-unit studyにGLMMは必要ないというご意見でしょうか?
いままでは問題とされてこなかったけど本当は問題なわけです。だから、統計モデルとしてexplicitに扱いましょうよ、という問題提起だと思っていただければと思います。
返答としてはこんなもんなんですが、インスパイヤされたことをエントリにしてみました。そちらもぜひご覧ください。
2009年11月05日
■ A three-choice reaction time task using Arduino
I have been using Facebook for a while. I imported my blog and it works ok. Now I decided that I will occasionally post some entries in English.
Recently I made a simple reaction time task using Arduino. So it will be a good material to write in English because it may interest English-speaking readers.

Click to enlarge
Arduino is a set of hardware (with AVR microcontroller) and software (IDE with a simple, C-like language) for physical computing. It is very cheap ($30 hardware and free software) and is very easy to start. If you have an LED, you can run a program to blink the LED within 5 min.
I have been interested in physical computing. But the PIC, the most popular microcontroller, was too difficult for me, with a complicated, assembler-like language.
Recently I read some issues of MAKE magazine and know what arduino is. I bought Arduino Duemilanove, just as a hobby, and played around for a while. I noticed that I can use it for a simple reaction time task, which would be usually built on DAQ + LabView, Matlab or TEMPO. So I started and this is the preliminary result.
As in the figure, Arduino Duemilanove is connected via an USB cable to my PC. A 006P battery is also connected. It works standalone when detached from the USB cable. All of the other parts are plugged into a solderless breadboard. It has three LEDs. From D9-11 pins, an LED and a resistor (330 Ω) are serially connected to the GND pin. A small speaker is connected between the D3 pin and the GND pin. Three buttons are connected between the A0-2 pin and the GND pin.
Now I start arduino IDE on the PC and upload the program 'Reaction_time2.pde'. here is the code. The code is based on a sample program on the arduino web site (Reaction Time Tester).
Then detach the USB cable. It will run standalone. Start the task by pushing the middle button. Then three LEDs flashes. After a variable delay (defined by iti_min and iti_max), one of the LED turns on. Your task is to push the correct button (left button for left LED and so on) as early as possible. If you choose the correct one, the LED turn off with a high tone. If you choose the incorrect one, the LED does not turn off, with a low tone. After 10 trials, three LEDs flashes again and the task terminates.
After the task, reconnect the USB cable and run the program 'Read_score2.pde', which is actually a subset of the 'Reaction_time2.pde'. Here is the code. Then you will get a result like that on the serial monitor of the IDE:
Begin Ready 0 3 959 1 1 1 939 1 2 1 515 1 3 2 530 1 4 3 914 1 5 1 510 0 5 1 732 1 6 1 507 1 7 3 518 1 8 1 434 1 9 2 422 1 56 Completed
The first column: trial number; the second column: target number; the third column: reaction time in ms; the fourth column: correct (=1) or incorrect (=0).
There are a lot of rooms for improvement but I am happy that it looks good enough for replacing some of otherwise bigger systems.
2009年04月25日
■ ランダムシャッフル関連
これまでpseudorandom sequenceが必要なときにはMATLABでrandpermを使っていたのだけれど、現在使用中の行動制御プログラム(C言語ライクでショボめな言語を持っている)でpseudorandom sequenceを作る必要が出てきたので探してたら、ちょうど結城氏のサイトでそのネタが扱われてました。
もとの話はたとえば1,2,3,,,,,9,10というsequenceがあったときにこれをかき混ぜてシャッフルしてやるときのアルゴリズムの問題です。わたしの場合はたとえば、条件Aと条件Bとがpseudorandomに混ざっているようにしたい、たとえば、n=10trialのブロックで8回条件Aが出て(p=0.8)、2回条件Bが出るようにして、その出現パターンには法則性がないようにしたい、というわけです。ですので、たとえば1,2,3,,,,,9,10の数列をシャッフルして作った数列のうち、1-8を条件Aに割り振って、9-10を条件Bに割り振ってやればいいわけです。
ということで、まずは結城氏のサイトにあった正しい例と良くない例とをMATLABで試しに作って確認してみました。
loopno=1000000; data=1:10; lengthdata=length(data); %--正しくない例----------------- randdata=zeros(loopno,lengthdata);randdata(:)=NaN; tic; for j=1:loopno outdata=data; for i=1:lengthdata % generate random number from [1 lengthdata] r=unidrnd(lengthdata); % swap outdata(i) with outdata(r) temp=outdata(i); outdata(i)=outdata(r); outdata(r)=temp; end randdata(j,:)=outdata; end toc; histdata=zeros(lengthdata,lengthdata);histdata(:)=NaN; for i=1:lengthdata histdata(i,:)=hist(randdata(:,i),data); end %--正しい例----------------- randdata2=zeros(loopno,lengthdata);randdata(:)=NaN; tic; for j=1:loopno outdata=data; for i=1:lengthdata-1 % generate random number from [i+1 lengthdata] r=i-1+unidrnd(lengthdata-i+1); % swap outdata(i) with outdata(r) temp=outdata(i); outdata(i)=outdata(r); outdata(r)=temp; end randdata2(j,:)=outdata; end toc; histdata2=zeros(lengthdata,lengthdata);histdata2(:)=NaN; for i=1:lengthdata histdata2(i,:)=hist(randdata2(:,i),data); end %--カラー表示でまとめ----------------- mag1=min(histdata(:)); mag2=max(histdata(:)); cellmean=sum(histdata(:))/length(histdata(:)); figure subplot(2,1,1) imagesc(histdata, [mag1 mag2]) axis square subplot(2,1,2) imagesc(histdata2,[mag1 mag2]) axis square
2009年03月23日
■ Mutual informationとdecoding
さてさて4日連続投稿、のはずが遅れてしまいましたが前回のつづきです。これまでで最長エントリではないかと思います。
Mutual_informationとdecodingの関係ということがずっとあたまに引っかかってました。ひとつは以前のわたしのエントリ(「「補正」が必要なのは、モデル化が不充分である証拠 」)でmutual informationのupward biasについて書いたときに、最上さんのブログエントリ(「情報量の有用性、補正、ビニング」)でレスポンスをいただいていて、近日中にレスポンスしますと言ったきり放置中だったということがあります。ただそれだけではなくて、BMI関連につなげてencoding-decodingについてもっときっちり考えておきたいと思っていたのですけど、ずっと手つかずのままでいたということもあります。Kay et.alのNature 2008とMiyawaki et.al.のNeuron 2008についてきっちり読んでおきたいのですが、まだそこまでたどり着けてません。vikingさんのところで神谷さんとの非常に興味深いやりとりがあるのですが。
そういうわけで、今回はmutual informationのほうに重きを置いて、いろいろ読んで考えたことをまとめておきたいと思います。最上さんのエントリへの直接的返答というよりはそれに刺激されてエントリを作成したという側面のほうが強いです。直接的なレスポンスに関してはいちばん下で書きます。
んで、以前のわたしのエントリですけど、
超背伸びして書きました。怪しいところをwebで確認したりせずに書いた。もうしらない。厳しくせずに、褒めて伸ばしてほしい。
で、情報理論ってなんか嫌いなんですよね。っていうかニューロンの発火の解析関連での情報理論の応用ってのが嫌いってのが正しいのか。
というかんじで逃げをうちながら極論を言ってたのでかんじが良くなかったなあと思います。そのときはmutual informationそのものの話よりかは「補正」が必要な場合の例のうちのひとつとして挙げたつもりでいました。、昔勉強したとき("Spikes" MIT pressが出版された時代)の記憶をたどりながらなんでかなりあやふやな話でしたが。ただ、せっかくレスポンスがいただけたので、いい機会ですからmutual_informationのupward biasの問題について現状に追いついてみようと思います。というわけでいくつか論文を読んでまとめてみます。まずはこちらから:"Correcting for the Sampling Bias Problem in Spike Train Information Measures" Stefano Panzeri, Riccardo Senatore, Marcelo A. Montemurro, and Rasmus S. Petersen. J Neurophysiol 98: 1064-1072, 2007
まずは導入ですが、刺激sと応答rとのあいだのmutual information MI(s,r)は二つのエントロピーの引き算:MI(s,r) = H(r) - H(r|s)で計算できます(direct method)。それぞれのエントロピーはresponse entropy: H(r) = Σp(r)*log2(p(r))、noise entropy H(r|s) = ΣΣp(s)*p(r|s)*log2(p(r|s))として計算されます。Mutual informationのバイアスとは、けっきょくのところその元となるエントロピーのdownward biasを反映しています。つまり、試行数が少ないとき(rの場合の数K << 試行数Nを満たさないとき)、rの分布が正確に推定できないため、エントロピーが低く計算されてしまう。しかも、この効果はnoise entropyのほうが大きいから(試行数が少ないから)、mutual information全体としてはupward biasとなる、というわけです。
それでこれを補正するために使われるのがMiller-Madow補正(1955)というやつで、これをニューロンのスパイクの例に応用したのがPanzeri and Treves (1996)でした。これが最上さんが説明してくださった、p*log(p)をテーラー展開をして二次の項までを見たときの近似値の話でした。ちなみにPanzeri and Treves (1996)でベイズ推定をしているのはこの補正でのbin数(R, R_s)についてですので、わたしがイメージしていたベイズ的なmutual informationの推定というのとは少々違っているようです。
わたしがイメージしていたような、trial数が充分あることを仮定していないときのmutual informationの推定というものは
- Wolpert DH,Wolf DR (1995) "Estimating functions of probability distributions from a finite set of samples."(pdf) Physical Review E 52: 6841-6854.
- Ilya Nemenman, William Bialek, and Rob de Ruyter van Steveninck (2004) "Entropy and information in neural spike trains: Progress on the sampling problem"(pdf) Physical Review E 69, 056111
あたりのことだったようです。ここで前回の比率のベイズ推定のエントリの応用問題ですが、応答rはK binあるので多項分布です。多項分布の自然共役分布はDirichlet分布となりますので、これをpriorとおいてベイズ推定をします。あるk binでのスパイク生成率の最尤推定値がy_k/n (実データでnスパイクのうちy_kスパイクがk binに落ちた場合)だったとすると、priorをa_k/A (k binのcountをa_kとして、A = Σa_k)とおくと、あるk binでのスパイク生成率のMAP推定値は(y_k + a_k)/(n + A)となります。でもってnoninformative priorだとa_k = A/K (K=bin数)となるというわけです。(このへんはHausser and Strimmer "Entropy inference and the James-Stein estimator, with application to nonlinear gene association networks"(pdf) の説明を使いました。)
後者のNSB entropy estimation methodは、これをさらに改良して複数のDirichlet分布の重み付けみたいにしてやると推定が良くなることを見つけた、という話でした。これは元のPanzeri and Treves (1996)よりもbiasが少ないということで優れています(Figure 3A,B)。たしかに、良くも悪くもpriorの選択の善し悪しが推測の成績に大きく影響を与えるようです。
このような状況でPanzeri and Trevesが巻き返しを図ったのが、
Montemurro, Senatore and Panzeri "Tight Data-Robust Bounds to Mutual Information Combining Shuffling andModel Selection Techniques" Neural Computation 19, 2913-2957 (2007)
であるようです。この論文はnoise correlationのうちでcorrelationのない成分H_ind(r|s)を計算したり、刺激と応答をシャッフルしたH_sh(r|s)を使ってやると、NSB methodよりも優れた成績のときがある(Figure 3C, D)というものなのですが、なんか部分的な改善というかんじでわたしにはあまりピンと来ませんでした。
ともあれ、これがだいたいPanzeri et.al., JNP 2007の内容でした。
さてさてまだつづきます。直接バイアス問題に関わるわけでもないのですが、entropyおよびmutual informationの信頼区間のベイズ推定をしたという仕事があります:
- Kennel, M., Shlens, J., Abarbanel, H. D. I., Chichilnisky, E. J. (2005). "Estimating entropy rates with Bayesian confidence intervals." Neural Computation, 7, 1531-1576.
- Jonathon Shlens, Matthew B. Kennel, Henry D. I. Abarbanel, E. J. Chichilnisky. (2007). "Estimating Information Rates with Confidence Intervals in Neural Spike Trains." Neural Computation 19:7, 1683-1719.
前回のエントリで比率の推定についてMLEおよびベイズ推定によって推定のばらつき具合を評価して信頼区間を求めましたが、エントロピーも有限個のデータがどのbinに落ちるかという確率から推定されるものですので、同様なアプローチが可能です。エントロピーおよびmutual informationについて、多項分布のMLEから信頼区間を計算したもの、さらにMiller-Madow補正をしたもの(信頼区間には変化無し)、ベイズ推定で信頼区間を計算したものを提示しています。
Mutual informationも比率のデータと同様、信頼区間が付いたものとして扱うのが筋だと思いますので、こういう形で統計に乗っけてやるのがいいんじゃないかと思いました。そもそもmutual informationで出てくる数字のバイアスを問題にしていたのは、いったんinformation rateが計算された後にそれが確定した値として統計で扱われるというあたりに違和感を持っていたからだったわけです。(ROCがひとつの確定した値として一人歩きするのと同じ。) 思えば統計的な扱いとmutual informationとを対立させるというよりは、mutual informationという量を統計的に扱ってやるというのが正しいアプローチなのかもしれません。
まだつづきます。わたしがこのへんの論文を読んでいた動機の一つは、さいきんはやりのencoding-decodingのスキームと、mutual informationを用いた解析とはどういう関係にあるかという点でした。ちょっと話は飛びますが、encoding-decodingの話のほうでさいきん読んだレビューについて言及します:
Wu MC, David SV, Gallant JL. "Complete functional characterization of sensory neurons by system identification."(pdf) Annu Rev Neurosci. 2006;29:477-505.
これはvisual neuroscienceの分野で使われているreverse correlationとかspike triggered averageとかm-sequenceを使ったsystem identificationとかSVMを使った推定といった、刺激sからニューロンの応答rへの変換r=f(s)のfを推定するすべての研究についてMAP推定の一種として捉えてまとめたレビューです。MLEもpriorを使わないMAP推定の一種として捉えられるし、parametricなWiener-Volterra展開的なアプローチも、SVMを含んだカーネル法によるnonparametricなアプローチもモデル選択の違いとして統一的に捉えられることを示しています。このようにしてモデルを作成して、MAP推定(点推定)をしてやって、corss validationをしてやってモデルの善し悪しを評価する、というスキームが示されます。
このコンテクストの中では、mutual informationはモデル選択の善し悪しの評価法としてcorrelation coefficientなどと並列して扱われています(correlation coefficientと比べてmutual informationの方がよいかどうかはわからない、みたいな記載があります)。たしかにそうして考えてみると、mutual informationというのは統計解析の中でいうR^2に対応したものと考えることが出来るのではないでしょうか。データが有限ならupward biasを持っているという意味でも似ています。そういう意味では、mutual informationのオリジナルな意味、データの伝達の正確さの評価に戻って、mutual informationの定量化がなにをしているかということを考える意義があります。
さてこのようなencoding-decodingのスキームと、mutual informationを用いた解析とはどういう関係にあるか。その意味でJonathan D. Victorのこれらの論文に書いてあることは非常に役に立ちました。
Jonathan D. Victor. 2006. "Approaches to Information-Theoretic Analysis of Neural ActivityApproaches to Information-Theoretic Analysis of Neural Activity." Biological Theory 1:3, 302-316.
Jonathan D. Victor, Sheila Nirenberg "Indices for Testing Neural Codes" Neural Computation, December 2008, Vol. 20, No. 12, Pages 2895-2936
前者にはShannon entropyでできることの限界を強調してます。後者では情報理論的解析とベイズ推定とを並列的に扱って、どちらを使うのが刺激sから応答rへのcodingの問題(あるコーディングが使われている可能性を除外する方法)に向いているかの異論をしています。前者の論文にはこう書いてあります:
Fundamentally, the Shannon theory was designed for characterizing communication systems whose principles were understood, not for the "inverse problem" of determining the principles by which a system works from observations of its behavior.
それからこちら:
Moreover, the Shannon theory does not attempt to describe the relationship between a sensory or motor domain and neural activity (i.e., the nature of the neural representation) but merely provides an index of how faithful this representation is.
そういう意味では、direct methodのいいところは、刺激sから応答rへの変換に関して明示的にモデルを作らなくてもmutual informationを計算することによって、encodingの過程についてなんらか示唆を得ることができるという点にあるのでしょう。ただし、まったくassumption-freeなわけではなくて、応答rのbinの切り方とか、時間幅の問題とか、そういうところで隠れてcodingのモデルが入り込んでいるんじゃないかと思うのです。長々と書きましたが私の主張はなにかといいますと、刺激sと応答rとはものすごく違った構造をしているわけでして、その二つをつなげるmutual information MI(s,r)を作るために、非明示的にモデルのassumptionが入っているので、direct methodというものはなんか変なんじゃないのか、ということです。
Reconstruction methodのほうはmutual informationの計算という意味ではずっとクリーンです。MI(s,r)の代わりに応答rからdecodeした刺激s_estを使ったMI(s, s_est)を計算するわけです。刺激は自分でデザインできるからbinとかの問題もクリヤーだし、なによりsと s_estという、同じ構造のものを比較しているわけですから。もちろん、MI(s, s_est)はモデル化が悪ければいくらでも下がるので、lower boundしか決めることができない( MI(s, s_est)<=MI(s,r) )わけですが、reconstruction methodははモデル化の部分を上記のMAP推定的なスキームに任せて、mutual informationをモデルの評価に限局して使うということで、こっちだけにしといた方が良いんではないだろうか、という気がしてきたのです。
まあ極論ですが。実際には、刺激空間がよくわかっている低次視覚野ならばモデル化のほうからアプローチする方が効率がよいし、高次視覚野ではなるたけ最適刺激にassumptionをおかずに多くの種類の刺激を使って応答を見て、それを比較的応答モデルに対するassumptionの少ないmutual informationで評価してcodingについての示唆を得る、というのが現実的であるということなのでしょう。
さてここまで書いたところでPanzeriがQuirogaといっしょに書いたレビューが出ました。これについても読んでおきましょう:
Rodrigo Quian Quiroga and Stefano Panzeri "Extracting information from neuronal populations: information theory and decoding approaches"(pdf) Nature Reviews Neuroscience 10, 173-185 (March 2009)
現在の文脈で重要なのは、"Complementarities of decoding and information theory"のセクションなのですが、decodingはposteriorのarg maxをとっている(MAP推定をしていちばんもっともらしい刺激を推定する)のに対して、mutual informationはそれ以外にも情報があることを定量化している、という点を強調します。だから、mutual informationでは2番目にlikelyな刺激についての情報とか、ある図形が非常にunlikelyであることとかも持っているだろう、というわけです。
ただこれって、あくまでMAP推定に限局した問題だと思うんですけどね。posteirorの情報を持っているということは、確率密度分布を持っているわけだから、刺激26がmost likelyだけど、刺激29もそのつぎにlikelyだとかそういうことはdecodingの方でも言えるわけですよね。だからあまり説得的に思えないのですが。
どうやってdecodingとmutual informationを組み合わせるかという点では、decodingできたinformation MI(s, s_est)をdirect methodによるMI(s,r)との比率で評価する(「95%の情報がdecodeできている」といった評価ができる)ということが書いてあって、これはいいなと思いました。つまり、BOX3に書いてあることですが、われわれがdirect methodで計算しているmutual informationというのは応答rそのものではなくて、rをカテゴリー分けしたりいくつかの処理をしたf(r)なわけです。(さっきわたしはこの点を捉えて、非明示的なモデリングが入っている、と指摘したわけですが。) すると、
MI(s,f(r)) < MI(s,r)
が成り立ちます。そのうえで、reconstruction methodで使われるdecodingでも、けっきょくまったく同じ形でのrの変換(r -> f(r))が不可避なわけです(spike countのカテゴリー化であれ、時間windowの幅であれ)。だからdecodingによる逆変換をg()としておくと、reconstruction methodによって計算できるmutual information MI(s, s_est)では、
MI(s, s_est) = MI(s, g(f(r)) <= MI(s,f(r)) < MI(s,r) MI(reconstruction method) <= MI(direct method)
がなりたちます。Upward biasの問題は依然つきまといますが、そういう意味では、decodingのperformanceの評価としてはいいんではないかと思います。この、direct methodとreconstruction methodで挟むという話自体は新しいものではなくて、私自身は以下のような論文で見ましたが、もっと広く使われてもいいと思いました。
- Borst A, Theunissen FE. "Information theory and neural coding." Nat Neurosci. 1999 Nov;2(11):947-57
- Buracas GT, Zador AM, DeWeese MR, Albright TD. "Efficient discrimination of temporal patterns by motion-sensitive neurons in primate visual cortex." Neuron. 1998 May;20(5):959-69.
あとはdecodingの成績をまとめたconfusion matrixを元にしてmutual informationでの「次元の呪い」を除けないか、みたいな話も。これもdecodingとmutual informationとを組み合わせる方法として有望だと思いました。
イントロの部分でdecodingとmutual informationがどんな風に"intrinsically related"であるかを示すことを目指す、ってあったんですが、ここについてはあまり満足がいくかんじがしません。
思うんですけど、どんな風に"intrinsically related"であるかって、ベイズ推定はp(s|r) ∝ p(r|s) * p(s)を使っていて、mutual informationはp(r)とp(r|s)を使っていて、共通のものを使って計算しているという意味でintrinsically relatedなわけですよね。それなら、ベイズ推定の計算から推定されたp(r)とp(r|s)でもってmutual informationを計算する、みたいなことをすれば良いんではないのでしょうか。p(r)とかは積分しないといけないわけですけど。ベイズ推定の方はcross validationが使えるけど、mutual informationのほうはそれに対応したものがない(shuffleしたもの?)というあたりが推定の問題でもあるわけだから、同じ確率密度分布を共有して計算することで推定の確かさも揃えて扱うことができるんじゃないでしょうか。ちょっともう素人がわかったような口をきいてるかんじのもの言いになってる気がするんで恐縮ではあるのですが。
さて、いろいろ書きましたが、もちろん統計解析と情報理論は深いところで繋がりあっているので、そのへんをもう少し勉強しなくちゃなあと思ってます。その意味では情報幾何での、mutual information - fisher information - KL divergence - maximum likelihood estimationあたりを包括して捉えられる図式を勉強しなきゃなあ、ということで「新版 情報理論の基礎」村田昇を読むのを自分の宿題にしてます。
ふと思い立って村田昇教授のサイトを見たら「少数データを用いた推定」というのを発見。KL情報量よりrobustなBregman情報量というものがあるらしい。きりがないのでこのへんまでとします。
さて、以上を踏まえて、最上さんのブログエントリ(「情報量の有用性、補正、ビニング」)に応答してみたいのですが、補正公式、ベイズ統計関連、binningなどについては以上ですでに言及しました。脳が尤度推定やベイズ推定をしてるという最近の話題には私も興味があるし、とくに意見の相違はないですね。この話題と解析としてのベイズ推定とを混同しないほうがよい、というのはたしかにそうだと思いました。ただ、これも深いところで通底しているような気がしてますが。
ベイズ統計の是非の話というのはじつは二層あって、[頻度主義 vs. ベイズ主義(モデルパラメータthetaを固定しているとするか、確率変数としてとらえるか)]という話と[priorを使わない(MLEまで) vs. 使う(MAP推定、ベイズ推定まで)]という話があるのではないかと思います。今回の話は後者でした。脳が確率密度分布を持っているってのはどちらかというと前者の話だったのではないかと思います。
後者の問題に関してですが、前回のエントリを作りながら思ったのですが、あるていど単純な問題にはベイズ統計はいらないし、それなりに複雑な問題で、priorを使わないと明らかに損しているようなモデルではpriorを使うことによって道が開けるわけです。それぞれの実験の状況で、利用可能なデータ量(maximum likelihood)と、利用可能なpriorとによって、どっちがより効率のよいモデルを作れるかによって決まるんではないかと思います。充分データが集めることができて、それで充分なforward modelを作ることができるならMLEのほうがよいし(priorを使うことによる不安定性を回避できる)、データ収得に限りがあるようなら、priorを使ったほうが推定の成績と効率は良くなる。でもって、脳計測の場合はしばしばデータ収得に限りがある状況のほうが多いのではないでしょうか。(「物理をやっていた者の思考として自然」というのは統計物理のように前者が妥当な状況でのことではないでしょうか。)
あと、ぶっちゃけ情報量の利用はプラグマティックな意味でも役に立ちます。
これ以降の部分での複数ニューロンのデータの集積に関する話は、データいじっている方の実感なので面白いと思いました。じっさい、わたしも解析してていつも悩ましい問題です。ただ、別のニューロンのmutual informationを足すというのはその意味上よいのでしょうか? つまりmutual informationの単純な足し算というのは、別々のニューロンが独立した情報を持っているときの情報量を意味しているわけで、そうすると、意味がありすぎると思うのです。一般的なsingle-unit recordingのデータで同時記録をしていないデータを集める場合、多ニューロンのmutual information量の集積は、それらの複数のニューロンのデータから抽出可能な(提示した図形に関する)情報量を計算することとして捉えることができます。そうすると、複数のニューロンのデータを足していけば、似た反応特性を持っている分、得られる情報量の増加は目減りしていくわけで、単純な足し算をするよりかはこういった反応選択性の相関を考慮した集積をしたほうがよいのではないでしょうか。ちょっと発想がdecodingに寄りすぎているのかもしれませんが。
あと、上記のPanzeri et.al., JNP 2007を読んだかぎり、direct methodによって、biasの問題に対処して意味のあるmutual informationを計算するためには、NSB methodのような現状でいちばん良いものですら N_s > 2-4 R (R: 全応答の場合の数)という縛りがあるわけです。そうすると、たとえば5ms windowで0/1 spikeに分類してtime window 8個分でR=2^8=256としても、N_s (刺激ごとの試行数)に500trial以上必要なわけですから、RSVP使うとかしてN_sを相当稼がないといけないわけです。いったんこういう数字を見てしまうと、direct methodを使ったmutual informationの計算というのはかなり敷居が高いんではないだろうか、と思いました。
以上です。長大になりすぎましたが、こうやって書いていくことでまえに考えていたときよりはずいぶんいろいろ明確になってきました。レスポンスしてくださった最上さんにお礼申し上げます。
# viking
僕のblog(しかもモメたエントリ)にリンクをいただいて大変有難いのですが、今回の件とfMRI mind-reading studyの件ってそんなに関連していますか? 工学系出身の身としては、fMRIでmind-readingというのはもっと泥臭い部分(良くも悪くも力業)の話だと思っておりますもので・・・あくまでも比較の問題ですが、むしろERP/MEGの逆問題推定の方が話題としては親和性が高いかもしれませんよ。
# pooneilどうもおひさしぶりです。前者が「力業」に帰結する話なのかどうかはまだ私は勉強不足なのでよくわかりません。そこへ辿りつくための枠組みあたりをまとめているつもりです。ほんとうは立場上、そんなのんきなこと言っているようではいかんのですが。後者が親和性が高いことは二百も承知でして、それが以前にLFPのまとめを作ったりしたことと関係してたりします。
# vikingmind-reading methodのチュートリアル的な総説がNeuroImageに出てます。ご参考までに。
Machine learning classifiers and fMRI: a tutorial overview (Pereira F, Mitchell T, Botvinick M, Neuroimage. 2009 Mar;45(1 Suppl):S199-209)
http://dx.doi.org/10.1016/j.neuroimage.2008.11.007
2009年03月19日
■ 比率のデータにエラーバーを付けたいんだけど
最高ですか! (<-奇をてらったごあいさつ)
どうもご無沙汰しております。ここに書いているということは暇だということを必ずしも意味しないけれど、ここに書いてないということはかならず忙しい。んで、しばらく頭の中をぐるぐる回っていたものがあったので、そのへんを吐きだしてすっきりさせて論文書きに集中したいと思いますので、いろんなところに不義理を働きつつもこんなもの書かせていただきます(超低姿勢)。
...なんて前口上をつけた原稿を作ってあったんだけど、なぜか久しぶりの三日連続投稿に。
んで、いきなり口調が変わるのですが、比率のデータにエラーバーを付けたくならないっすか?なんでもいいんだけど、たとえば二択の課題の成績が試行数120で正答数73で正答率61%だったとします。これは二項検定によって両側検定5%水準でchance level (50%)よりは有意に大きいです(図1の(1))。それを直感的に示すようなエラーバーをバーチャートに付けたい。これが今回の問題です。
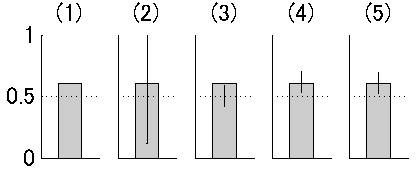
図1
普通は比率のデータにエラーバーなんて付けません。73/120という一つのデータだと思われているからでしょうね。でももしこれが正答数でなくて、反応潜時のデータかなんかだったらもちろんSDを付けることができます。同じように、73/120というデータのばらつき具合にエラーバーを付けてやりたいと思うのですよ。
まずは、間違った方法ではありますが、正答に1、誤答に0を割り振って、たんにそのSDを計算します。するとmean = 0.61, std = 0.49という値が出てきます(図1の(2))。もちろんこれは誤差の分布を正規分布と仮定しているのが間違いなわけです。(これをgeneralized linear modelでリンク関数をbinomialにして計算すれば間違いにはなりません。これは最尤推定でやっていることと同じのはずです。これについては後述。)
それでは今度は、比率の検定に基づいて考えてみましょう。ここでは二項検定で検定をしているわけですからそれを反映させましょう。つまり、試行数120で帰無仮説H_0:母比率p=0.50の場合の二項分布Bi(n,p) = Bi(120, 0.50)を計算します。Rだとこんなかんじ:
total <- 120 correct <- 73 x <- seq(0, total) bino_prob1 <- dbinom(x, total, 0.50) plot(x,bino_prob1)
図2
両側検定5%有意水準で正答数70以上もしくは48以下だと有意です(図2の(1))。今回は73だから有意でした。この範囲を図に無理矢理入れるならこんな図(図1の(3))になりますが、こんなの誰も望んでない。だいたい試行数にしか依存してないし。
同じ発想で母比率が73/120だったときに実データがどのくらい散らばるかということを二項分布Bi(120, 73/120)
total <- 120 correct <- 73 x <- seq(0, total) bino_prob2 <- dbinom(x, total, correct/total) plot(x,bino_prob2)
で評価してやることも出来ます(図2の(2))。こうするとエラーバーの範囲は[0.5167 0.6833]となります(図1の(4))。これはもっともらしいですが、統計的な裏付けに欠けます。
さてこのへんからが本題です。Maximum likelihood estimationを考えてみましょう。データ73/120は既知で確率theta = 0-1のほうが変数で、尤度関数は
likelihood ∝ theta^correct * (1-theta)^(total-correct)
となります。
theta <- seq(0,1,1/total) likelihood <- theta^correct * (1-theta)^(total-correct) Lc <- likelihood / sum(likelihood) plot(theta,Lc)
このピーク値を見るとたしかに0.61になっていて、correct/totalがthetaの最尤推定値になっていることが確認できます(図2の(3))。またさらにこの尤度関数のcumulative density functionを作ってやると(図2の(4))、最尤推定値の95%信頼区間を[0.519 0.691]というふうに評価することが出来ます(図1の(5))。
この値は図1の(4)とほとんど変わりません。両者の違いは、二項分布では横軸xがデータの個数で離散的になっているのに対して、最尤推定では横軸thetaが連続値になっているという点ですが、thetaのほうを離散的にして計算すると(theta = [0:1/120:1]としてやる)微妙に違う値になります(図3の上 緑が二項分布、青が尤度関数、図3の下 緑が尤度関数-二項分布の引き算)。数学的にはべつものなんでしょう。つまり、二項分布Bi(n,r,p)のグラフを縦に見るか横に見るか、という話になります。くわしくは「ベイズ統計学は本当に有効か?」赤嶺達郎および「枠どり法とPetersen 法の区間推定における伝統的統計学とベイズ統計学との比較」 赤嶺達郎(pdf)(今回の話のネタ元のひとつ)を参照、ということで。
Webを探したら「検定と区間推定」奥村晴彦(TeX本で有名な方)にRでのimplemantationがありました。なんかこれを読むといま書いたことのすべてが書かれているような気がしますが、まあこうやって書いてみることは悪くないことで。
> binom.test(73,120,0.5)
Exact binomial test
data: 73 and 120
number of successes = 73, number of trials = 120, p-value = 0.02208
alternative hypothesis: true probability of success is not equal to 0.5
95 percent confidence interval:
0.5150437 0.6961433
sample estimates:
probability of success
0.6083333
ともあれ最尤推定まで考えると、それなりに意味のあるエラーバーが書けそうです。役に立つと思うんですけど、なんでみんな使わないんですかね。
さて、こんなことを計算している目的の一つはベイズ統計のお稽古だったわけで、「ベイズ統計学入門」渡部洋著を元にしてさらに先に進みます。
図3
Maximum likelihood estimationで得た最尤推定値はベイズ統計の立場から見れば、無情報のprior (p(theta)が一定)を用いたときのMAP推定値と等価です。つまり、さっきの尤度関数(図2の(3))はposteriorの分布と捉えることが出来て、点推定をするとピーク値の0.61を取ることになり、区間推定をするなら図2の(4)のように信頼区間を計算することが出来ます。
ベイズ統計の場合には、信頼区間(confidence interval)ではなくて、信用区間(credible interval)として、全体の10%の面積が除外されるような縦軸の値を設定して計算します(図2の(3))。これはMAP推定自体がピーク値(mode)を使っていて、median (=50% percentile)ではないということからすれば納得がいきます。こうして計算された信用区間は[0.520 0.692]となります。これはさっきの信頼区間の値[0.519 0.691]とほとんど変わりません。分布が歪んでないのでそりゃそうでしょうね。
教科書まる写し口調ですが、今まで扱ってきた二項分布をBi(total, correct/total)と表現しておくと、この尤度関数L(=ベイズ統計でのposterior)はベータ分布Be(correct+1, total-correct+1)と表現できます。二項分布のベイズ推測での自然共役事前分布がベータ分布であることがわかっています。つまり、priorがベータ分布なら、posterior ∝ likelihood * priorもベータ分布ということ。
無情報のpriorを使う場合にはこれはBe(1,1)と書けて、これはpriorではtotal=0, correct=0となっていることと等価です。このときのposteriorは
posterior ∝ likelihood * prior = Be(correct+1, total-correct+1) * Be(1,1) = Be(correct+1, total-correct+1)
となるわけです。確認のためこのベータ分布をplotしてみますと、これまでの尤度関数とまったく一致します(図3の上 赤がベータ分布だが青に重なって見えない 図3の下 赤が尤度関数-ベータ分布の引き算)。
面白いのは、ベイズ推定の式がたんにベータ分布の中身Be(a, b)の足し算になっている点ですね。もしここでpriorとして無情報のprior(Be(1,1): total=0, correct=0)を使わずに、局所一様分布のpriorというやつを使うとBe(1/2, 1/2)となります。これはtotal=-1, correct=-1/2としているのと同じということで、これは得られたデータから1 trial分差っ引いてposteriorを作っているということになります。おお! ここでも出てきた補正の思想!
というわけでpriorとしてはBe(1,1)とかBe(1/2,1/2)とかがありうるのだけれど、はっきり言ってこのような単純な状況ではベイズ統計が出てくる必要性はあまりないと言えましょう。ではどういうときに必要かというと、複数のセッションを足し合わせるような状況ですね。(このへんから北大の久保拓弥先生のweb資料(pdf)に依拠しながら進みます。)
これまで使ってきた73/120というデータですが、じつはこれが5回分のセッションの集計だったとします。たとえば、{16/30, 24/30, 12/20, 11/20, 10/20}というデータだったとします。この5点をプロットしてそれのSDでエラーバーを付ける、というやりかたがありますが、これは試行数の120を使ってないので正当な評価とは言えないでしょう。(セッションごとのばらつきを評価しているというのはわかるのだけれど、それが0.5からdeviateしているかどうかを検定するとしたらそれは意味がない。)
データのモデル化を考えてみると、各セッションごとに二項分布Bi(n, 73/120)から出てきたと考えるとします。これはセッションごとにthetaが一定(=73/120)であるとするモデルですが、二択の心理実験とかではだいたい当てはまらないわけです(これはoverdispersionが起こっているかどうかで検証できる)。そこでセッションごとのthetaが73/120からばらつく(とりあえず正規分布でSD=0.1として生成してやると{0.52 0.80 0.60 0.55 0.51, ....}となる)というふうにするのがrandom effectモデルで、このSDを決めてやるために経験ベイズ、階層ベイズが出てきて、そうするとセッションごとのばらつきとセッション全部のばらつきとを評価することが出来て、エラーバーとしてもそれを示すのが正しいのだと思うのだけれど、おっとここで時間のようです(<-逃げた)。
いったん逃げたけど帰ってきた。やりかけなんだけどRでの解析を書いておくと、まずデータ作り:
id <- factor(c(rep(1,30), rep(2,30), rep(3,20), rep(4,20), rep(5,20))) y <- c(rep(1,16), rep(0,14), rep(1,24), rep(0,6), rep(1,12), rep(0,8), rep(1,11), rep(0,9), rep(1,10), rep(0,10)) data3 <- data.frame(y=y, id=id)
これをidを無視してセッションをまとめたときの解析は、generalized linear modelでリンク関数をbinomialにしたもので、こんなかんじ:
result1 <- glm(y ~ 1, family = binomial, data = data3) summary(result1)
結果は
> summary(result1)
Call:
glm(formula = y ~ 1, family = binomial, data = data3)
Deviance Residuals:
Min 1Q Median 3Q Max
-1.369 -1.369 0.997 0.997 0.997
Coefficients:
Estimate Std. Error z value Pr(>|z|)
(Intercept) 0.4403 0.1870 2.354 0.0186 *
---
Signif. codes: 0 ‘***’ 0.001 ‘**’ 0.01 ‘*’ 0.05 ‘.’ 0.1 ‘ ’ 1
(Dispersion parameter for binomial family taken to be 1)
Null deviance: 160.68 on 119 degrees of freedom
Residual deviance: 160.68 on 119 degrees of freedom
AIC: 162.68
Number of Fisher Scoring iterations: 4
となっていて、Interceptのestimateは0.4403で、standard errorは0.1870。だから比率の最尤推定値および95%信頼区間は
> 1 / (1 + exp(-0.4403)) [1] 0.6083305 > 1 / (1 + exp(-0.4403+0.1870*1.96)) [1] 0.5184366 > 1 / (1 + exp(-0.4403-0.1870*1.96)) [1] 0.6914315
というふうに正しく計算できます。
また、セッションごとにIDが付いたデータの場合はGLMM(混合モデル)を使って、さっきのdata3を解析すると、
library(glmmML) result3 <- glmmML(y ~ 1, family = binomial, cluster = id, data = data3) summary(result3)
というので計算が出来ます。この結果は
> summary(result3)
Call: glmmML(formula = y ~ 1, family = binomial, data = data3, cluster = id)
coef se(coef) z Pr(>|z|)
(Intercept) 0.4337 0.2269 1.912 0.0559
Scale parameter in mixing distribution: 0.2777 gaussian
Std. Error: 0.2753
Residual deviance: 160.2 on 118 degrees of freedom AIC: 164.2
となって、idのrandom effectのSEが0.2753と出てきます。このため、interceptの推定もglmから多少ずれるので、比率の推定も以下のように多少低めに出ていて、信頼区間も広くなっています。ひとつ突出している24/30が効いている分を考慮しているかんじがうかがえます。
> 1 / (1 + exp(-0.4337)) [1] 0.6067568 > 1 / (1 + exp(-0.4337+0.2269*1.96)) [1] 0.497244 > 1 / (1 + exp(-0.4337-0.2269*1.96)) [1] 0.7064955
ただ、glmmMLの結果の見方がまだよくわかってないので("Scale parameter in mixing distribution: 0.2777 gaussian" ってなんでしょう。以前の資料を見るとここはSDが表示されていたんですけど。)、このへんで止まってます。
Chance levelから有意かどうか、というのはgeneralized linear modelまで来ると、H_0のrejectという形ではなくてモデル選択ということになるので、inteceptが0 (比率が0.50)のモデルと最尤推定で得られたinterceptでのモデルとでAICを比較するということになるんでしょう。論文書きのときはpが付けられないと困っちゃうんだけど。
さてさて、ここからが本番です(本気)。長々と引っ張ってきましたが、このエントリの最終目的はじつはmutual informationのバイアス問題に関連しているのでした。有限の試行のデータから刺激sと応答rとの間からmutual informationを計算するときに必要なのはp*log2(p)の計算なわけですが(応答rがK binあるうちのi binに落ちたときの確率をpとする)、このpの最尤推定値として、あるbinに落ちたspike数yとその総数nを実測データから計算してつくったy/nを用いているわけです。このことはエントロピー推定のバイアスを生むわけですが、そもそもcountデータからの比率の推定に基づいているのだから、今回扱ったような手法でエントロピーおよびmutual informationでもベイズ推定と信頼区間を計算してやることが可能となります。
基本に戻ると、刺激sと応答rとのあいだのmutual information MI(s,r)は二つのエントロピーの引き算:MI(s,r) = H(r) - H(r|s)で計算できます(direct method)。それぞれのエントロピーはresponse entropy: H(r) = Σp(r)*log2(p(r))、noise entropy H(r|s) = ΣΣp(s)*p(r|s)*log2(p(r|s))として計算されます。
ここで比率のベイズ推定のエントリの応用問題ですが、応答rはK binあるので多項分布です。多項分布の自然共役分布はDirichlet分布となりますので、これをpriorとおいてベイズ推定をします。あるk binでのスパイク生成率の最尤推定値がy_k/n (実データでnスパイクのうちy_kスパイクがk binに落ちた場合)だったとすると、priorをa_k/A (k binのcountをa_kとして、A = Σa_k)とおくと、あるk binでのスパイク生成率のMAP推定値は(y_k + a_k)/(n + A)となります。でもってnoninformative priorだとa_k = A/K (K=bin数)となるというわけです。(このへんはHausser and Strimmer "Entropy inference and the James-Stein estimator, with application to nonlinear gene association networks"(pdf) の説明を使いました。)
というわけでこれも手計算してみようかと思いましたが、bin数Kが大きいとあっという間に次元が増えてたいへんなので止めとしておきます。ともあれ、mutual informationでもベイズ推定して信頼区間を付けられるというわけです。今日はこんなところまで。あしたにつづきます。
図4
追記:コイダーマンさんからコメントいただきました。どうもありがとうございます。なるほど、図3での緑(二項分布)と青(尤度関数)の違いはcorrect=20; total=20の場合を考えるとより明確になりますね。さっそく図を作ってみました(図4)。二項分布だとcorrect ratio 100%しかありえなくなってしまうけど、尤度関数だとcorrect ratio 90-95%あたりの可能性もそれなりにあるということがこの図だとよくわかります。
# コイダーマン
こんちは。
私も行動実験や心理実験をやってて同じことを悩んでましたヨ!
自分が読めるようなわかりやすい本には図1(4)の説明しか載ってなくて、
でもその方法だと、たとえば正解率が100%(20回中20回とか)のときに
信頼区間がゼロになっちゃうんですよね。
でも、そんなわけない!
真の確率は98%なんだけど、たまたま結果が100%になったってことだって起きるはずだ!
当時は尤度の読み方どころか文字すらも知らなかったので
自分で一から考えて答えにたどり着いたときは感動したです・・車輪の再発明と言うんでしょうか。
適当に二項分布の母確率pを仮定して、
そのときに実験データ(たとえば20回中20回)が生じる確率を求める。
図の横軸を仮定した母確率のpにとって、縦軸を実験データの発生確率でグラフを書くと・・
母確率pは0~1まで均等にあると仮定して、そのpが選ばれるのをΔpと考えると
・・あ、信頼区間が計算できちゃった。やった!って。
Mutual Infomationの信頼区間計算ってのはいいですね!
これまで私はあまり考えずにブートストラップのような方法で求めましたです。
平均発火頻度をポワソン分布でフィッティングして、
試行回数分のデータを擬似発生させて、自分がきめたBINで相互情報量を計算。
これを1000回とか繰り返したら相互情報量のばらつきがわかるかも、とか。
でもこれじゃ
ある刺激への応答が全部0spk/sだったときに信頼区間がゼロになって困る。
繰り替えしが5回とか少なかったりすると、真の平均発火頻度が0以上である可能性は相当高いのに。
続編期待してます。
# コイダーマンあ・・書き込みした後に思い出した・・
二項分布の信頼区間を求めようとしたのは、実験の解析が目的じゃなかった!。ゲームRagnarokOnlineで錬成だったか何かの成功確率が、いくつかの条件間で差があるのかないのかを匿名某で議論してたときのことだったワ!
心理実験やってたから、なんてウソウソでした!
コイダーマン、どうもありがとうございます。(言いにくいのでさん付け省略で。)
なるほど、20/20の例を使えば、図3上の二つのプロットの違いがより明確に示せますね。追記しておきます。
>RagnarokOnlineで
それ正直に申告しなくてもいいよ!!!
つーかそれアカデミックな議論ですね。
わたしも某巨大掲示板で複数のランキングの相関係数を計算して議論したことがあるけど。
2009年01月13日
■ DKL色空間についてまとめ
さてさて今度はDKL色空間についていろいろ調べる必要が出てきたので、まとめを作りました。数式がたくさんあるので、エントリでは簡略化してます。くわしくはPDFファイルをどうぞ。こちらから:dkl3.pdf
[要旨]
1) DKL色空間とは、色空間を網膜以降の色処理の三つのチャネル(Luminance, L-M, S-(L+M))の軸によって表現したものです。モニタのRGB値からDKL空間への変換を簡単に説明します。2) Brainard 1996に準拠して、LMS色空間からDKL色空間への変換行列を導出します。3) さいごに、DKL色空間が持つややこしい点について言及します。
[さまざまな色空間]
網膜には三種類の錐体(L-cone, M-cone, S-cone)があって、脳内の色の処理はL-, M-coneの情報を使っているparvocellular pathwayとS-coneの情報を使っているkoniocellular pathwayとに分かれて処理されています。このような状況で便利なのがDKL色空間による色表現です。
網膜から上丘へ直接入力するretinotectal pathwayではS-coneの入力がないと考えられています。そこで、輝度は一定のままに、S-coneへの刺激だけが変化するような刺激(S-cone isolating stimuli)を作ってやれば、S-coneを選択的に刺激できます。もしこのような刺激を処理できないとしたら、そのような情報処理はretinotectal pathwayを介しているという証拠になります。
というわけですが、マイナーすぎて日本語による資料がありません。そこでまとめを作成してみました。わたしは神経生理学者でして、心理物理学者ではありませんので、間違いを指摘していただけるとありがたいです。まさにそれこそがこういう文書を公開した理由なわけでして。
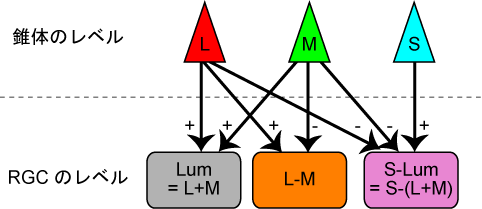
図1 反対色応答
イントロとかは最小限でいきましょう。図1に網膜での反対色応答の形成の図式を作りました。網膜には三種類の錐体(L-cone, M-cone, S-cone)があって、それぞれの足し算引き算によって網膜以降の色処理の三つのチャネル(Luminance, L-M, S-(L+M))ができます。このような三つのチャネルによって、CRTモニタに表示された色がどのように表現されるか、というのがここでの問題です。
図2 色空間とその変換
図2は色空間とその変換の流れです。
図3 RGB 空間
モニタの刺激はRGBそれぞれ256階調ありますので、3次元で表示することができます(図3)。
図4 xyY 空間equiluminant plane
XYZ空間はxyY空間に変換できて、x,yで色を表して、Yでluminanceを表します。有名なCIE xyYのequiluminantな平面です(図4)。
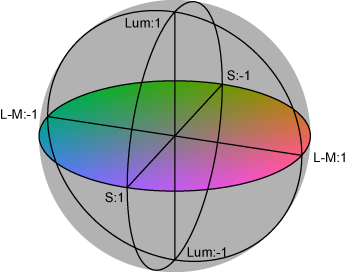
図5 DKL 色空間
LMS色空間はそれぞれのconeでのexcitationの大きさで表現した座標です。LMS色空間は言ってみれば、網膜の錐体のレベルでの色表現です。DKL色空間はさらにもう一段階処理が進んだ、retinal ganglion cell以降での色表現を表したものです(図5)。
じっさいにLGNのニューロンの応答がDKL空間でうまく表現されることが報告されました。この論文がDerrington AM, Krauskopf J, Lennie P (1984) "Chromatic mechanisms in lateral geniculate nucleus of macaque." J Physiol (Lond) 357:241-265.でして、著者の三人の頭文字を取ってDKLと呼ばれるようになりました。それより前に、MacLeod and Robert M. Boynton (1979) "Chromaticity diagram showing cone excitation by stimuli of equal luminance" J. Opt. Soc. Am., Vol. 69, No. 8, p.1183-1186 というのが出版されていまして、こちらはMacLeod-Boynton color spaceと呼ばれます。式の表現の違いはありますが、基本的に同じものです。
[RGB空間からLMS空間への変換]
(省略。くわしくはPDFをどうぞ。)
L-cone, M-cone, S-coneの吸光特性については出版されている論文を使います。いちばん有名なやつは
Smith, V. C. and Pokorny, J. (1975). Spectral sensitivity of the foveal cone photopigments between 400 and 500 nm." Vision Research, 15, 161-171.
ですが、さいきんはもっと新しいので、
Stockman, A., and Sharpe, L. T. (2000). "Spectral sensitivities of the middle- and long-wavelength sensitive cones derived from measurements in observers of known genotype." Vision Research, 40, 1711-1737.
というのがあります。Stockmanのweb siteにデータもあるので、テーブルを自分で入力する必要はありません。これらはhumanのデータです。しかも刺激のサイズによって値が違います。よって、最終的にはなんらか別にpsychophysicalなvalidationの必要性が出てきます。
(省略。くわしくはPDFをどうぞ。)
以上の変換に関しては、Brainard, D. H., Pelli, D.G., and Robson, T. (2002). "Display characterization. In the Encylopedia of Imaging Science and Technology."(pdf file) J. Hornak (ed.), Wiley. 172-188.にくわしい説明があります。この論文はwebからフリーで入手可能です。
さて、このLMS空間からDKL空間への変換(d)を導出するというのがこの文書の本題です。
[LMSからDKLへの変換行列の導出]
つづいて、LMSからDKLへの変換をします。ここの参考文献はBrainard, D. H. (1996). Cone contrast and opponent modulation color spaces."(pdf file) In Kaiser and Boynton, Human Color Vision, 2cd edition, Optical Society of America, Washington, DC.です。この論文はwebからフリーで入手可能です。以下でやってることはこの論文と同じなのですが、計算のステップを省略せずに書いて、確認しながら進めてゆきます。
(省略。くわしくはPDFをどうぞ。)
というわけで、LMSからDKLへの変換は一次変換でした。初めに出てきた図\ref{fig:fig1}での変換(b)-(e)はすべて一次変換です。ですので、PCのモニタで表現できる部分はDKL空間の中では傾いた平行四辺形みたいなものになっています。図\ref{fig:fig3}にあるような球形ではないことに注意。つまり、DKL空間のうちでCRTモニタによって表現できるところは限られていて、normalizationにもよりますが、半径1の球の中で定義できないところもあるし、外で定義できるところもあります。
あと、ここまでゴリゴリやりましたが、じっさいにはbackgroundとしては灰色を設定するでしょう。そうすると、$L_0とM_0とS0$の比を一定にしたままbackgroundの輝度を変えていって、変換行列AとDKL座標の値がどう変わるかを計算してやることができます。そうするとどこかに不動点があるかとかわかるはずですが(Y/2な気がする)、面倒くさいので止めました。
[DKL色空間が持つややこしい点]
前の章で多少言及しましたが、じつはnormalizationの項k_1-k_3は実験の目的によって決まり、一意には定まりません。つまり、pooled cone contrastでのnormalizationは、Lum. L-M. Sの軸での絶対値がperceptualに同等であることを保証しません。もしあるluminanceの刺激とあるS-cone contrastの刺激とを比較したい場合には、detection taskかなんかをやって正答率が同等のところで比較してやらないといけません。このへんの事情については、Brainard 1996でも銘記されています。
また、大元のDKL論文は刺激にCRTモニタを使っていますが、DKL座標のLumimanceの軸の1はMichelson contrastが1になるように設定されています。つまり、DKL = [1,0,0]^tがRGB = [255,255,255]^tだとすると、backgroundがその半分の輝度になるようにしてあります。そしてこのようにして決めたbackgroundからS-lumの軸で提示可能な最大値をS-lum軸の1としています。L-Mについては明示的に書かれてないんですけど。ともあれ、DKLの値はモニタの値に依存します。つまり、モニタのRGBの輝度のバランスによっていくらでも変わるということです。よって、DKL論文だけ引いているような研究だとL-MとSとの絶対値についてはあまり考慮していないと思っておいた方がよいんではないでしょうか。
以上です。謝辞:生理学研究所・感覚認知情報研究部門の鯉田孝和さんにいろいろ教わりました。御礼申し上げます。
2008年10月17日
■ Tononiのinformation integration theory of consciousness
Tononiのinformation integration theory of consciousnessってのに興味があるのだけれど、懐疑的だ。
Giulio Tononi "An information integration theory of consciousness" BMC Neuroscience 2004, 5:42
脳のglobalな特性をどう扱うかという問題意識はわたしは大好き。でも、Edelman-Tononi理論とかBaars-DehaneのGlobal Workspace Theoryとかには近づいてこなかった。こういう人たちの意識論というのはだいたいあまりに機能主義的だと思ってたし、この種の天下り的モデルは信用してない。
そしてなにより、Tononiが使っているeffective information、そしてΦ (integrated information)というものが理解できないので態度を保留してきた。
たとえば、この人の他のempiricalな論文ではこのmeasureが使われていない。たとえばあの睡眠時にTMSをしたScience論文だけれども、ERPの解析をしているだけだ。それどころか、タイトルがeffective connectivityであるにもかかわらず、effective connetivityすら計算されていない。(Effective connectivityじたいが「脳活動から計算された因果性のconnectivity」と「刺激による応答から見たconnectivity」と混ざって使われているようにも思える。)
Tononi自身がどっかで書いていたけれども、empiricalなstudyで定量的に取り出せる値ではないようだ。
また、Φ (integrated information)はeffective connetivityとかなり関連のある概念であるにもかかわらず、その関連が十分説明されていないようだ。
さいきんの論文では言及はされているようだが:Balduzzi D, Tononi G. "Integrated information in discrete dynamical systems: motivation and theoretical framework." PLoS Comput Biol. 2008 Jun 13;4(6):e1000091.
というわけで、このΦが、たとえば大規模同時記録とかからなら計算できるのかとかそういうのを知ったうえでつっこんでみたいと思う。これが宿題。
わたしは神経生理学者としては、意識について脳のglobalな特性からアプローチするネットワークモデルというものがたいがいトップダウン的なモデルだけになってしまって、empiricalなデータと接点を持たないという点にフラストレーションを抱えてきた。
F. Varelaはけっきょくそういうネットワークモデルを提出せずに、gamma-waveの解析だけを提示した。たんに氏の健康状態ゆえだったのかもしれないけど、それには意味があったように思える。
だから、わたしがしたいと思っていることは、ニューロンレベルでの現象ともっとマクロな現象とを繋ぐあたり、つまり、「多ニューロン同時期録でのCCGによるmonosynapticなconnectivity」から、「fMRIやEEGによる脳領野間でのeffective connectivity」までを繋ぐようなものに向かってきていて、それがわたしがこれからBMI/BCIに関わっていこうとするときのモチベーションだったりする。
Information Integration Theoryに関してはわるねこさんの「Information Integration Theory of Consciousness」にくわしい説明があり。
- / ツイートする
- / 投稿日: 2008年10月17日
- / カテゴリー: [トノーニの意識の統合情報理論] [脳科学メモ]
- / Edit(管理者用)
2008年07月13日
■ 神経科学大会シンポジウム無事終了しました!
どもども。ここ最近出張しまくりの激動の日々を送っておりました。5/9-16でフロリダのNaplesでVSSに参加。6/19-23に台北で開催されたASSC2008に参加して、帰国したその日にそのままLAに飛んでUniversity of South CaliforniaのLaurent Ittiのところに2週間弱滞在してHFSP関連のコラボレーションの仕事をしてきました。そのあいだにカルテクにも見学しに行って、Christof Kochのラボでトークをさせてもらってきました。7/6に帰国してすぐに神経科学大会というかんじで、まだ時差ボケが抜けきってません。来週の火曜日(7/15)には京大医学部で開催される電子情報通信学会の7月のNC研研究会でトークです。ここまででひとくぎり。しばらく岡崎でおとなしくしているつもりです。お次のイベントはいよいよ生理研研究会(9/11-12)でしょうか。
神経科学大会のシンポジウムのほうですが、会場は立ち見の方も出て、熱気ある雰囲気の中で進めることができて非常に良かったのではないかと思います。質問もたくさん出たし。あれだったら総合討論の時間を入れても良かったかもしれませんね。私自身に関しては、座長としてのしゃべりとか、トーク自体とかけっこうひっかりまくってましたが、もう勢いでなんとかしたというかんじです。
また別にレポート書くかもしれませんけど、まずは来てくださったみなさま、どうもありがとうございました。以下にシンポジウムに言及してくださったブログをリンク:
* スウィングしなけりゃ脳がない!
* ◎脳研究のタマゴ◎
* 大「脳」洋航海記
shokou5さんとは前日の懇親会でもお話を。私からは、フジロックのマイブラ行きたいけど行けない!とかこれからは萌えとロボティクスだとか(またかYO!)。
vikingさんも見に来てくれてシンポジウム後にお話ししました。
(前回のエントリにコメントしてくださったatsushiさん、Hikaru.Tさんのコメントは内容を考慮してこちらに移動させていただきました。)
# atsushi
初めまして!いつもblog拝見しております。
神経科学大会の口演お疲れ様でした。
もうほとんどあのセッションがメインで行ったので、とっても楽しかったですし、visual awarenessのセッションは今大会で一番盛り上がってた気がします。
僕は学部生でまだ脳について勉強し始めたばかりなので全然知識もないですが、視覚意識にすごく興味があるのでこちらのblogでもかなり勉強させて頂いてます。
お話できる機会があればよろしくお願いします♪
あ、もし両側V1破壊したらvisual awarenessは片側のときと同じようにdSCを使ってretainされるんでしょうか?
pooneilさん Neuroscience2008、お疲れ様でございました!atsushiさんと同じく、僕も今回のシンポジウムを一番楽しみにしておりました。想像以上に興味深く拝聴させていただきました。本当にありがとうございました!!
# pooneilatsushiさん、Hikaru.Tさん、どうもありがとうございます。この分野に興味を持っておられる方がたくさんいることをシンポジウム会場でも実感しました。来年の神経科学大会は開催地が名古屋で、当ラボの伊佐教授が大会長です。Plenary lectureのひとつはChristof Kochが確定しています。おそらくは来年も意識関連の話題を扱うことができるのではないかと思います。ご期待ください。
atsushiさんの質問ですが、両側の条件を扱った研究は大昔に行われていますが、この条件でのdSCの寄与を直接検討した研究はないと思います。おそらくは片側のときと似たものが見つかるのではないかと予想していますが。ちなみに両側の条件を扱った研究で有名なのは1970年代にニコラス・ハンフリーによっておこなわれたものです。単行本で「赤を見る―感覚の進化と意識の存在理由」ニコラス ハンフリーというのが出版されていて、これの中にけっこう記述があります。ご興味ありましたら読んでみるとよいかもしれません。
吉田さん、
土谷です。
視覚意識のシンポジウムは結構評判よかったみたいっすね。リンク先のページも見てきたら、俺よりもさらに若い世代の人らが興奮してたみたいで、よかった。これなら、来年も何かネタを考えて、やっても良いかも、と思い始めました。クリストフが、トークだけじゃなくて、シンポジウムみたいなものも、やりたい、みたいに言ってたし。
ただ、何人かの感想だけだと、フィードバックとしては物足りないですね。ASSCみたいに学会後アンケートがないので、これらの意見が客観的に評価できないのが問題です。そういうアンケートの集計とかって、おそらく外部に頼む必要があるとか、色々大変だと思いますが、将来学会を改善していくには非常に有効だと思いますので、来年の時には是非実施してください!
総合討論もいいかとは思うんですが、あんまり質問とか出て来ないとサムイので、TICSみたいに、"outstanding questions" を各演者が用意しといてそれをたたき台に最後に20分ぐらい時間を取るとかすれば良かったなと思います。
あと、来年の会場は会場内で移動しやすいところが良いです。マジで自分のシンポジウムに遅刻しそうだったので...
# pooneil土谷さん、シンポジウムではどうもありがとうございました。
> クリストフが、トークだけじゃなくて、シンポジウムみたいなものも、やりたい、みたいに言ってたし
これいいですね。ぜひ実現できるように各方面に働きかけたいと思います。
フィードバックに関してですが、学会後のアンケートは今回もありました。(要旨集の冊子に挟まれてる。) ただ、特定の講演に対する感想というのはありませんでした。ですので、たとえば現状のアンケートに「いちばん面白かった講演(シンポジウム・ポスター)はなにですか?」というようなものを作って集計・発表するとかいうやり方はあるかと思います。
総合討論に関してですが、じつは今度9月に行う生理研研究会(「動機づけと社会性の脳内メカニズム」)では総合討論の時間を取ることにしているのですが、まさに土谷さんが言うのとまったく同じことを計画しています。「TICSみたいに、"outstanding questions" を各演者」に用意しておいてもらうといい、と松元さんにもメールしていたのですが、TICS、というところまでまったく同じなのですごいびっくりしました。(それとも前にこの話聞きましたっけ?) さらに、あらかじめ若手の人から総合討論でのお題を募集しておいて、面白いお題を出してくれた人にトラベルアワードを出す、ということも計画してます。このくらいやればそれなりに盛り上がるんではないか、というわけです。研究会の方はそんなかんじでそれなりに準備が可能ですけど、学会でやるのは難しいかも。
来年の生理研研究会も秋にやることになるかと思いますが、もしかしたら神経科学大会(9/16-18)に近接して設定して、シンポジウムの招待講演者にそちらにも出てもらうとかそういうやり方ができるかもしれません。この場合は「国際研究集会」という枠組みになると思いますが。もうすこし考えてみます。
来年の神経科学大会の会場はもう決定しています。今年ほど複雑ではありませんが、建物の右と左を行ったり来たりしないといけないかもしれません。ちなみにフロアマップ。
# vikingどうも、シンポジウムは大変興味深く拝聴いたしました。nhpには詳しくないもので、質問が冗長になってしまったかもしれません。今後はもう少しかっちり論点をまとめてから伺うようにいたします。
自分のblogでも書きましたが、東京国際フォーラムは思った以上に使いにくかったですね・・・名古屋は5年前に行ったことがあって使いやすかった記憶(でも周囲に食事できるところが全然なかった気も)があるので、期待したいと思います。
# pooneilvikingさん、どうもありがとうございます。ブログをリストに追加しておきました。
そうです、名古屋は昼ご飯が問題なのです。金山まで出るとそのまま戻ってこなかったりとかして。宮下研のときの横浜での神経科学大会みたいにフードコートとか作るとかそういう手が必要かもしれません。
匿名問題ですが、私自身は名前を出していることからもおわかりのように実名派です。わたしの場合、実名で情報発信することのメリットはむちゃくちゃ大きかったと思います。自分が名前を出してさらけ出したからこそ、安心してコメントを書き込んでくれて、自分が何者であるかも明かしていただけた方も多かったのではないかと思います。ここを元にして出来た人の繋がりに感謝しております。
とはいえ、実名であるが故に、ポスドク問題など、荒れる問題には意識してタッチしないようにしたりとか、それなりに気を遣っているのも事実です。(言いたいことはあるんだけれど、言葉尻を取られないようにきっちり書かないといけないかと思うと気が重い。) これはわたしのnhpという言葉遣いを理解しているvikinさんならご存じではあるかと思いますが。
とりとめが無くなってきたのでこのあたりで。
>pooneilさん
そうですね。どちらを選択するかは悩ましいところですが、まぁそれぞれの考え方があっても良いかなと僕は考えております。もっとも、僕のblogはあまりにも業界内では有名になりすぎていて、素性を隠す理由がもはやなくなりつつありますが(汗)。
pooneilさん、大変お忙しいスケジュールの中、質問に丁寧に答えて頂きありがとうございました。m(__)m
リンクも貼っていただき、ありがとうございました。
今たまたま京大にいるので明日の講演も拝聴させて頂くと思います(ストーカーみたいですが笑)。楽しみにしています。
では。
いつも楽しく拝見させて頂き、大変勉強させて頂いています。意識に興味があってサルの電気生理を始めた僕としては、先日のシンポジウムは大変刺激的で興奮してしまい、一枚脱いでしまう程でした(実際会場自体に熱気がこもっていたのもあるかもしれませんが)。
僕が知らなかった事が多すぎたせいかもしれませんが、日本の意識研究があのセッションで確固たるものになりつつあることを示された、そんな歴史的な瞬間であるように思われました。
神経科学者SNSのイベントで学会の時に飲み会があるようなので、是非これから先、参加させて頂こうと思いますので機会がありましたらよろしく御願いします。
# pooneil桑原さん、どうもありがとうございます。
歴史的な瞬間、とはさすがにびっくりですが、これからもいろいろ仕掛けていくつもりです。
よろしくお願いします。
いま過去のエントリを見てたら昨年自分で書いてた>>「TICSみたいに、"outstanding questions" を各演者」(20071019 http://pooneil.sakura.ne.jp/archives/permalink/001119.php)
2008年05月24日
■ ASSC12近づいてきました
ASSC12が近づいてきましたね! 今年は台北で開催されます。とりあえずホテル予約したりとか活動開始。中山駅前にホテルを予約して士林夜市へ行く気満々。
プログラムと要旨も出そろいました。ざっと見たいセッションをまとめておきます。
22日午後のわたしの出るセッションはかなり良いですよ。はじめがわたしのblindsightの話、つぎがMotion Induced Blindnessの話ってこれは20080515で書いた、attentionかけた方がawarenessが無くなるっていうKen Nakayamaラボの人の話。そのつぎが金井良太さんのperceptual fading + rTMS。おそらくは"Time-locked Perceptual Fading Induced by Visual Transients" J Cog Neurosci 2003でやったことをVincent WalshのところでrTMSと組み合わせたという話のはず。んで最後がAlex Maierのflash supression中のV1のLFPを記録してCSDでlayerの議論をするという話。これは今度の7月の神経科学大会のシンポジウムに出してもらった内容と同じ。このセッションがいちばんニューロサイエンス色が強いんではないでしょうか。
土谷さんは21日にAlexとのコラボレーションの方の話をします。MTでのdecodingの話。Awarenessとattentionの関係の話の方は22日にKochが話すみたい。んで、このKochの入っているシンポジウムですが、これはさいきんのNed Blockの"Consciousness, accessibility, and the mesh between psychology and neuroscience"(PDF) Behavioral Brain Sciences 2007でのバトルが再燃するものと思われます。というのも初めに出てくる3人(Christof Koch, Victor Lamme, Sid Kouider)はみなNed BlockのBBSにコメンタリを書いた人でして、それを最後にNed Blockが迎え撃つ、という形になっています。予習超重要。ちなみにNed Blockはphenomenal consciousnessとaccess consciousnessという分け方を提唱した人。そんなもん一緒に決まってるだろとか前者だけがconsciousnessだろとか煽ってみたいところだけどこういうのはきっちり本文に当たらないとニュアンスが消えてしまう。BBSの要旨を訳したものがshokou5さんのところにあります。ちなみにshokou5さんのところの「悦ばしき知識:鏡について」にはArational Agentさんが登場してます。20060111のコメントにもあるようにArational Agentさんの(元)ボスがThomas Metzinger。ASSC12でもKeynote lectureをやります(ここまで話を繋げてみたかったというわけ)。
じつはわたしがいちばん期待しているのは川人先生のKeynote Lecture。川人先生は1997年にKawato M (1997) "Bidirectional Theory Approach to Consciousness"(1994年に京都であったシンポジウムをOxford Pressで単行本化したもの)で、自意識のことを、sensorimotor interactionのmodule(これ自体が内部モデル)をモニターして脳自身のモデルを作る内部モデルと捉えていました。そこから10年経ってどういう差分があるか、ということに興味があります。(川人先生は強烈なMarrのproponentですから、明確に表象主義の立場を取っていて、上記の考え方がRosenthalの表象主義的意識論と親和性を持つものであることもこの論文で明記されています。一方で内部モデル説はsensorimotor theory、つまりawarenessは行動とカップルしている、というような立場とも親和性があるのではないかと私は考えています。これがわたしの「順モデル=awareness論」です。もっとも、川人先生もこの論文で「順逆モデルでの緩和過程によって、von der Malsburgの言うcoherent perception and behaviorが達成される」という言い方をしています。たぶんほとんど同じことを言ってるんではないかと思うのですが。さらに言えば、川人理論に基づくといわゆる表象説とsensorimotor説は違うドメインの議論をしているのだ、なんてまとめ方が可能だと思ってます。なんつてまた超大事なことをカッコの中で書いてみました! 北_田暁大メソッドと命名。*注)
*注 アマゾンの嗤う日_本の「ナシ_ョナリズム」の書評でのモワノンプリュ氏による「ついでながら、この著者は『責任と正義』でも重要なことを注や補遺に紛れ込ませていたが、今回も自分の立場表明を目立たない注(p264)で行っており、悪癖だと思う。」が元ネタ。
# shokou5
ご無沙汰しております。記事をふたつもご紹介いただきありがとうございました。ASSCも台北も羨ましいです。わくわくするようなセッションが目白押しですね。なんとゆうか、こういうことをまじめに議論している人たちが世界にこんなにたくさんいるってゆうことがとても素敵だと思います。僕もいつかは関連する研究をやって出席してみたいものです。
数年前台湾旅行したのですが、士林の近場だと北投温泉がお奨めです。日帰り入浴の温泉あります。時代と場所がわからなくなるようなインチキな温泉街な感じです。
どうもこんにちは。そうでした、トラックバックしておけば良かったんですが、さいきん習慣が無くなってまして。
「悦ばしき知識:鏡について」はやりとりの展開の過程が面白かったです。
北投温泉いいですね。一日まるまる使えたら淡水とかと併せて行ってこれるのですが。
2008年05月17日
■ VSSポスター見に来てくれた人
私の発表の方ですが、今回はポスターでした。たしか口演で申し込んだと思うんですが、落ちた。ま、繰り返してプレゼンスを上げてゆくしかないです。
んで、けっこうたくさんの人が来てくださったので良かったです。今回は若い人だけでなくてPIクラスの人も見に来てくれた。これはよかった。あとさすがVSSで、Psychophysicianからいろいろ助言をいただいて感謝してます。
Mervin Goodaleが見に来てくれて、すごい面白がってくれた。論文出たら送ってくれって言われた。
眼球運動関連だと、Richard Krauzlis(Salk Institute)、Denis Pelisson(INSERM)にもひととおり話を聞いてもらった。このへんを説得させることができるかどうかが出版時は大きな問題になるはずだけど、納得してもらえた様子でした。
それから、Christian Casanova(Universite de Montreal)、Harry Orbach(Glasgow Caledonian University)、Rufin Vogels(Katholieke Universiteit Leuven)など。
下條信輔先生(Caltech)にはpsychophysicianとしていろんな突っ込みを。この論文でどこまで突っ込んだ方がいいか思案していたところはあったのだけれど、hit-missの比較だけよりはもうちょい行ったほうがいいかなという気がしてきました。ちなみに質問いただきましたが、nhpでconfidence ratingというのは記憶課題に関してはあることは知ってます:"Rhesus monkeys know when they remember" PNAS | April 24, 2001 | vol. 98 | no. 9 | 5359-5362。せっかくなんで調べてみたらそれ以外にもいろいろある様子です:Confidence judgments by humans and rhesus monkeys. J Gen Psychol. 2005 Apr;132(2):165-86、それから、Confidence judgments by rhesus macaques on a serial memory task(PDF)。このへんを組み合わせるともっと面白いことができるかもしれません。
それから佐々木由香先生(Harvard Medical School)にもくわしく説明を。SDTのあたりは"this is neat"と言っていただけたました。
それから日本から来た先生方ではNTTコミュニケーション科学基礎研究所の五味裕章先生、東京大学総合文化研究科の村上郁也先生。東北大学電気通信研究所の塩入諭先生にはawarenessとはいったい何ぞや、みたいな話になりました。
あとは若い方にもたくさん。カルテクの土谷さんともHakwan Lauのbiasの話をしました。そのへんはこんどのASSCのポスターに入れる予定です。あと、若い方でなんどか話をした経験があるのだけれど未だ名前を覚えてない、という人も複数いたりするのでそのへんと知り合いになっときたいのですが、私引っ込み思案なもんで(ホント)。
というわけで皆様ありがとうございました。このネタでなんとかデカイの打ちたいんですけどね。
# abc
pooneilさん初めまして。よく拝見させていただいています。
上に挙げられている、Lisa Sonのサルの実験のビデオが彼女のページにアップされています。
トークンを失ったときのサルのがっくり感がとても面白いです。
http://www.columbia.edu/~lks16/LisaVideos.htm
# pooneilどうもこんにちは。いまやっとムービーが見れました。本気で頭抱えてますね。これはおもしろい。
ご紹介どうもありがとうございました。
これどのくらいトレーニングしたら可能なんだろ。
2008年05月15日
■ VSS終了
VSS08終了しました。出発まではあと半日あるけどいろいろやるべきことが。
自分の発表についてはまたこんど。ともあれ見たセッションのまとめを:
------------------
5/9
------------------
16.152 Improved blindsight near the hand is associated with increased fMRI activation in the superior parietal-occipital cortex
Liana Brown, Jody Culham, Greg Kroliczak, Melvyn Goodale
Patient MB。Temporal lesionでoptic radiationを損傷して四分の一視野を損傷したblindsight。
Targetの近くに手を添えるとreachingの成績が良くなる。fMRIでAIPとV6aが活動する。
------------------
5/10
------------------
9:30 am
21.15 Collicular vision guides non-conscious behavior
Marco Tamietto, Franco Cauda, Luca Latini Corazzini, Silvia Savazzi, Carlo Marzi, Rainer Goebel, Lawrence Weiskrantz, Beatrice de Gelder;
Weiskrantzおよびde Gelderが入っているから例のaffective blindsightの患者さん。
fMRIでSCの活動をみてる。でも、fMRIのデータはスライド一枚で一瞬で終了。
Purple color patchの刺激だとS-coneを主に刺激するから上丘の活動はなくなる。
(厳密な意味でのS-cone isolating stimulusを使っている訳ではないらしい。)
いっぽうで、pulvinarの活動はPurple color patch刺激でもなくならない。
Pulvinarといってもどの領域かで大違いなのだけど、そのへんの情報はなし。
Pulvinarの活動は同側の上丘からだけではなくて対側からとかextrastriateやもっといろんなところからトップダウンで来てる可能性があるんで、これだけでは何ともいえないけど、同側の上丘からだけで決まる訳ではないということははっきりした。
------------------
5:30 pm
25.25 Motion induced blindness: The more you attend the less you see
Olivia Carter, Robert Luedeman, Stephen Mitroff, Ken Nakayama
MIBで二つ点を出しておいて、片方をattendするように指示を出しておくとそちらがより消えやすくなる。つまり、attentionとawarenessが相関しない例がまた一つ増えたということ。
------------------
5:45 pm
25.26 Adaptation-induced blindness
Isamu Motoyoshi, Sayuri Hayakawa
今回みた発表で一番印象的だった。Gaborのmotion gratingでadaptationさせたあとで同じ向きのgabor(テスト刺激)を見せるとadaptationによって検出閾値が上がるのだけれど、テスト刺激の出し方をじんわり出してやってabrupt onsetがないようにしてやるとテスト刺激が見えなくなる。かなり強烈。おもしろい。Nature二つ目いったかも。
すぐイメージングやった方がいいと思った。私もこれの応答を記録してみたい。
------------------
26.413 Effects of luminance contrast on visual responses in frontal eye field
Richard Heitz, Geoffrey Woodman, Pierre Pouget, Jeremiah Cohen, Jeffrey Schall
FEFでcontrast変えて刺激の応答をみると、暗くなるごとにonsetが遅れてゆく。立ち上がりがぬるくなるとかじゃなくて。
------------------
5/11
------------------
8:30 am
31.11 Preserved motion processing and visuomotor control in a patient with large bilateral lesions of occipitotemporal cortex
Melvyn A. Goodale, Marla E. Wolf, Robert L. Whitwell, Liana E. Brown, Jonathan S. Cant, Craig S. Chapman, Jessica K. Witt, Stephen R. Arnott, Sarah A. Khan, Philippe A. Chouinard, Jody C. Culham, Gordon N. Dutton
8:45 am
31.12 Preserved processing of motion and dorsal stream functions in a patient with large bilateral lesions of occipito-temporal cortex
Jody C. Culham, Jessica K. Witt, Kenneth F. Valyear, Gordon N. Dutton, Melvyn A. Goodale
これまでの患者D.F.さん(LOCのbilateral lesion)での知見をより大きな損傷を持つ新しい患者さんM.C.さんで再現。両側でMTより後ろがまるまるない。それでもmotionは感じることができる。つまり、Ridoch syndrome。両側でpulvinarのactivationあり。
------------------
5/12
------------------
43.423 Neural Dissociation between Visual Awareness and Spatial Attention
Valentin Wyart, Catherine Tallon-Baudry
JNSに出てたネタに関して、刺激が出る前のgammaからawarenessがあるtrialを予測できる、というもの。
問題意識は近いけど、みてるところがvisual cortexなのでselection biasとかそういう話に入ってない。
Attentionではないことの証拠としてはcued locationに依らないということで押さえている。
こっちはいい。
Arousalではない、ということの証拠には、arousalで上がることが知られているalphaではこのような予測ができないことを示している。これは傍証ですね。Selection biasみたいな発想がない。
------------------
43.508 Neural Correlates of Motion-Induced Blindness in the Human Brain
Marieke Scholvinck, Geraint Rees
MIBで点が見えたときと見えなかったときとをボタン押しで知らせてfMRIで比較するとV1,V2で局在した活動がみられるのだけれど、見えたときの方が活動が低い。なんだこれ?
------------------
43.527 Application of reverse correlation to the study of visual and extraretinal signals in the macaque superior colliculus
Christopher Pack, Jan Churan, Daniel Guitton
上丘でreverse correlation。1-5 degくらいの明点、暗点が全面積の1-5%くらいに出る。12ms/frame。SGIのvisuomotor neuronから記録している。
だいたいexcitatory fieldだけ。Inhibitory fieldのあるものもあるがすくない。時間応答は明点、暗点どちらでもexcitatory-inhibitory。Latencyは40-60ms。
Presaccadic remappingも見ているが、reverse correlationだとmodulateされるニューロンが見つかってこない。一緒に聞いてた村上さんも同意してたけど、同じ方法でFEFやLIPの結果を見てみたい。
------------------
5/13
------------------
9:30 am
51.25 Decoding orientation-selective responses to real and illusory contours
Frank Tong, Yukiyasu Kamitani
以前のNature Neuroscienceの方法でillusory contourを見せてdecodeできるかどうか検証。
Real contourだと成績が90%なのに対してillusory contourだと70%程度。V1,V2,V3のどこを使っても成績に差はない。
0-90degのかわりに45-135degのillusory contourを使うとなぜかV1ではreal contourと同じくらいの成績が出る。
------------------
11:45 am
52.26 A Bayesian Model of Visual Search and Recognition
Lior Elazary, Laurent Itti
Surprise modelを使ってsearch and recognition (=ウォーリーを探せ)をさせるとSIFTやHMAXと同じくらいの成績が出る。しかもかかる時間はsurprise modelのほうが1000倍早い。
------------------
53.514 A visual target in the blind hemifield of hemidecorticate patients reduces latency and improves accuracy of antisaccades
Olga Savina, Andre Bergeron, Daniel Guitton
Guittonのところで論文が出てるhemidecorticated patientの話。Normal fieldに刺激を出してblind fieldにAntisaccade。加えてBlind fieldに刺激を出すとreaction timeがはやくなる。
こんどはeccentricity変えて、弁別できるかどうか調べるって言ってた。
Olgaさんは私のポスターにも来てくれてすごく熱烈におもしろがってくれた。昨年のSFN発表もreferしてくれてるし。
患者さんについて論文に出てこないあたりをいろいろ教えてもらった。
------------------
3:15 pm
54.14 Identification of optic radiation in-vivo using diffusion tensor imaging and fiber tractography
Anthony Sherbondy, Robert Dougherty, Brian Wandell
DTIでoptic radiationを可視化。ConTrackというやつでLGN-V1をつなぐところ、というかんじで指定してやるときれいに可視化できて、昔のdisection法に依るものとconsistentな結果を得ることができた。これは応用すべき。
図へのリンクがJ Visionのsiteにあり。
この話はもともとYamamoto et.al., 2005でDTIによるoptic radiationのimagingの結果が、Ebeling 1988のdisection法による結果とずれている(たとえば、optic radiationのMeyer's loopの部分のanterior tipのレベル)という問題点があったのだけれど、それをDTIの方法の側面の改良で解消した、という側面があるらしい。
------------------
3:30 pm
54.15 Topography of responses to colour and luminance in human subcortical visual pathways as revealed by high-resolution fMRI at 7T
Marcus Grueschow, Jochem Rieger, Jörg Stadler, Claus Tempelmann, Hans-Jochen Heinze, Oliver Speck, John-Dylan Haynes
7TマシンでLGNとSCをimaging。上丘でもcolor刺激(L+M, L-M)のコントラストに比例して活動が大きくなる。
質問で、上丘はpulseの影響が出やすいところだが補正しているか、と聞かれて、補正してないと言ってた。
------------------
2:45 pm
54.22 Decoding Frequency and Timing of Emotion Perception from Direct Intracranial Recordings in the Human Brain
Naotsugu Tsuchiya, Hiroto Kawasaki, Matthew Howard, Ralph Adolphs
以前生理研でトークをしてもらったときの前半部と同じ内容。マイケルジャクソンの顔のモーフィングは聴衆に受けてた。
------------------
6:00 pm
55.27 Boosting perceptual learning by feedback manipulation
Kazuhisa Shibata, Shin Ishii, Noriko Yamagishi, Mitsuo Kawato
以前ASCONEのときにチューターをしてくださった柴田さん。
Perceptual learningの成績のフィードバックを実際よりよいものを返してやるとlearningが促進する。Perceptual learningですら「褒めると伸びる」ということなのか、ともあれそのへんの解明が進んだら面白い。
------------------
56.317 Strong exogenous attraction to attention by unique eye of origin --- evidence for a bottom-up saliency map in the primary visual cortex
Li Zhaoping
以前からTICSとかでも出てる、V1にsaliency mapがある、という話なのだけれど、実験的検証のためにKolb and Baun 1995のblindsight in normal subjectのパラダイムを援用してる。Kolb and Baunに関しては私も興味あって、昨年のASCONEで使ってみようかと思って調べてみたらこの実験自体は再現性がない、とか問題になったことがあったのを知ってやめた経緯あり。続報も出てないし。
とか思ってたら今回関連するネタが出てきたのでもう少し読んでみた方がいいかも。J Visionに出版済み(Journal of Vision (2008) 8(5):1, 1–18)。
ちなみにこの件についてLaurent Ittiとも話をしたけど、Li Zhaopingのいうsaliency mapはIttiがいうところのfeature mapであって、featureごとに持っていて統合されていない。このへんでreconcileできるらしい。
------------------
56.519 Dissociating preattentive vision and preattentive attentional guidance
Louis Chan, William Hayward
Detection taskだとattentional captureがなくて、localization taskだとattentional captureがある。
Featureのswitchではdetection taskでだけ遅くなる。
だからdetectionはfeature mapに直接アクセスしていて、localizationはsaliency mapにアクセスしている、という結論。
解釈の妥当性はおいておいて、この図式は面白いかも。
------------------
56.527 Modeling interactions between visually-responsive and movement-related neurons in FEF during saccade visual search
Braden A. Purcell, Richard P. Heitz, Jeremiah Y. Cohen, Gordon D. Logan, Jeffrey D. Schall1, Thomas J. Palmeri
Diffusion modelで積分値が重要か元の値が重要かという検証だったけどどちらも同じくらい当てはまりがよい、という残念なオチ。
------------------
2008年03月28日
■ ASSC2008口演やります
今年の意識学会(ASSC)の年会は台北で行われます:ASSC2008。
昨年はポスターを出しました。そのときの様子は20070625あたりに。
ここで土谷さんとAlex Maierと知り合ったので私にとっては非常に重要な学会です。ちなみに今度の神経科学大会(東京)であと私とATRの神谷さんの四人でシンポジウムをやります。タイトルは「主観的視知覚の神経メカニズム-心理物理、fMRI、電気生理による実験的アプローチ」です。ぜひ見に来てください(宣伝)。
でもって今年のASSCのほうですが、はじめてアジアで開催されるASSCの年会ということもあって、各方面から働きかけがあって、日本からも多くの方が参加して講演をされます。スケジュールはこちら。Keynote Lecture 4コマのうち二つが日本からで、ATRの川人光男先生と霊長研の松沢哲朗先生が話をされます。それからシンポジウムでは理研の田中啓治先生が話をされます。それからTutorialでは順天堂の北澤茂先生とNTTの西田眞也先生によるものがあります。
というわけでわたしもここはチャンスと見てオーラルのセッション('concurrent sessions')に応募していたのですが、これが無事通りました。まだ時間は未定ですが、現在出ているプログラムを見ると、三日目(6/22)に"Blindsight in Normal and Lesioned Brains"というセッションがあるのでここに間違いないでしょう。
ということで論文出せてないのにorzいろんなことやってます。こういう活動が実のあるものとなるよう祈りながら。
それでは、台北で僕と握手!
# 金井
こんばんわ。
いま、ASSCのプログラム見ていたら、おなじセッションみたいですね。
http://www.ym.edu.tw/assc12/concurrent_sessions.html
おひさしぶりです。
ASSCのプログラム、アップデートしてたんですね。(メールでの連絡は来ないのだけれど、webの方は着々とアップデートしているようです。)
このあいだまではセッション名だけがわかっていたので、"blindsight"のセッションはどんなかんじになるのかと思ってましたが、要はMIBやflash suppressionなどを含めたperceptual suppression全般を含んだ表現として使っているようです。
なるほど、いっしょのセッションです。Vincent Walshのところに移られてからの仕事がまとまったのですね。仕事が順調に進んでいるようで素晴らしいです。http://kanair.cocolog-nifty.com/blog/2007/12/ucl_a101.html
たしかに、blindsightというにはセッションの内容が、blindness全般ですね。最初はアジアでやるASSCにどれだけ人がくるのか心配していたのですが、おもしろそうなトークがたくさんあるので楽しみです。台湾で会いましょう。
2008年01月01日
■ Intrinsicな注意、extrinsicな注意
あけおめ。(フライング気味。)
「注意」について、また無防備に書いてみます。
注意の分類で、ある場所に注意を向ける"spatial attention"と、ある物体の属性に注意を向ける"object-based attention"みたいな分け方がありますが、あのobject-basedって言葉が気に入らないんです。ここでいう属性とはあるobjectにintrinsicな性質のもので、あるobjectの空間配置のようなextrinsicな要因に向けるのがspatial attentionであると考えると、intrinsicかextrinsicかと分類することが出来るのではないでしょうか。もしくは、object-basedとnon-object-basedに分けてみて、objectに向けた空間的注意と虚空に向けた空間的注意とを分けてみるとか、あるtime pointに向けた注意とかをnon-object-basedに入れるとか。なんてことはすでに言われているのだろうなあと考えつつ、ここからさらにひねり(脱線)を加えてみます。
言った舌も乾かぬうちからというかんじですが、では空間的配置というのは純粋にextrinsicなものでしょうか。物体は空中にとどまることはできないし、軽いものは水に浮きます。空間的配置はそのobjectのintrinsicな特性によって制限されるという意味においてpurely extrinsicではないわけですね。
そうして思い至ったのが、affordanceというものはobjectを外から操作してやるというextrinsicな要素へ変化を加えようとするものなのだけれど、そのobjectのintrinsicな要素によって制限され、形作られる。思えばaffordanceという概念はそのようにobjectのintrinsicな要素をその外まで引き延ばして考えるような思考法ではないか、というわけです。もう少し先に行けるのではないかと思うのだけれども、唐突にここで話は終わってしまうのでした。がーん。
# viking
さすがはpooneilさん、affordanceに絡めるとは面白い視点ですね。そうしますと、attentionは常にそのobjectの持つcontextに影響を受けるということになるような・・・例えば、騙し絵か何かを見せられていて、AとBという解釈が可能な時に仮に被験者がAとしてか認識できないとしたら、Bにおいて○○(Aで物、Bで顔に見えるとしたらBにおける「目」とか)に対してattentionを向けろといわれても、被験者はたぶんできませんよね。ただ、それがcontextの処理だけを意味するのか、attentionのaffordance由来の挙動に関連するのかは、何とも区別しがたい気がします。
2007年11月30日
■ TEMPOのつぎはなんですかね
(過去日記10/10)
いよいよ明日が研究会です。んで、科研費申請書書きとかしたり。泣きそう。
それとは関係ないんだけど、いま実験にはTEMPOを使ってます。フリーズすることもないし、プログラムを作るのとかもいったんマスターしてしまえばとくに問題はないのだけれど、高い。
あと、ディスプレー関係は弱い。いまどき1024*768あたりまでしか対応してないし、ボードとかの依存性があったりして、もうひとつ新しいシステムを作ったときはけっこう苦労させられた。心臓部はいまでもDOSなんでいろいろ限界に来てるんですよね。
あと、刺激提示に関してはサンプルプログラムが少々あるぐらいで、contrastいじったり、ガンマ比調整したりとかpsychophysicsをやるのには向いてないかんじです。
というわけでつぎシステムを構築するとしたらなにかなあと思うんですが、ひとつはpresentationですかね。fMRIやMEGのではこれの人が多いですよね。電気生理でどのくらい使えるか知りたい。
45日間試用可能で、一年のライセンスが240ドル(code activationにて)。
ググってみたらいくつか資料を見つけました。
せりか式
presentation入門(ATR脳活動イメージングセンタ)(PDF)
オンラインでタスクを分岐させることも出来るようだし、データをどこに貯めるかだけ考えれば問題ないような気がします。電気生理の方のシステムをどうするかによるのですが。
ヒト実験との連携を考えて、MATLAB + psychophysics toolboxということを考えていて、こちらもちょこちょこいじっては挫折してたのだけど、ASCONEの準備で光明を得ました。このへんについてはまた。
どうあれ、けっきょくは電気生理側の測定系との絡み次第なのですが、いまどきはなにがいいんですかね。
-----
というエントリを作ったまま放置してたので貼っておきます。Presentationでは電気生理データは追従できないですかね? あと眼球運動によって課題が分岐できる(たんに正しく見ていれば刺激が消えて報酬が出るとかだけど)かどうかがクリティカルなのですが。
# viking
TEMPOのことはあまり詳しくないんですが、TEMPO単体をデータ収録側に使って、(別のPC上の)Presentationで刺激を制御するというようなことでしょうか? トリガ信号をBNC分岐コネクタなどでTEMPOとPresentationの両方に入れてやれば、それなりにきちんと時間的同期の取れたデータの取得が可能だと思います。Presentation自体はWindows環境に合わせたルーチンの最適化をしていて、時間的同期の点ではそれほど問題ないはずですので。
コントラストとかガンマ比調整とかをなさりたいのであれば、VSGなどもよろしいかと。個人的には視覚実験はこちらの方がお薦めです。
http://www.crsltd.com/catalog/visage/index.html
現用版はViSaGeって言うんですね。実は今調べて初めて知りました。
大した情報でなくて恐縮ですが、以上ご参考までに。
# pooneilどうもありがとうございます。
TEMPOの代替のことを考えてます。値段がpresentationの10倍以上しますので。
具体的にはPlexonで電気生理データの収録が出来るようだったら、タスクコントロールと刺激提示の部分だけ出来ればいいのでそういうものでいちばんいいのはなんだろうか、というわけです。
ViSaGeの紹介どうもありがとうございます。もすこし調べてみるつもりです。ではまた。
なるほど、そういうことでしたら色々迷ってしまいそうですね。
E-Primeなんて物もありますけど、あれはトリガ入力に対する時間的同期の取り方が難しいので特に電気生理には向かない気がします。やはりPresentationがベストかと。
ViSaGeがVSGと同規格なら、コントロールはCコンパイラ(WinVC++でもOKです)が必要です。ライブラリだけ導入して、制御は別に何かプログラミング言語でやる必要があります。
2007年11月23日
■ Scholarpedia
調べものでwebを探していたら、Scholarpediaがいつのまにかたいへん充実しているのを発見しました。
書き上がったもののリスト(All articles)のところからいくつか探してみると、偉い先生がcuratorをやってたりします。以下に気づいたところをリスト。リストでは著者をはしょったりしてます。あと、まだレビュー中のものもあります。
- Amygdala by Joseph E. LeDoux
- Reward by Wolfram Schultz。Figure 1が例によって「当店の人気盛り込み」だったんで笑った。
- Reward Signals by Wolfram Schultz
- Mirror Neurons by Giacomo Rizzolatti
- Habenula by Okihide Hikosaka
- Cortical Memory by Joaquin M. Fuster
- Blindsight by Larry Weiskrantz
- Consciousness by Bernard J Baars
- Functional Imaging by Karl J. Friston
- Functional Magnetic Resonance Imaging by Seiji Ogawa
- Chaos in neurons by Kazuyuki Aihara。レビュー中。レビューの文章を見てると面白いです。
- Synesthesia by Vilayanur S. Ramachandran
- Binding by Synchrony by Wolf Singer
- Introspection by Morten Overgaard
私が知ってる若い人も書いてます。
- Visual Salience by Laurent Itti。Saliency mapの方はべつにエントリができているのでした:Saliency Map by Ernst Niebur
- Awareness of Intention by Hawkan Lau
- Attention and Consciousness by Naotsugu Tsuchiya。レビュー中ですが、レビュー形式に後述の(3) wiki-style peer-review processを選択してます。
ちなみにレビュー形式はいろいろ選べるようになっているようです。いまのところ3パターン見つけました。(1) "old-fashioned peer-review process: all comments should be put into the 'reviews' part of the article" (2) "mixed-mode peer-review process: minor changes in the article, major suggestions in the 'reviews' part" (3) "wiki-style peer-review process: reviewers are encouraged to modify the article directly"です。
さらにまだこれからできる項目も含んだリストを見ると、どんどん大きくなっていくだろうことが予想できます。これは良くなるのではないでしょうか。現状はまだ記事ができあがってきたという段階で、更新やレビューなどが活発になるところでまではきてないようですが。
たしか元々wikipediaはレフェリー付きでやったらうまく広がらなくて、だれもが編集できることによって爆発的に広がったという経緯があったはずですが、アカデミックな話題に関してはscholarpediaのようにやったほうがうまくいきそうに思います。ともあれ、このような形でフリーな記事ができるということは素晴らしいことです。ライセンス形態はちょっとよくわからんですが。
2007年11月08日
■ SFNから帰ってきました
San Diegoから無事帰ってきました。いま成田から名古屋の接続待ちです。
ぶじ、体調も崩さずに発表も終えてきました。
到着した晩が神経科学者SNSのオフ会でした。そこでいろんな方とお話を。神経科学者SNSの活用法として、nhpの電気生理についてのノウハウの意見交換をclosedでやるのはどうだろう、とか熱弁を振るったり。
ネット関係では、Shuzoさんやしげさんとお会いして、局所回路ネットワークの解析にさいきん大きなブレークスルーがあったかどうか、というような話を聞きました。あと、ネットワークの動態の記述に局所的な要因で説明したり(Laurrentのバッタの話みたいに薬をかけて発火頻度は変えずにoscillationだけ変えるといったトリック)、localなpertuabationを加えることで説明したりする(海馬スライスのカオスの制御の話とか)のはネットワークの記述じゃなくて局在論へ戻ってるだけではないか、とか言ってみたり。でもそういう感じで「秘孔を突く」というのはネットワークのなんつーか特異点みたいなところをidentifyするということであって、それでいいのかも、とか考えたりとか、やっぱり熱弁。
なんか感化されて、Buzsakiの"Rhythms of the Brain"を買ってきました。ちなみにネットの各所でもレビューが出てます。金井良太さんのサイトのエントリ。わるねこさんのサイトのエントリ。ykenko1さんのサイトのエントリ。
いろんな人と会って、将来の計画を画策。論文出す方が先決なんですが、なんだかんだと生理研研究会とか神経科学大会シンポジウムとか招待口演とか計画練ってます。
発表の方は無事済みました。スライド、原稿の準備、しゃべりなど神経科学大会より向上したのではないかと思います。Pptのアニメーション機能で「アピール」機能使いまくり。全体的に詰め込み気味だったのですが、だいたい10分で話が終了。もっとゆっくりやっても良かったみたいです。
質問の方はダメダメ。二番目の質問で何言ってるかわからなくて言葉尻捉えて関係有りそうなことしゃべってから、"am I answering your question? "とか聞いたら、"No, but it's ok"とか言われてがっくり。あとでSchmid(後述)とかとしゃべっているときにその話題が出て、要するにlesionがperfectだったかどうか聞きたかったということらしい。もちろんそれに対する答えはあったのだけれど。というわけでやっぱり英語に関してはダメダメ。今年はとくに滞在が短かったということもあるけど、とくにダメだった気がします。ヤバイ。
セッションは"eye movement: physiology and behavior"を選びました。視覚のオーディエンスにも聞いてもらいたかったけど、上丘のニューロン活動に詳しい人に聞いてもらいたかったということでこちらを選択。Stefan EverlingとかBrian Corneilとかがすごく面白がってくれたので良かったのではないかと。あと、Alex Maierから話を聞いて聞きに来たNIMHのLeopold研のMichael Schmidがトークのあとでやってきて話をしましたが、かなり関係があることやってます。急がなくては。Stefan EvalingとかMichael Schmidには帰りにラボに寄って話をしないか、と言われたけど次の日帰らなくてはいけなかったので泣く泣くまた次回に、という話に。
あと、Vikingさんも見に来てくれて、少し話をしました。じつは今回が初めて。
Laurent Ittiとのコラボレーションの方の仕事もぶじ発表が終わって安心。LaurentはSFNははじめてかもしくは久しぶりだったらしく、あまりにスライドセッションが多すぎて、個々のセッションのスコープが狭くなってしまっているのを嘆いていました。どの口演に関しても質問があまり出ないし。たしかに彼が本拠としているVSSはconcurrent sessionがたった二つで、みんなが同じセッションを聞いているという一体感があります。これはASSCも同じですが。やっぱあんまり規模が大きくなるのはいいことないと思うんですけどね。
そんなこんなで無事帰ってきました。
# Shuzo
オフ会でお話できて良かったです。
ネットワーク関連の議論ですが、局在論というご指摘、確かにそうかもしれません。ただ、manipulateすべきものをidentifyする、というのは、何を研究するにしろやはり重要な気がします。そして、因果関係的なことにも迫れる時代が来るかも?という期待が持てるだけでも一歩前進のような気がしてます。(と生意気こいてみます。。。)
ところで、ブザキ本を買われたとのこと。
哲学書のように、「ここ、書いたブザキ以外わからんのでは?」と思える超難解な箇所が多々ありますが、なかなか楽しめる本だと思います。人によってつかれる「秘孔」が違いそうですが、個人的には、後半の海馬関連の話をどのように思われるのか(どれくらいフェアか)、ぜひpooneilさんにお話をお聞きしたいところです。
また直接お会いできるのを楽しみにしています。
オーラル発表、お疲れ様でした。
こちらこそ、ほんの少ししかお話できませんでしたがpooneilさんにお会いできて嬉しかったです。神経科学SNSのオフ会は、諸事情により参加できず残念でした。また皆さんとお酒片手にお話できるような機会があればぜひその時は参加したいと思います。
SFNはどんどんオーラルの細分化がきつくなって、良くないですね。3年前に参加した時にも聞いた声ですが、「じっくり議論したいからあえてポスターにした」という人が増えるのも道理かと思います。もっともポスターはポスターでいい加減な発表も少なくないので、多少S/N比が悪いような気もしなくもないのですが・・・ポスターもポスターで半日よりは1日欲しいぐらいなんですが、今の発表件数だと致し方のないことですね。次回からは何か改善がなされるよう期待しております。
# pooneil>Shuzoさん
ブザキ本、まだちらちら最終章(13章)だけ読んだところですが、これすばらしいですね。わたしが考えてることとけっこう近いことが書いてあるのを見つけて、いろいろ焦ったり。このへんは自分の仕事の進行に従って、自分の現在の構想について追々書いていきたいと考えております。
あと、海馬関係ですけど、11章のうち、episodic memoryの項だけ読みました。Episodic recallとcued recallとをきっちり分けた上で、episodic recallに先立つspontaneous phase processionを探せばいいかもしれない、みたいな仮説を提示しているところは面白いのではないかと。以前から書いてますが、episodic memoryの問題は最終的には意識の問題ですので(episodic memoryは、"mental time travel"とでも言うべき追体験=主観的経験、が伴うのがその条件のひとつ)、ただのcued-sequential memory以上のものであるepisodic memoryを特徴づけるものはなんなのか、将来のネタのひとつとして興味持ってます。
あと、私の名前の入った論文がreferされてて喜んだり(どういう文脈でreferしているかはいまのところ不明ですけど)。Varela and Thompson 1991やO'Regan and Noe 2001がどういう文脈でreferされてるかもこれから検証です。
>vikingさん
あまりゆっくり話が出来なくて残念でした。お客さんが待ってたので話をさっくり切ってしまった次第。
このへん、ポスターセッションの時もいつも悩ましい問題です。たとえば、ポスター聞きに行って話をしてたら、途中でポスタープレゼンターの知り合いらしいのが割りこんで来てこっちは放置されて挨拶と世間話が始まって、苦々しくそのポスターから離れていく、こんな経験を何度かしたことがあります。(そういうわけでわたしはポスター聞くのがなんか不得意です。人混みの出来たポスターに割り込んで質問をするということがある時期から出来なくなりました。) そういう意味でも大変心苦しいことでした。
ちなみにオーラルセッションはいろいろ問題有るけど、より多くの人にアピールするためにはこちらの方がよいと思っています。ただし、どのセッションを選ぶかがものすごく重要でして、これを誤ってなんか泡沫候補みたいなところに集められたりするとほんとがっかりします。
論文出さないとけっきょく人は集まってくれないし、論文出すためにたくさん人に集まってほしいし、という鶏が先か卵が先かって話なんですけど、けっきょく論文を出し_続け_てプレゼンスを上げていくしかないというのが結論です。わたしは途中でsubjectを変えたのでこのへんもういちど積み直しというところがあって、なかなか苦しいところです。なんかイギリスのロックミュージシャンがアメリカ進出のために地道にアメリカ全国ツアーをするのと重ねてみたり。
それではまたの機会に。
tough problemsから取り組まれたとは。恐れ入りました。
エピソード記憶の部分に対するコメント、さすがなコメントで勉強になりました。ありがとうございます。コメントを読みながら、最近RedishたちのグループがJNSで報告しているCA3での面白い現象を想起しました。その論文、まだしっかり読んでませんが、pooneilさんが指摘していただいた文脈で読んでみるとさらに面白いかも?と思いました。今デスクの上にあるので、読んでみます。
ちなみに、pooneilさんたちのお仕事、p338でしっかり紹介されてますよ。
2007年10月29日
■ SFN2007が近づいてきました
SFN2007が近づいてきましたね。いつのまにかitinerary plannerのダウンロードページも出来てました。
で、ダウンロードして使ってみましたが、これがクソ遅い。立ち上げるとなんかえんえんデータ読みこんでるんですけど。なにこれ。全部のデータベース読み込んでるんでしょうか。アホか。いいからとりあえず検索画面出したり、saveしたitinerary読むとかそういうことしてからバックグラウンドでデータベース読み込むとかすればいいのに。この世界にはフリーのキラーアプリとかないんですかね。
以前学会のオンライン登録システムでフリーなやつとかを探したことがあったんですが、どの学会でも同じようなことやってるのになんとかならないのかなと思います。
ぱらぱらアブスト見てますが、皆さんいろいろ進展しているなあ、と感慨深い。わたしもこうしちゃいられません。
ちなみにわたしは6日の午前に発表です。Slideにチャレンジで前々回(2005年)のリベンジ。ていうかけっきょく英語勉強してきませんでした。だんだん焦ってきました。内容自体は神経科学大会と同じだけど、まだいろいろ改良できるところがあるのでギリギリまでいじることになるんだろうなあ(他人面して)。
San Diegoへは4日午後に到着、その晩は神経科学者SNSのオフ会。次の日5日の昼も向こうの共同研究者と会って議論。夜は前ラボの飲み会に行くか、いま画策している別系統で行動か。次の日6日の午前に発表が済めばやっと一息、でも次の日7日の早朝にSan Diego出発。いつも通り強行スケジュールですが、在米の方やご無沙汰している方とお会いできるのを楽しみにしております。
2007年10月26日
■ MindPapers
David Chalmersのサイトにオンラインで読めるconsciousness関連の論文のリストというのがあったんですけど、それがexpandした形で"MindPapers"というサイトになった、というのがASSCのMLで流れてきました。
"What is it Like?"の項目だけで論文が35個あったりするともう気が遠くなってしまいます。俺たちは上り始める。 長い、長い坂道を、って調子で上ってるとこのまま人生が終わってしまいそうなので、自分ができることを何とかやっていきます。
……この一生で「カラマーゾフの兄弟」は読めないかもしれない。「知覚の現象学」や「物質と記憶」は読んでおきたい。「存在と時間」にはたどり着けないかも。とか考えたりします。いかんいかん。
ではまた。
2007年10月24日
■ Vocaloid2と不気味の谷、そこからリアリティーの研究へ。
Vocaloid2ですが、けっきょくそのあとやっとこさ届きまして、ぽちぽちいじってます。ネット世界ではその間にさまざまな話題がすぎてゆきましたが、それはほぼスルーで。黙って自作曲作るべし、ということで。
でもひとつメモっておきたくなったのが、「不気味の谷」の概念との関連について。
「不気味の谷」wikipedia日本語版、英語版
The "Uncanny Valley" Masahiro Mori(原著の英訳)
3Dに関しては、リアルな造形のアンドロイドが不気味に見えるという例で、阪大石黒研のアンドロイドの例がいちばんわかりやすいですね。「不気味の谷」の概念が正しいかは別としても、これは怖い。ついでにGIZMODOのムービー。
2Dに関しては、CGでリアルに造形した顔が怖いというやつで、WIREDの記事とか。
けっきょく人間の認知として、顔の認知はとても感度が高い(つまり、顔に関する微妙な違いに気付く能力は他のもの、鳥とかテレビとかの弁別と比べてものすごく高い)ので、なんらかの不自然さを読み取ってしまうのだと思います。それはいったいなにか。このへんは脳科学が出来ることがあるんではないかと思うんです。顔認知に関する議論はここでもいくつか扱いました。(ざっくり考えればモーフィングで刺激作って現象が定量的に再現できればいいわけですが、要は完全にリアルになる手前に安定なポイントがあるということに意義があると思うんです。)(つーか、wikipedia英語版の方を見ると論文ありますね。あくまでrobotics側からのものみたいですけど。)
たぶん声の認知に関しても、視覚的な顔の認知と似たようなことが起こっていると考えられます。だから、vocaloid2について不気味の谷のことを考える意義があって、いま多くの人がそのことに気付いています。
CNET blog
注文の多い古書店
たろの日記ページ
END_OF_SCAN
「セキュリティ&コンサドーレ札幌」(vocaloidの話ではないけど、「手前の山」に言及してます)
んで、「キャラクター性を抽出した、ある抽象度の、表現のレベル」(あるキーワードを回避するためまわりくどい)によってその谷の手前の山が大きくなっているということなんではないか、というわけです。これってもしかしたらそういうキャラクター性を取り扱う糸口になるんではないか、脳科学として可能なことがあるんではないかと思っているのです。これが将来のネタのひとつになるんではないかとけっこうまじめに考えてます。以前から冗談めかして吹聴してますけど(20051115のエントリとか)、どのようにしてキャラクター性が生まれるのか、ということは意識研究と繋がってくるはずです。
つまり、意識の研究は「何を見ていると思っているのかについて、どう信憑性を確保しているのか」という側面からアプローチすることによって「リアリティー」の問題に繋がると考えています。つまり、たんなる受け身のvision (こっちはventral pathwayが主役)では、意識経験を外界によるものと幻覚とを区別することはできない。それらを区別するためにはverificationの可能性を持っているかどうかが重要であり、そこにsensorimotor contingency(こっちはdorsal pathwayが主役)が関わってくる、というのが現在のアイデアです。ある物体に実際に触れて確かめなくても良いのだけれど、確かめる手段を持っていること、つまりその可能性を持っているかどうか、が意識経験とリアリティーに重要であるだろうと考えています。夢は意識経験か、とか桶の中の脳問題とか、開眼手術後の視覚とかはこのラインで考えられないかと思うんです。(こういう重要なことをこういうエントリに差し込んでみる。)
元の話の方をもう少し展開させますね。顔認知においてこのような不気味の谷がある一方で、biological motionの研究からもわかるように、われわれは単なる点の動きからでも人間や生き物の動きを読み取ることが出来ます。ここは面白い対照だと思います。これはけっして矛盾ではないと思います。つまり、たぶんどっかで図と地の関係が反転するのですな。人間ぽい要素の方が大きくなってきてそっちが地になって、そのなかに混じったロボっぽさが図となって目立つときに気味悪さが出てくる。音声に関しても、以前のボコーダー的な発声の場合はロボっぽさが地だったと思うけど、vocaloid2は人間っぽさが地になって、調整が下手だと出てくるロボっぽさが図になり、気味悪さが出てるのだけど、キャラクター性によって補われている。そのようなキャラクター性による補完ができない人が、「ぜんぜん人間らしいと思えない」と言ってる、ってのが現在の状況なんではないかと思うんですけど(スルーできてないし)。
たしかにここにキャラクターが介在していなかったら、けっこう気味悪かったかもしれません。んで、これはいったい手前の山が高くなったということなのか、それとも谷を乗り越えたのか、ってのがひとつの論点です。たしかに、顔認知の類推でいけば、谷の手前としか思えない。でも、これから音声合成の技術が上がってさらにリアルなものができたときに気味悪さが生まれるかというとそうとも思えない。谷を乗り越えられる人と乗り越えられない人がいるように思います。わたしは乗り越えちゃいました。
ちなみに、実際に使ってみればとそれなりにわかるもんなのですが、歌詞を入れないで「アー」だけで旋律を作るとロボまる出しなんです。日本語を入れるとそれが一挙に人間らしくなります。この過程は面白い。さらにロボっぽさを消すために細かくいろいろ設定する必要があるのですが、わたしはそちらにはこだわらずに使用しております。
たぶん、将来的には音声合成のレベルはもっと上がる(もっと平板さが少なくなるとか)のだろうけど、それは製作過程でロボっぽさが消えるのと、細かい調整の手間が減るという方向であって、本質的にはそんなに変わらないんじゃないか、そんなふうに予想してます。つまり、ボコーダー的な使用法からいまのvocaloid2での使用法というところ(上記の図と地の反転)がやっぱりいちばん大きかったんではないかと思うのです。ちなみに図と地の反転がこの不気味の谷間とは別もんであるだろうことは、2D,3Dでの顔の造形について考えれば明らかです。不気味の谷自体は図と地の反転が起こってからの現象であるとは言えるでしょうけど。
ではまた。
(ちなみにこの商品名を一回も書いていないのは、検索でひっかかんないようにということと字面のバランスを取るためです。回りくどいエントリでした。)
12/19追記:稲葉振一郎氏のエントリでも関連した話をしているのを発見。
2007年10月07日
■ ASCONE無事終了しました
ASCONE無事終了してかえってきました。とても楽しかったです。感想などもろもろについてはまた研究会の後ということで。
2007年09月12日
■ 神経科学大会終了
発表の方は無事終わりました。まあ、練習に時間をかけた分くらいは伝わったのではないでしょうか。伝えることをメインに、かなり削りました。トークで詰まってしまうことがないようになんとかセリフ全部記憶してゆっくりしゃべるよう心がけました。
いつもできないのが、節目で一息つくとか、観客全体を見渡すとか、緩急を付けること。ようするに、スクリーンばっかり見っぱなしにならないようにしたいんですけどね。やっぱSteve Jobsリスペクト。SFNのときこそうまくやるつもりです。ちなみにSFNのトークもsaccadeのセッションです。Visionのひとの方が面白がってくれそうなのだけど、データは上丘ニューロンの活動なもんで、苦渋の選択です。
あと、オチをうまく付けたいんですけどね。"Thank you for your awareness of my talk, thank you even if it is a random choice for you"とか言ったらクドいですかね。
あとでポスター会場で福島研に所属しているガイジンさんに、おまえのトークを見たけどジェスチャーが激しいので日本人じゃないのではと思った、と言われて苦笑い。
んで、ラーメン二郎 横浜関内店と吉村家本店に行ってきたという出張でした。ラーメン二郎のほうは、「汁無し」を頼みそびれたので(店に入る前に指定する必要があったのに気付かず、ふつうの小ぶたになった)、明日また行って頼む予定。ラーメン博物館の方はいいや。
明日はplexonの講習会。まわりのひとはかなり導入しているようですけど、わたしはまったくわかってないんで基礎的なところからです。
2007年08月30日
■ ラボでの論文コメントの例
どもども。けっきょく皆既月食の方は雲が厚かったですが、何度か雲の隙間から見ることが出来て満足しました。いちばん楽しいのは皆既月食が明けて月が光り出す瞬間だと思いますが、そこは雲が厚くて見られず。家族と一緒に見たけどいちばんはしゃいでいたのが私だったとかそういうオチ。それもまたよし。
こないだのエントリで
ウェブを漁るといろんなラボのジャーナルクラブの記録(論文リスト)が出てきます。あれに「要旨見てもわからないようなこと」を3行コメント付けてくれればすごく有益だと思うんですけど。
なんて書いてましたが、富山大の田村先生のところを見てたら「富山大学生理学徒然記」というブログ形式でのセミナー記録を発見。すかさずリンクしておきます。ここは記憶関係メインですが、ラボでやってることを反映してnonhuman primatesだけでなくrodentsもあって、よいです。
そういえば、霊長研の三上先生のラボのサイトにある「脳の世界」、ここもいろいろな論文が採りあげられていて、私もあんちょことして使いました。たとえば、Sommer and Wurtzをセミナーで採りあげたときには「これから行う運動を感覚系に知らせる」のお世話になりました。
2007年08月23日
■ ASCONE2007で講師をやります
ASCONE2007で講師をやることになりました。告知が遅れてしまいましたが、今日が参加応募の締め切りの日でした。
ASCONEは日本神経回路学会および統合脳のサポートによるオータムスクールです。若手の研究者が講師になって、参加者には講義を聞いてもらうだけでなく、演習を通してよりふかく積極的に参加していただくというものです。
昨年のASCONE06については統合脳のサイトに概要や参加者の感想などを含んだ開催報告(PDF)がありますので参考になると思います。
今年は『知覚と運動を結ぶ脳機能』というテーマになっています。わたし自身はまだ現在の仕事が論文になってませんし、正直自分で良いんだろうかと思うところはありますが、引き受けたからにはきっちりやりたいと思いますので、参加者の皆さま、よろしくお願いします。
2007年06月15日
■ 「視知覚研究の融合を目指して-生理、心理物理、計算論」参加しました
生理研で毎年6月頃に行われている視覚研究会「視知覚研究の融合を目指して-生理、心理物理、計算論」に参加してきました(プログラム:PDF)。昨年は発表させていただきました(20060609にさらっと記載)。
今回の演題でいちばん印象深かったのは産総研のシステム脳科学研究グループの小村さんの仕事でしょうか。いまのところ視床のattentional modulationという印象の仕事ですが、実験デザインがかなりかっちりしていてデータが強烈。これもかなりいいとこ行きそうです。
懇親会は例年通り、立食形式の後にとなりの座敷部屋で10時まで語りモードで。これいいやり方だなあと思います。どこか飲み屋へ移動するとそこで研究の話が切れてしまいがちですが、座敷形式だと研究の話をする雰囲気が持続するのです。これは見習いたい。統合脳の班会議でも、以前の長野の松代での会では立食形式の懇親会の後に大きな座敷の部屋に丸テーブルが10個とかそんなかんじの部屋に移動して語り合ったり、さらにホテルの部屋になだれ込んで夜遅くまで語り合ったりという機会があってとても良かったです。札幌や東京ではちょっとそういう機会がないので残念。ようするに私は「合宿」がしたいのですな。いい年して言うのもなんですが。(自分はことし、二十七になります。白髪がめっきりふえたので、たいていの人から、四十以上に見られます。ウソ。)
研究会終了後はラボツアー開催。うちのラボをぐるっと回ってもらってから、最後に私が以前発表したポスターを前にして電気生理データを説明して強制的に議論モードへ。(パックのツアーで土産物屋に連れていかれるのと同じみたいな。) そこでいろいろアドバイスをいただきました。どうもありがとうございます。これからこれをとりこんで来週のASSCポスターの作成です。
生理研のメリットはいろいろありますが、ひとつは研究会を開催して各地各分野の方を議論する機会を持つことが出来ることです。今回も同年代の神経生理学者たちとお話しできるだけでなく、心理物理の方と議論が出来たのは大きな収穫でした。私のポスターのROCカーブの説明で「y=-xの軸で対照でないからunequal variance modelで考える」みたいな話が一発で通じてしまうのにほとんど感涙。いや、プロにとっては当たり前なのでしょうけど。ホワイトボードが真っ黒になりました。
ちょっとテンション上がり気味で書いてみました。かつては書いてから数日おいて冷静になって見直してから載っけてたのですが、そういうことしてるとそもそもエントリを書かなくなることがわかったのでちょっと勢い付けぎみで。
2007年06月08日
■ 神経生理必読文献リスト作ってみました
神経生理必読文献リストを作ってみました。1980年代までのもので、non-human primatesでのsingle unit studyで読むべきだろうというものについてリストしてあります。初期のものについてはfrogだったりcatだったりしますが。全部読んでいるわけではありません。それから、どうしても分野が偏ります。いまのところvisionとsaccadeがメインです。追加を歓迎します。RabbitだけどH Barlowとかも入れるべきか。初期の方はどこがいちばんのoriginかというと難しいです。
おもにニューロン記録について作ってみましたが、同様にして、anatomyやlesion studyやhuman patient neuropsychologyなどでも作ることが出来ることでしょう。これらは歴史がありすぎて、私はフォローできません。そもそもフランス語だったりするし。逆にimagingは新しすぎてまだ作る自信がありません。
[1959 LETTVIN: Frog optic tectumのbug detector]
[1959 Hubel and Wiesel: V1のreceptive field]
[1963 Hubel and Wiesel: V1のocular dominance columnのdevelopment]
[1966 Evarts: voluntary movementとM1ニューロン]
[1967 Cross correlation analysis]
[1969 Fetz: operant conditioning]
[1971 WurtzとGoldberg: 上丘とサッケード]
[1971 Prefrontalでのsustained activity]
[1972 上丘のsensory mapとmotor map]
[1976 Sparks: population coding]
[1978 Mountcastle: コラム説]
[1981 Gross: face neuron]
[1981 Goldberg: LIPのattention theory]
[1983 Hikosaka: SNr]
[1985 Desimone: V4とattention]
[1986 Georgopoulos: population coding]
[1986 Shultz: dopamineと強化学習]
[1987 Richmond and Optican: firing patternの時間情報]
[1988 Miyashita: 側頭葉と長期記憶]
[1988 Merzenich: Somatosensoryのreoarganization]
[1989 Goldman-Rakic: working memoryとprefrontal cortex]
[1989 Newsome: perceptual decisionとMT]
[1989 Logothetis: binocular rivalryと神経活動]
[1959 LETTVIN: Frog optic tectumのbug detector]
J. Y. Lettvin, H. R. Maturana, W. S. McCulloch and W. H. PittsWhat the Frog's Eye Tells the Frog's Brain.
Proc. IRE 47 (1959) 1940-1951
MATURANA HR, LETTVIN JY, MCCULLOCH WS, PITTS WH.
Anatomy and physiology of vision in the frog (Rana pipiens).
J Gen Physiol. 1960 Jul;43(6)Suppl:129-75.
PMID: 13768335
[1959 Hubel and Wiesel: V1のreceptive field]
HUBEL DH, WIESEL TN.Receptive fields of single neurones in the cat's striate cortex.
J Physiol. 1959 Oct;148:574-91.
PMID: 14403679
HUBEL DH, WIESEL TN.
Receptive fields, binocular interaction and functional architecture in the cat's visual cortex.
J Physiol. 1962 Jan;160:106-54.
PMID: 14449617
HUBEL DH, WIESEL TN.
Shape and arrangement of columns in cat's striate cortex.
J Physiol. 1963 Mar;165:559-68.
PMID: 13955384
[1963 Hubel and Wiesel: V1のocular dominance columnのdevelopment]
WIESEL TN, HUBEL DH.SINGLE-CELL RESPONSES IN STRIATE CORTEX OF KITTENS DEPRIVED OF VISION IN ONE EYE.
J Neurophysiol. 1963 Nov;26:1003-17.
PMID: 14084161
WIESEL TN, HUBEL DH.
EFFECTS OF VISUAL DEPRIVATION ON MORPHOLOGY AND PHYSIOLOGY OF CELLS IN THE CATS LATERAL GENICULATE BODY.
J Neurophysiol. 1963 Nov;26:978-93.
PMID: 14084170
HUBEL DH, WIESEL TN.
RECEPTIVE FIELDS OF CELLS IN STRIATE CORTEX OF VERY YOUNG, VISUALLY INEXPERIENCED KITTENS.
J Neurophysiol. 1963 Nov;26:994-1002.
PMID: 14084171
[1966 Evarts: voluntary movementとM1ニューロン]
Evarts EV.Pyramidal tract activity associated with a conditioned hand movement in the monkey.
J Neurophysiol. 1966 Nov;29(6):1011-27.
PMID: 4961643
Evarts EV.
Relation of pyramidal tract activity to force exerted during voluntary movement.
J Neurophysiol. 1968 Jan;31(1):14-27.
PMID: 4966614
[1967 Cross correlation analysis]
Perkel DH, Gerstein GL, Moore GP.Neuronal spike trains and stochastic point processes. I. The single spike train.
Biophys J. 1967 Jul;7(4):391-418.
PMID: 4292791
Perkel DH, Gerstein GL, Moore GP.
Neuronal spike trains and stochastic point processes. II. Simultaneous spike trains.
Biophys J. 1967 Jul;7(4):419-40.
PMID: 4292792
Kimura M, Tanaka K, Toyama K.
Interneuronal connectivity between visual cortical neurones of the cat as studied by cross-correlation analysis of their impulse discharges.
Brain Res. 1976 Dec 17;118(2):329-33.
PMID: 1000297
Toyama K, Kimura M, Tanaka K.
Cross-Correlation Analysis of Interneuronal Connectivity in cat visual cortex.
J Neurophysiol. 1981 Aug;46(2):191-201.
PMID: 6267211
Toyama K, Kimura M, Tanaka K.
Organization of cat visual cortex as investigated by cross-correlation technique.
J Neurophysiol. 1981 Aug;46(2):202-14.
PMID: 6267212
[1969 Fetz: operant conditioning]
Fetz EE.Operant conditioning of cortical unit activity.
Science. 1969 Feb 28;163(870):955-8.
PMID: 4974291
Fetz EE, Finocchio DV.
Operant conditioning of specific patterns of neural and muscular activity.
Science. 1971 Oct 22;174(7):431-5.
PMID: 5000088
Fetz EE, Finocchio DV.
Operant conditioning of isolated activity in specific muscles and precentral cells.
Brain Res. 1972 May 12;40(1):19-23.
PMID: 4624487
Fetz EE, Baker MA.
Operantly conditioned patterns on precentral unit activity and correlated responses in adjacent cells and contralateral muscles.
J Neurophysiol. 1973 Mar;36(2):179-204.
PMID: 4196269
[1971 WurtzとGoldberg: 上丘とサッケード]
Wurtz RH, Goldberg ME.Superior colliculus cell responses related to eye movements in awake monkeys.
Science. 1971 Jan 8;171(966):82-4.
PMID: 4992313
Goldberg ME, Wurtz RH.
Activity of superior colliculus in behaving monkey. I. Visual receptive fields of single neurons.
J Neurophysiol. 1972 Jul;35(4):542-59.
PMID: 4624739
Goldberg ME, Wurtz RH.
Activity of superior colliculus in behaving monkey. II. Effect of attention on neuronal responses.
J Neurophysiol. 1972 Jul;35(4):560-74.
PMID: 4624740
Wurtz RH, Goldberg ME.
Activity of superior colliculus in behaving monkey. 3. Cells discharging before eye movements.
J Neurophysiol. 1972 Jul;35(4):575-86.
PMID: 4624741
Wurtz RH, Goldberg ME.
Activity of superior colliculus in behaving monkey. IV. Effects of lesions on eye movements.
J Neurophysiol. 1972 Jul;35(4):587-96.
PMID: 4624742
[1971 Prefrontalでのsustained activity]
Fuster JM, Alexander GE.Neuron activity related to short-term memory.
Science. 1971 Aug 13;173(997):652-4.
PMID: 4998337
Kubota K, Niki H.
Prefrontal cortical unit activity and delayed alternation performance in monkeys.
J Neurophysiol. 1971 May;34(3):337-47.
PMID: 4997822
[1972 上丘のsensory mapとmotor map]
Robinson DA.Eye movements evoked by collicular stimulation in the alert monkey.
Vision Res. 1972 Nov;12(11):1795-808.
PMID: 4627952
Schiller PH, Stryker M.
Single-unit recording and stimulation in superior colliculus of the alert rhesus monkey.
J Neurophysiol. 1972 Nov;35(6):915-24.
PMID: 4631839
[1976 Sparks: population coding]
Sparks DL, Holland R, Guthrie BL.Size and distribution of movement fields in the monkey superior colliculus.
Brain Res. 1976 Aug 20;113(1):21-34.
PMID: 821585
Lee C, Rohrer WH, Sparks DL.
Population coding of saccadic eye movements by neurons in the superior colliculus.
Nature. 1988 Mar 24;332(6162):357-60.
PMID: 3352733
[1978 Mountcastle: コラム説]
Mountcastle VB. An organizing principle for cerebral function. In: Edelman GM, Mountcastle VB, editors. The mindful brain. Cambridge (MA): MIT Press, 1978: 7-50.Mountcastle VB.
The columnar organization of the neocortex.
Brain. 1997 Apr;120 ( Pt 4):701-22. Review.
PMID: 9153131
[1981 Gross: face neuron]
Bruce C, Desimone R, Gross CG.Visual properties of neurons in a polysensory area in superior temporal sulcus of the macaque.
J Neurophysiol. 1981 Aug;46(2):369-84.
PMID: 6267219
Desimone R, Albright TD, Gross CG, Bruce C.
Stimulus-selective properties of inferior temporal neurons in the macaque.
J Neurosci. 1984 Aug;4(8):2051-62.
PMID: 6470767
Perrett DI, Rolls ET, Caan W.
Visual neurones responsive to faces in the monkey temporal cortex.
Exp Brain Res. 1982;47(3):329-42.
PMID: 7128705
[1981 Goldberg: LIPのattention theory]
Bushnell MC, Goldberg ME, Robinson DL.Behavioral enhancement of visual responses in monkey cerebral cortex. I. Modulation in posterior parietal cortex related to selective visual attention.
J Neurophysiol. 1981 Oct;46(4):755-72.
PMID: 7288463
Goldberg ME, Bushnell MC.
Behavioral enhancement of visual responses in monkey cerebral cortex. II. Modulation in frontal eye fields specifically related to saccades.
J Neurophysiol. 1981 Oct;46(4):773-87.
PMID: 7288464
[1983 Hikosaka: SNr]
Hikosaka O, Wurtz RH.Visual and oculomotor functions of monkey substantia nigra pars reticulata. I. Relation of visual and auditory responses to saccades.
J Neurophysiol. 1983 May;49(5):1230-53.
PMID: 6864248
Hikosaka O, Wurtz RH.
Visual and oculomotor functions of monkey substantia nigra pars reticulata. II. Visual responses related to fixation of gaze.
J Neurophysiol. 1983 May;49(5):1254-67.
PMID: 6864249
Hikosaka O, Wurtz RH.
Visual and oculomotor functions of monkey substantia nigra pars reticulata. III. Memory-contingent visual and saccade responses.
J Neurophysiol. 1983 May;49(5):1268-84.
PMID: 6864250
Hikosaka O, Wurtz RH.
Visual and oculomotor functions of monkey substantia nigra pars reticulata. IV. Relation of substantia nigra to superior colliculus.
J Neurophysiol. 1983 May;49(5):1285-301.
PMID: 6306173
[1985 Desimone: V4とattention]
Moran J, Desimone R.Selective attention gates visual processing in the extrastriate cortex.
Science. 1985 Aug 23;229(4715):782-4.
PMID: 4023713
Spitzer H, Desimone R, Moran J.
Increased attention enhances both behavioral and neuronal performance.
Science. 1988 Apr 15;240(4850):338-40.
PMID: 3353728
[1986 Georgopoulos: population coding]
Georgopoulos AP, Schwartz AB, Kettner RE.Neuronal population coding of movement direction.
Science. 1986 Sep 26;233(4771):1416-9.
PMID: 3749885
Schwartz AB, Kettner RE, Georgopoulos AP.
Primate motor cortex and free arm movements to visual targets in three-dimensional space. I. Relations between single cell discharge and direction of movement.
J Neurosci. 1988 Aug;8(8):2913-27.
PMID: 3411361
Georgopoulos AP, Kettner RE, Schwartz AB.
Primate motor cortex and free arm movements to visual targets in three-dimensional space. II. Coding of the direction of movement by a neuronal population.
J Neurosci. 1988 Aug;8(8):2928-37.
PMID: 3411362
Kettner RE, Schwartz AB, Georgopoulos AP.
Primate motor cortex and free arm movements to visual targets in three-dimensional space. III. Positional gradients and population coding of movement direction from various movement origins.
J Neurosci. 1988 Aug;8(8):2938-47.
PMID: 3411363
[1986 Shultz: dopamineと強化学習]
Schultz W.Activity of pars reticulata neurons of monkey substantia nigra in relation to motor, sensory, and complex events.
J Neurophysiol. 1986 Apr;55(4):660-77.
PMID: 3701399
Romo R, Schultz W.
Dopamine neurons of the monkey midbrain: contingencies of responses to active touch during self-initiated arm movements.
J Neurophysiol. 1990 Mar;63(3):592-606.
PMID: 2329363
Schultz W, Romo R.
Dopamine neurons of the monkey midbrain: contingencies of responses to stimuli eliciting immediate behavioral reactions.
J Neurophysiol. 1990 Mar;63(3):607-24.
PMID: 2329364
[1987 Richmond and Optican: firing patternの時間情報]
Richmond BJ, Optican LM, Podell M, Spitzer H.Temporal encoding of two-dimensional patterns by single units in primate inferior temporal cortex. I. Response characteristics.
J Neurophysiol. 1987 Jan;57(1):132-46.
PMID: 3559668
Richmond BJ, Optican LM.
Temporal encoding of two-dimensional patterns by single units in primate inferior temporal cortex. II. Quantification of response waveform.
J Neurophysiol. 1987 Jan;57(1):147-61.
PMID: 3559669
Optican LM, Richmond BJ.
Temporal encoding of two-dimensional patterns by single units in primate inferior temporal cortex. III. Information theoretic analysis.
J Neurophysiol. 1987 Jan;57(1):162-78.
PMID: 3559670
[1988 Miyashita: 側頭葉と長期記憶]
Miyashita Y, Chang HS.Neuronal correlate of pictorial short-term memory in the primate temporal cortex.
Nature. 1988 Jan 7;331(6151):68-70.
PMID: 3340148
Miyashita Y.
Neuronal correlate of visual associative long-term memory in the primate temporal cortex.
Nature. 1988 Oct 27;335(6193):817-20.
PMID: 3185711
Sakai K, Miyashita Y.
Neural organization for the long-term memory of paired associates.
Nature. 1991 Nov 14;354(6349):152-5.
PMID: 1944594
[1988 Merzenich: Somatosensoryのreoarganization]
Clark SA, Allard T, Jenkins WM, Merzenich MM.Receptive fields in the body-surface map in adult cortex defined by temporally correlated inputs.
Nature. 1988 Mar 31;332(6163):444-5.
PMID: 3352741
Wang X, Merzenich MM, Sameshima K, Jenkins WM.
Remodelling of hand representation in adult cortex determined by timing of tactile stimulation.
Nature. 1995 Nov 2;378(6552):71-5.
PMID: 7477291
[1989 Goldman-Rakic: working memoryとprefrontal cortex]
Funahashi S, Bruce CJ, Goldman-Rakic PS.Mnemonic coding of visual space in the monkey's dorsolateral prefrontal cortex.
J Neurophysiol. 1989 Feb;61(2):331-49.
PMID: 2918358
Funahashi S, Bruce CJ, Goldman-Rakic PS.
Neuronal activity related to saccadic eye movements in the monkey's dorsolateral prefrontal cortex.
J Neurophysiol. 1991 Jun;65(6):1464-83.
PMID: 1875255
[1989 Newsome: perceptual decisionとMT]
Newsome WT, Britten KH, Movshon JA.Neuronal correlates of a perceptual decision.
Nature. 1989 Sep 7;341(6237):52-4.
PMID: 2770878
Salzman CD, Britten KH, Newsome WT.
Cortical microstimulation influences perceptual judgements of motion direction.
Nature. 1990 Jul 12;346(6280):174-7. Erratum in: Nature 1990 Aug 9;346(6284):589.
PMID: 2366872
Salzman CD, Murasugi CM, Britten KH, Newsome WT.
Microstimulation in visual area MT: effects on direction discrimination performance.
J Neurosci. 1992 Jun;12(6):2331-55.
PMID: 1607944
Britten KH, Shadlen MN, Newsome WT, Movshon JA.
The analysis of visual motion: a comparison of neuronal and psychophysical performance.
J Neurosci. 1992 Dec;12(12):4745-65.
PMID: 1464765
[1989 Logothetis: binocular rivalryと神経活動]
Logothetis NK, Schall JD.Neuronal correlates of subjective visual perception.
Science. 1989 Aug 18;245(4919):761-3.
PMID: 2772635
Leopold DA, Logothetis NK.
Activity changes in early visual cortex reflect monkeys' percepts during binocular rivalry.
Nature. 1996 Feb 8;379(6565):549-53.
PMID: 8596635
Sheinberg DL, Logothetis NK.
The role of temporal cortical areas in perceptual organization.
Proc Natl Acad Sci U S A. 1997 Apr 1;94(7):3408-13.
PMID: 9096407
# nhk
これは素晴らしい。リストをみていろんな感想がわきましたが、一番強く感じたのは、当然ながら、キーパーソンというのは少数なんだなということです。何か追加した方がよいものに気づいたらまたコメントさせていただきます。
# pooneilどうもありがとうございます。偏りはしょうがないので、各界の方に補足していただきたいところです。
2007年04月04日
■ ポスター見に来てくれた人
無事Annual Meeting of the Society for the Neural Control of Movement(NCM)から帰ってきました。
学会としてはoculomotorよりはlimb movement系のほうが多い感じ。参加者は全体で300人程度。常時ひとつのシンポジウムかポスターセッションというかんじで、一体感があってよいです。以前の生理研COEシンポジウムとかがイメージとしては近いです。
わたしのポスターの方はほぼ常時だれかと話しているというかんじで、皆さんおもしろがってくれて、なかなか手応えを感じました。来てくれた人でバックグラウンドのわかる人をメモっておきました。日本人の方は割愛ということで。
- Brian D. Corneil@University of Western OntarioはDoug Munoz familyのひとり。R. A. Andersenのところでparietalのシグナルを使ったBMIの仕事(Science 2004 "Cognitive Control Signals for Neural Prosthetics")に加わった後でカナダに戻ってきた。以前からeye-head coordinationの仕事をしていて(Neuron 2004 "Visual Responses on Neck Muscles Reveal Selective Gating that Prevents Express Saccades")を出してる。
- Kevin JohnstonはStefan Everling@University of Western Ontario(こちらもDoug Munoz family)のところの人。最近Neuronを出したけど("Top-Down Control-Signal Dynamics in Anterior Cingulate and Prefrontal Cortex Neurons following Task Switching")、今回のポスターはJNS 2006 "Monkey Dorsolateral Prefrontal Cortex Sends Task-Selective Signals Directly to the Superior Colliculus"がpro- anti-saccadeだったのをmemory-guided saccadeにしたものを出してました。
- Neeraj J. Gandhi @University of Pittsburghは以前はDL Sparksのところにいた人。今回はeye-head coordinationのシンポジウムで、Vision Research 2001 "Experimental control of eye and head positions prior to head-unrestrained gaze shifts in monkey"の延長上の仕事として、上丘からの記録やOPNの刺激の話をしてました。この人は回路の人なので、私の話でどのくらい厳密に回路の議論ができているのか、というような質問が来た。
- H.H.L.M. GoossensはVan Opstal@Netherlandsのところのひとで、上丘のニューロン記録の論文あり:"Dynamic Ensemble Coding of Saccades in the Monkey Superior Colliculus"。
- Peter Bremenも同じラボで、外部への出力が不要な眼球運動測定法をポスターで出してました。Journal of Neuroscience Methods 2007 "Using double-magnetic induction to measure head-unrestrained gaze shifts. I. Theory and validation"。Van Opstal自身は来訪せず。いろいろ聞いてみたいことはあったのだけれど。
- Thomas Schenk@DurhamはもともとZihlのところでV5(=MT)損傷の神経心理をやってた人(Neuropsychologia 1997 "Visual motion perception after brain damage: I. Deficits in global motion perception")。今回は患者DFさんの話(Nature Neuroscience 2006 "An allocentric rather than perceptual deficit in patient D.F.")をポスターに出してたけど見逃しました。
- Pierre Baraduc@CNRSはWolpertのところにいた人で、reachingで内部モデル関連。Current Biology 2004 "Consolidation of Dynamic Motor Learning Is Not Disrupted by rTMS of Primary Motor Cortex"とか。この人はすごくこちらの話がよくわかってて、根掘り葉掘り質問されたけど、なんとか最終的には納得してもらえたと思います。
こう並べてみると、どういう人が客層なのかということがわかって収穫。場所柄か、比較的ヨーロッパの人が多い印象。なお、今回の学会はoculomotorに関しては上丘かその下流(OPNとかEBNとか)の人が中心で、FEF,LIPとかのひとはほとんどいませんでした。そのへんはたぶん7/8-13に行われるGordon Research Conferenceのoculomotor system biologyの方に流れてるのでしょう。こちらも参加したかったけどさすがにちょっと無理。
2006年12月27日
■ Corrigendumとerratumって違うの?
Nature Neuroscience 9月号に出た"High-resolution imaging reveals highly selective nonface clusters in the fusiform face area" Kalanit Grill-Spector, Rory Sayres & David RessのCorrigendumが出ました。ていうかCorrigendumとerratumと別枠であるんですね。知らなかった。wikipediaで見ると両者同じ扱いだけど、Nature Neuroscienceでこの9月号の目次を見ると、Corrigendumとerratumの両方があります。区別がわかりません。どちらもタイポのレベルの訂正だし。
Corrigendum: Nature Neuroscience - 10, 133 (2007) "High-resolution imaging reveals highly selective nonface clusters in the fusiform face area" Kalanit Grill-Spector, Rory Sayres & David Ress
もとの話はhigh resolution fMRIでFFAを調べたらfaceに選択性のある領域に紛れてnon-faceにも強い選択性を持つ領域があった。これまでの解像度ではそれは平均化されて見逃していたのだ、という話でした。
しかし、かつて一緒に論文を出したこともあるNancy Kanwisherのところから"Does the fusiform face area contain subregions highly selective for nonfaces?"が出てきて、脳の外を使ったノイズのデータでも同様なnon-faceのselectivityが出せてしまうことが指摘され、Alex Martinは"Measuring selectivity in fMRI data"でnormalizationの仕方が悪いのでselectivityをoverestimateしていること、さらにより適正なnormalizationを使って自前のデータを解析したらselectivityがなくなったことを示しています。
というわけで、Kalanit Grill-Spectorはオリジナルのデータのfig.4とfig.8は正しくないことをこのcorrigendumで表明したというわけです。他の結果は変わらないらしいけど、この論文のmain result (non-faceへのselectivity)をsupportするのがこのfig.4なので、ほとんど結論を撤回したとしか思えません。生きのこったfig.6-7はnon-face selectiveパッチの存在をサポートするものではありません。
ま、詳しく読んでみないと本当のところはわからないですけど、って恫喝訴訟に備えたかのようなフレーズを付け加えてみたりして(<-お!時事ネタ!)。
# ご隠居
ごぶさたです.投稿規定に説明が載っているようです.
以下引用.
CORRECTIONS
Errata: errors introduced by the journal during editing or production and for which the journal takes responsibility. Our policy is to correct such errors in cases where they distort the scientific meaning or the bibliographic record, or where they have significant potential to damage the reputation of the authors, the journal and/or third parties.
Corrigenda: errors introduced by authors, for which they take responsibility. Our policy is to allow authors to correct such errors in cases where they distort the scientific meaning or the bibliographic record, or where they have significant potential to damage the reputation of the journal or third parties.
Addenda: additional information from authors that, while not correcting a specific error, nevertheless alters the interpretation of a paper. Publication of addenda is at the discretion of the editor.
# pooneilあけましておめでとうございます。今年もよろしくお願いします。
なるほど、"Editorial policies" http://www.nature.com/neuro/about/ed_policies/index.html#corrections
のところに表記を見つけました。Erratumはpublisherによるエラーで、Corrigendumはauthorによるエラー、ということですね。知らなかった。
さらに探してみたところ、"Nature journals' correction and retraction policy"
http://npg.nature.com/npg/servlet/Content?data=xml/05_corrections.xml&style=xml/05_corrections.xsl
でCorrigendumなのかRetractionなのかの判断基準を見つけました。
"Retractions are judged according to whether the main conclusion of the paper no longer holds or is seriously undermined as a result of subsequent information coming to light of which the authors were not aware at the time of publication."
これを見ると、main resultであるかどうかが争点となるべきところであるように思いました。
2006年05月22日
■ 「Arational Agentの神経倫理学日記」はじまってます
これまでに当サイトにも記事を寄稿していただいているArational Agentさんがブログを始めています:Arational Agentの神経倫理学日記
認知神経科学と倫理学とをつなげる領域について詳しく書かれてます。
というか3/8へのコメント書き込みでさりげなくリンクされていたのにまったく気付きませんでした。
さいきんガザニガの「脳のなかの倫理―脳倫理学序説 」を読んだのですが、倫理的問題で神経科学の知見を踏まえずに過剰反応しているケースなどに神経科学者が警鐘を鳴らす、とかそんな論調でした。かなりざっくりと、どういう論点があるか指摘して終わる、ってかんじです。(追記:日本語を少し手直ししました。) Arational Agentの神経倫理学日記を見ると、すでにこの分野ではもっと深い議論がなされているようすがわかります。
ともあれ、期待しております。
# Arational Agent
お励まし、どうもありがとうございます。神経科学と哲学・倫理学の境界領域では、いろいろ面白い議論が行われているようなので、それらを調べて、紹介していきたいと思います。気長にお付き合いください。
2006年02月07日
■ Change blindnessとかapparent motionとか。
以前書きかけたものを貼ります。
Change blindness / Change detectionとかapparent motionとかについて調べてました。まずChange blindness / Change detection。
- "Neural Correlates of Change Detection and Change Blindness in a Working Memory Task" Luiz Pessoa and Leslie G. Ungerleider
- JNS '04 Wurtz "Subcortical Modulation of Attention Counters Change Blindness" せっかくやるからにはawarenessに絡む方が面白いのだけれど、attentionの話になってしまっている、という印象。
- Trends in Cognitive Sciences Volume 9, Issue 1 , January 2005, Pages 16-20 "Change blindness: past, present, and future" Daniel J. Simons and Ronald A. Rensink。この分野のオリジネーターか。Alva Nöeが表象関連について文句を付けている。Attentionの問題とかgapの問題とかかなり複雑で、扱いにくそう。
- Annu. Rev. Psychol. 2002. 53:245–77 "CHANGE DETECTION"(著者サイトのpdfファイル) Ronald A. Rensink。とくにわたしが気になっているのはchangeとmovementの問題なんだけど、けっこうさらっと抽象的に処理されているようすなので困る。どっかで扱われていると思うんだけれど、ちゃんとこの概念の始まりから追っているわけではないので見つからない。いや、端的にgapなしだとapprent motionが起こるのにgapありだとapparent motionが起こらないから、という説明が成り立つケースもあるのではないかと。
こちらはapparent motion。
- PLoS biology "Primary Visual Cortex Activity along the Apparent-Motion Trace Reflects Illusory Perception"
- Science '03 "Contributions of the Visual Ventral Pathway to Long-Range Apparent Motion"
- JNS '02 "Apparent Motion: Event-Related Functional Magnetic Resonance Imaging of Perceptual Switches and States"
2005年12月14日
■ 神経科学雑誌の投稿規定まとめ
まだ小休止。ryasudaさんのところに書いたコメント関連で。
投稿する雑誌ごとに論文の投稿規定があって、文字数が決まっているのだけれど、だいたいオーバーするものです(前回のエントリーにも書きました)。そこでどうするかというのが問題なのですが、投稿規定を守って文章削ってから投稿するか、必要なロジックと情報を全部入れて通ってから削ればよいとするか、ということになります。だいたいは大目に見てもらえるくらいにオーバーしている程度に必要なことは盛り込んで投稿する、ということになるでしょう。どのくらいオーバーしても大目に見てもらえるかの見積もりが人によって違いそうですけど。あと、オンラインの投稿システムとかで厳密に文字数が決まっているとこの技が使えないのだけれどどうなんだろ。
ということで、つい、投稿規定を調べてしまったのでいくつかメモ。2005年の12月13日現在ということで。もっといろんな規定はあるけど、原稿のスタイルについてだけです。ほかの雑誌について知ってる方は補充を。とくに最近投稿した人はオンライン投稿システムで気づいたこととか教えてください(上記の文字数などのソフトウェアレベルでの制限とか)。
[Nature]
Formatting guide: manuscript preparation and submission- They do not normally exceed 5 pages (One page of undiluted text is about 1,300 words.)
- no more than 50 references
- a summary, separate from the main text, of up to 150 words
- They do not normally exceed 4 pages (One page of undiluted text is about 1,300 words.)
- no more than 30 references
- a fully referenced paragraph, ideally of about 200 words, but certainly no more than 300 words
Article
Letters
-
[Science]
General Information for Authors いま気づいたけれど、ScienceはもはやMaterials and Methodsはsupporting online materialに入れる、と明示してあるのね。 - up to ~4500 words or ~5 journal pages
- up to 6 figures or tables
- a maximum of 40 references
- up to ~2500 words or ~3 journal pages
- up to 4 figures or tables
- a maximum of 30 references
Research Articles
Reports
-
[Nature Neuroscience]
Contents type - 2,000-4,000 words (excluding abstract, Methods, references and figure legends)
- no more than 8 display items (figures and/or tables)
- Abstract typically 100-150 words
Article
-
[Neuron]
Information for Authors - under 65,000 characters (including spaces, figure legends, and references)
- no more than 8 figures
- A single paragraph of fewer than 150 words
Article
-
[PNAS]
Information for Authors - six printed pages or 47,000 characters, including all text, spaces, and the number of characters displaced by figures, tables, and equations.
- Abstract. no more than 250 words
Article
-
[JNS]
Organization of the Manuscript - an abstract (not to exceed 250 words)
- an introductory statement (without heading; not to exceed 500 words)
- a description of the experimental procedures or methods, description of the results, a discussion of the experimental findings (not to exceed 1,500 words)
- References, limited to approximately 60
- ここは規定の遵守を厳しめに書いてます。"Submitting an incomplete manuscript or a manuscript that does not adhere to these limits will cause a delay in publication, and possible review."
Article
-
[JNP]
Instructions for Preparing Your Manuscript - Abstract: one-paragraph abstract of not more than 250 words
Article
ここまでで力尽きた。JNP要補充。Cerebral Cortex、PLoS biologyなども加えておきたいのだけれど。
2005年12月13日
■ 小休止で余談
線条体の話はまだ続きますが小休止。
月曜日にjournal clubで扱いました。SugrueのNature Review Neuroscienceの流れに組み込んで、SugrueのScience、Dorris and GlimcherのNeuronと併せてNeural correlate of valuationということでやってみたのだけれど、詰め込みすぎて失敗。Free-choice taskを使うことの意義(valuationとactionとの一対一対応を外す)と、その難しさ(valuationとactionとを分離することに起因する難点)とを説明したかったのだけれで、そこへ行く前にtaskの説明をするだけでいっぱいいっぱい。
これまた余談だけど、彦坂研のNature '02の本文の第一文はこう:"Reward shapes goal-oriented behaviour." めちゃかっこいい。「破戒」の出だしの名文「蓮華寺では下宿を兼ねた。」これに通じるような。文章が短くて、しかも主語、動詞、目的語の全部に情報があって、ムダな言葉(指示語)がないわけです。こういう締まった文章で始まる論文を書きたいものです。わたしはキャラ的にクドいもんで、こういうあっさりさに欠けるだけに、そうありたいなという気持ちは強い。
あと、NatureやScienceっぽい出だしでいいな、と思うのは、"Suppose that ..."とか"Imagine that ..."みたいなかんじではじめて、日常生活でだれもが持っている経験(記憶はあるのに名前だけ出てこない"tip-of-tongue"現象とか、時計に目を向けると秒針が一瞬止まったように見える現象とかね)を採りあげて、それのneural correlateを明らかにしました、みたいなやつね。
ムダだと思うのは、"It has been recognized that ..."とか"A number of studies have shown that ..."(って自分の論文かよ!)とか、情報がほとんどない前置き。
長くなるのはしょうがないところがあります。著者としてはいろんな断り書きを付けたいものですから。よくないんだけれど。科学論文はおもしろいことの書いてある読み物であるべきなんだけれど(NatureやScienceに関しては明確にそう)、いっぽうで法律の文書的に、あれは言った、これは言ってない、ということを厳密に記した文章なのですね。だからつい、あれこれ断り書きを付けだくなる。「われわれは現象Aがおこることを見つけた。」が「われわれは条件Bのときに現象Aを見つけた。」になって、「われわれは条件Bのときに現象Aを見つけた。条件Cのときには現象Dが起こることを見出した。」になって、この二文の関係をあたかも関連のある現象であるかのように見せるために(ロジカルには正しくないときに使うズルテク)、「われわれは条件Bのときに現象Aを見つけた一方で条件Cのときには現象Dが起こることを見出した。」とかにしてどんどん文章が長くなってゆくわけです。
まあ、すべてのことにはトレードオフがつきものであり、私たちはパラメーターが多すぎる問題を適度なタイムスケールである程度マシなやり方(思いついた限られた選択肢の中で選んだマシな方であり、最適化された、とは言わない)でもって処理する。そういうわけです。それがむずかしい。お、強化学習ネタ。
つれづれと。
2005年11月02日
■ 世界の神経科学者の系統樹 Neurotree
20051101のコメントで「V1の人」さん(敬称付けにくいよ!)からNeuroTreeについての情報をいただきました。どうもありがとうございます。
んでさっそくやってみました。おもしろい。たしかにまだ抜けはいろいろあるけれど、この分野の人だったらいろいろ遊べます。試しにやってみましょう。"browse as guest"から入れます。
たとえば、Bob Wurtzの下にはDoug Munoz、Michael E Goldberg、Marc A Sommer、William T Newsome、Okihide Hikosakaなどなど。そこから二つさかのぼるとDonald Hebbになってその上にKarl LashleyとWilder Penfieldがあって、下にはMortimer Mishkin、Brenda Milnerとか。Larry Weiskrantzのところは上にKarl Lashley、Karl H Pribramがあって、その下にはCharles Gross、Alan Cowey、Mel Goodaleとか。Vernon MountcastleからMichael M Merzenich、Ranulfo Romo、Richard Andersen、Apostolos Georgopoulosとかかなり壮観。同じように強烈なのはPeter Schillerの下にMichael Stryker、Larry R Squire、John Maunsell、Nikos Logothetis、Victor Lamme、Jeffrey Schall、Edward Tehovnik、Carol Colby、Marc Sommer、Robert McPeekというやつですな。MITってすごいなあ。一方で、Evartsの下にJun TanjiとPeter Strickというのは少なすぎるような。Wurtz門下の日本人も彦坂先生しか入ってないし。
じつは以前、こういうファミリーツリーを作ったらどうだろう、なんてことを言ったことがあったんだけれど、すでにこんなものが出来ているとはおどろき。じつのところ、こういうのってのは諸先輩方と酒を飲みながら昔話とともに伝えられていく性質のものだったりするのだけれど、こういうのまでネットにあるというのはすごいことになってる。それから、オタク的にもこういうのに単純に心が躍るのですな。60-70年代英米ロックの世界では、バンドのメンバーがどう行き来したかを独特の字体でチャート化したPete Frameのfamily treesというやつがありまして(ネットで見ることの出来るものとしては、たとえばdebris.com、ここから行けるJethlo Tullのfamily treeとか)、なんかそれに似通ったものを感じるのですな。ついででなんですが、カンタベリーミュージックシーンにかんしては、Collapsoでこのようなツリーが作られております。
{kind=link}
# tny
yama_tah blogさんのところからやってきました。はじめまして。
これは研究者の家系図なのですね?すばらしい!
私の分野は(といっても修士なのですが)タンパク質科学なのですが、
こちらでもぜひやってみたいです。いや、本当におどろきました。
どうもはじめまして。わたしも教わって見に行ったぐらいなのですが、なかなかおもしろいですよね。また、情報の入力はボランティアに依存している様子で、ムラはあるけれど、だんだん充実してくるというweb2.0的アプローチなので、これからどのくらい充実してゆくか見るのも楽しみです。
2005年10月08日
■ 髄膜炎
昔のブックマークを整理してリスト化シリーズ。
2005年10月02日
■ susceptibility artifact
以前チタンによるMR Imagingでのsusceptibility artifactにかんして調べたことがあって、そのときのブックマークが出てきたのでそれをリスト化しておきます。(ネタに詰まったので昔のブックマークを整理している、というわけです。)
- American Journal of Roentgenology, Vol 171, 1207-1213 Minimizing artifacts caused by metallic implants at MR imaging: experimental and clinical studies
- J Dent Res 82(8): 602-606, 2003 Artifacts from Dental Casting Alloys in Magnetic Resonance Imaging
- J Neurosurg. 2002 Dec;97(6):1472-5. Quantification of susceptibility artifacts produced on high-field magnetic resonance images by various biomaterials used for neurosurgical implants. Technical note.
- Can MRI be done on patient with implanted cerebral titanium clips.
- Magnetic Resonance Imaging of Periarticular Titanium Screws: The Relationship of Screw Position to the Ability to Image Articular Cartilage
- Magnetic resonance imaging guided musculoskeletal interventions at 0.23T
- MR parameters for imaging titanium spinal instrumentation
- Relation Between Magnetic Susceptibility of Non-ferromagnetic Dental Alloys and Metal Artifacts in Magnetic Resonance Imaging (MRI)
- Anterior Cervical Spine Fixation: Material Technology
- Pedicle Screw Segmental Instrumentation
- ご隠居の推薦を追記:The role of magnetic susceptibility in magnetic resonance imaging: MRI magnetic compatibility of the first and second kinds
# ご隠居
ちと物理ちっくですが,次のレビューは参考になるかもしれません.The role of magnetic susceptibility in magnetic resonance imaging: MRI magnetic compatibility of the first and second kinds.
http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=search&term=8798169
チタンを使っている限りは,多少の工夫をしても,厳しいのではと思いますが,なにかよい解決方法はありそうですか?低磁場(=<1.5)でなるべく強い傾斜磁場を使って高解像度でSEのT1強調画像を取るというのが一つの方向なのでしょうが,アーティファクトを減らす方向にパラメータをふるとS/Nも下がるというのも問題ですね.
>>チタンを使っている限りは,多少の工夫をしても,厳しい
うーむ、やっぱりそうですよね。SNを犠牲にせずに全体として減らすのはシークエンスを工夫しても難しいようなので、とりあえずはphase encodingの方向に気を遣って、肝心の部分にアーティファクトがこないようにするとかそのくらいの対応までとしているのが現状です。
2005年09月19日
■ 「本日のPubMed RSS feed」付けてみました
PubMedがRSS feedを出すようになってますね。検索の結果を"Send to"から"RSS Feed"を選択してやって、"Create Feed"を押してやると、その検索の結果の最新版を見るためのRSS FeedのXMLファイルのurlが出力されます。RSSリーダを使ってる人はそれで読んでやればいいわけです。
Movable Typeはプラグインを使ってRSS feedを読み込むことが出来ます。そこで、エントリの一番上に本日のPubMed RSS feedというのを作って、再構築するたびに更新されるようにしました。(cronを使って自動更新するというのは以前に失敗したので今回はやってません。) たまに検索キーワード変えたりしたらおもしろいかもしれないけど、ま、とりあえずテストということでやってみます。あと、アンテナ使ってる人が混乱しないように、新しいエントリを書いたときだけこのRSS feedも更新する予定。(追記:でも、いまのままだとエントリ更新するたびにRSS feedも更新されて、RSS feedの方がhtmlファイルの中では上の方にあるので、はてなアンテナで表示を「シンプル」にしている人にはRSS feedの方しか見えなくて不便かも。ソース上では更新エントリの方が上に来るようにしたいけど、それはまたの機会に。)
どんなふうにimplementされてるか:ほんとうはFeeds.Appを使うと良さそうなのですが複雑で扱いにくそうなので、よりシンプルなGetXMLを使用しました。GetXML.plをプラグインのフォルダに入れて、index.htmlのテンプレートに以下のコードを追加。
<MTGetXML location="http://eutils.ncbi.nlm.nih.gov/entrez/eutils/erss.cgi" rss_guid="0XZeiMI1_Dil4pkz70GAfj7ORTWgMjk13GfVSNAE3rV"> <MTGetXMLElement name="channel"> <ul> <MTGetXMLElement name="item"> <li><a href="<$MTGetXMLValue name="link"$>"> <$MTGetXMLValue name="title"></a> <$MTGetXMLValue name="category"> </li> </MTGetXMLElement> </ul> </MTGetXMLElement> </MTGetXML>
rss_guid=のところは検索キーワードなどによって適宜変える、という感じです。
しかし、XMLファイルを読んでみると、せっかくのページやら発行年の情報が取れなくて、全部"description"のなかに入っている、というもったいないやり方をしていることがわかります。authorも複数あるときは"authors"の下に一人一人のauthorを入れるようにとかすればよいのに、分けることができないし。とか細かい話はよいとして。 追記:勘違いでした。PubMedが提供しているのはあくまでRSSなので、itemのなかにtitle, link, descripptionがある、というのはRSSで決められた形式でした。私が言ったようなことはXMLとしてDTDとかを作ったうえでの話で、方向性が違うのでした。
2005年09月17日
■ Speed-accuracy tradeoff
ごぶさたしてます。なんか不調。
一般的に言って、急げばやることはいい加減になるし、正確にやろうと思ったら仕事は遅くなるわけです。このようにspeedとaccuracyにはトレードオフの関係があるので、心理物理学的な研究における課題の効果を評価するためには正答率だけでなく応答潜時も見てやる必要があります。たとえば、マスキングなり、注意課題のprecueなりが正答率に影響を及ぼしたと主張するときには、正答率が上がった分反応潜時が遅くなってないか、検証しておく必要があります。というわけで、このへん読まなくては。PubMedで検索したのを羅列しときます。
Vision Research Volume 35, Issue 20 , October 1995, Pages 2825-2843 Investigating simple and complex mechanisms in texture segregation using the speed-accuracy tradeoff method
Biological Psychology Volume 51, Issues 2-3 , January 2000, Pages 173-199 Mechanisms of speed–accuracy tradeoff: evidence from covert motor processes
Psychonomic Bulletin & Review, Volume 11, Number 2, 1 April 2004, pp. 353-361(9) One process is not enough! A speed–accuracy tradeoff study of recognition memory
Experimental Brain Research Volume 158, Number 4, October 2004, p415 - 42 Validity of the speed-accuracy tradeoff for prehension movements
NeuroImage Volume 22, Issue 4 , August 2004, Pages 1775-1783 Feedforward and feedback processes in motor control
Vision Research Volume 39, Issue 11 , June 1999, Pages 1943-1962 A comparison of the dynamics of simple (Fourier) and complex (non-Fourier) mechanisms in texture segregation
Vision Research Volume 44, Issue 12 , June 2004, Pages 1351-1365 Temporal performance fields: visual and attentional factors Marisa Carrasco
PNAS | April 24, 2001 | vol. 98 | no. 9 | 5363-5367 Covert attention accelerates the rate of visual information processing Marisa Carrasco
TheScientificWorldJOURNAL 2005/5 p128-146 Speed-Accuracy Tradeoff Operator Characteristics of Endogenous and Exogenous Covert Orienting of Attention
また、おそらくきっちり考えればこの話題は、リーチング課題での軌道の速度がベル型になるのは終止点の分散を最小にしようとしているからだ、というWolpertの説(北澤先生のNetScience Interview Mailとか参照)とかともつながるのではないでしょうか。
2005年09月01日
■ スキナー。あとデネットとか。
というわけで松代以来ぜんぜん新しいエントリを書いてないのですが、さいきんはスキナーとか読んで行動分析学を学んだり、信号検出理論あたりのこと勉強したり、といろいろやってます。
ところで、心は実験できるか―20世紀心理学実験物語 ローレン・スレイター
via 05.8 K.Moriyama's diary(8/27)
がスキナーについて書いているらしいですね。
んでもって、NewSphere経由で見つけた、スキナーの娘Deborah Skinner BuzanによるGuardianの記事。"I was not a lab rat"。センセーショナルなので内容の説明はしないでおきます。
さらにそれを受けてのThe new york timesの記事。
また、amazon.comでのeditorial reviewにはNew England Journal of Medicine, September 2, 2004からの記事があり。
というわけでいろいろ騒動があるようす。
なお、スキナーの娘が入っていた"aircrib"の写真はネットだとここで見つかります:認知行動科学(‘02)。もちろん、これはスキナー・ボックスではありません。
また、スキナーの自由意志の議論あたり(" Beyond Freedom and Dignity")はちょうどさいきん翻訳の出たデネットの「自由は進化する」と併せて読んでみる価値がありそうです。調べてみたところ、じっさい、デネットはスキナーに言及しており、Dennett, "Skinner Skinned," in Dennett, Brainstorms, Cambridge: MIT Press, 1978などがある様子です。
Web上で探しても、
BEHAVIORISM: SKINNER AND DENNETT
Dennett Indented? A commentary on Daniel Dennett's "Skinner Skinned".
Dennett and the Darwinizing of Free Will By David P. Barash ("Freedom Evolves"の書評)
AGAINST BEHAVIORISM DANIEL DENNETT SKINNER SKINNED (1978) IN: PROBLEMS IN MIND, CHAP. 6(googleのキャッシュ)
などが見つかってきます。

また、6/22にも言及しましたが、理研の松元さんによるこの記事からもいろいろたどっていけます。神経研の渡辺正孝先生の仕事についても指摘あり。(松代の班会議でも渡辺正孝先生は行動分析学的な枠組みでのおもしろい計画を提唱してました。)
また、松元さん自身のScience 03' "Neuronal Correlates of Goal-Based Motor Selection in the Prefrontal Cortex"での、刺激-行動-報酬の組み合わせを変えてprefrontal cortexでのニューロン活動のmodulationを見る、というストラテジー自身がスキナーの三項随伴性(弁別刺激-オペラント行動-強化刺激)の枠組みをもとにしていることがScience出版前に書かれた数理科学の「脳における主体性」で明らかにされています。
2005年08月25日
■ Cannabis and the brain
"Cannabis and the brain" Leslie Iversen, Brain, Vol. 126, No. 6, 1252-1270, June 2003
THE MOLECULAR LOGIC OF ENDOCANNABINOID SIGNALLING Daniele Piomelli, Nature Reviews Neuroscience 4, 873-884 (2003); doi:10.1038/nrn1247
"Cannabinoid physiology and pharmacology: 30 years of progress" Allyn C. Howlett, Christopher S. Breivogel, Steven R. Childers, Samuel A. Deadwyler, Robert E. Hampson and Linda J. Porrino, Neuropharmacology, Volume 47, Supplement 1, 2004, Pages 345-358.
"The endogenous cannabinoid system and the treatment of marijuana dependence" Daniele Piomelli, Neuropharmacology, Volume 47, Supplement 1, 2004, Pages 359-367. Nature Reviews Drug Discovery 3, 771-784 (2004); doi:10.1038/nrd1495
THE ENDOCANNABINOID SYSTEM AND ITS THERAPEUTIC EXPLOITATION Vincenzo Di Marzo, Maurizio Bifulco and Luciano De Petrocellis
"Long-term effects of exposure to cannabis" Leslie Iversen, Current Opinion in Pharmacology, Volume 5, Issue 1, February 2005, Pages 69-72.
"Emerging properties of cannabinoid medicines in management of multiple sclerosis" Gareth Pryce and David Baker, Trends in Neurosciences, Volume 28, Issue 5, May 2005, Pages 272-276.
2005年05月02日
■ Psyche-D-ML
20050425のエントリで言及したPsyche-D-MLの"Direct Perception and Representationalism"スレッドのまとめができた模様("Epistemological Debate" Steve Lehar)。読むならここからかな。参加者は話がループしだして疲弊している様子。Steve LeharはBehavioral Brain Sciencesに長年かけてターゲット論文を出した人。レビューの過程でのeditorの手紙を公開してたりしてて、見てる方はけっこうビビった(現在は見つからない)。
2005年04月25日
■ Psyche-D-ML
Psyche-D-MLが最近ものすごいactiveなんだけど("Direct Perception and Representationalism"スレッドとか)、まとめて読もうと思っていたらさらに爆発的に投稿されていて(最近一週間で130投稿されてます)もう読むのをあきらめました。J. Kevin O’ReganとAlva NoëのBEHAVIORAL AND BRAIN SCIENCES '01 "A sensorimotor account of vision and visual consciousness."(pdf file)とか取りざたされているようなので気にはなります。しかし、これだけ流れが速いときはだれかが煽っていると考える方が自然なわけですが、じっさいのところどうなんでしょ。
2005年01月06日
■ 「証明はできないけれどこれは本当だ、と信じていることってありますか?」
via http://d.hatena.ne.jp/umedamochio/20050105/p4 The Edge Annual Question—2005
"WHAT DO YOU BELIEVE IS TRUE EVEN THOUGH YOU CANNOT PROVE IT?"
Great minds can sometimes guess the truth before they have either the evidence or arguments for it (Diderot called it having the "esprit de divination"). What do you believe is true even though you cannot prove it?
意識と心の哲学関連あたり(Nicholas Humphrey, Daniel C. Dennett, William Calvin, Steven Pinker, Susan Blackmore, Piet Hut, Thomas Metzinger, Ned Block and Tor Nørretranders)がいちばん面白そう。
NeuroscientistはJoseph LeDouxやTerrence Sejnowskiが参加。Sejnowskiは長期記憶はシナプスではなくて細胞間隙のcytoskeltonに保存されているから50年経っても保たれるのだ、と大胆な説を。
2004年12月30日
■ カテゴリー別保管庫=おすすめスレッドリスト
これまでのエントリをサブカテゴリでまとめて読めるようにしました。随時更新されてゆきます。最新のものはreadmeページで読めるようにしてあります。- Paper[314]
- Carpenter's LATER model[6] 一般に行動の反応潜時は逆数を取ると正規分布する。これをニューロン集団が表象する意思決定シグナルが時間とともに蓄積して閾値に達するというモデルにおいて蓄積速度が正規分布する、というLATER modelをCarpenterは作り、説明をしようとした。
- Episodic-like memory[6] エンデル・タルヴィングの最新の定義によれば、エピソード記憶とは過去の出来事に自分の身をおいて追体験する"mental time travel"である。意識の存在を実証できない動物において、そのようなmental time travelは可能か、もしくはそれのアナログとなるものは存在しうるか。
- Glimcher '04 Neuron 神経経済学[16] 被験者対コンピュータでナッシュ均衡になるようなゲームをしているときの神経活動を記録すると、大脳皮質頭頂連合野LIPは選択肢の期待効用の比を表象していると著者は言うのだが…
- Newsome '04 Science 選択行動[9] ある選択肢のどちらを選ぶかを決めるときに、人間や他の動物はその選択肢の持つ価値(expected value)によって選択する確率を変化させる。大脳皮質頭頂連合野のLIPニューロンはそのようなexpected valueの比をローカルな時間スケールで計算したものを表象していると言う。
- Two-photon in vivo imaging[3] ニューロンの活動を電極でひとつずつ記録してゆく"single-unit recording"と、脳全体の活動を見ることはできるけれども個々のニューロンの動態はわからない機能イメージング(fMRIなど)、この両者が抱える限界を克服するために個々のニューロンの活動を広い範囲から光学的に同時に記録する方法論が発展してきている。
- ガヤScience article論文[14] 池谷裕二君がRafael Yusteのところに留学して行った仕事。大脳皮質の切片上で自発的に活動するニューロンの活動パターンはまったくのランダムというわけではない。正確に同じ発火パターンを繰り返す"repeating sequence"がある。
- 伝染するあくび[4] あくびはうつると言うけれどもこれは「共感」などの高次な機能が備わっていることで可能な現象で、小さい子供ではあくびはうつらない。今回、霊長研の松沢先生がチンパンジーのアイちゃんはあくびがうつることを発見した。
- 脳科学メモ[126]
- Chronostasis[2] 時計の秒針に目をやった瞬間、秒針がとまったように見えることがある。これがChronostasisだ。人間は目を動かしている途中には視覚情報が入ってこないように情報をシャットアウトしている。そしてシャットアウトされている期間、人間の脳は目を動かす直前まで時間を巻き戻しているのだ。
- Goodale and Milner[6] "The visual brain in action"において、視覚腹側路がperceptionを、視覚背側路がactionに関わる視覚処理を、というように分けて考えることを提唱したために論争が起こっている。その根拠となったvisual agnosiaおよびoptic ataxiaの症例をよく読みこんで検討する必要がある。
- オートポイエーシス[7] 人間が世界を見るとき、一人称の世界から環境と関係するといったある種閉じた状況としてしか捉えることができない。脳を明らかにすることで人間の心を明らかに使用とするときに見えてくる三人称的世界と一人称的世界とのギャップの問題を、あくまでシステムの中から見る立場を徹底することで克服しようと試みた体系がオートポイエーシス。だが、当時発展しつつあった自己組織化の概念と混ざりあってしまったために定式化の問題を抱えてしまった。そのエッセンスをなんらかの形で継承していくことができないか考えつづけている。
- クオリアについての「心の哲学」[9] Qualia-MLで村上/felineさんとともにStanford Encyclopedia of PhilosophyのQualiaの項目を読みながらあれこれと議論した記録。広義のクオリア、知識論法、逆転スペクトル、逆転地球、などの基礎的な事項がおさえられてます。
- 内部モデル、遠心性コピー、アフォーダンス[9] リゾラッティらが前運動野で発見した「ミラーニューロン」は「自分の行動遂行」と「他者の行動の観察」とでinvariantに活動するニューロンだが、これは頭頂連合野や上側側頭溝などとネットワークを作っている。Arbibがこのシステムをアフォーダンスと結びつけようと考えているが、少なくともオリジナルのJJギブソンによるアフォーダンス理論と脳科学とは食い合わせが悪いはずだ。
- 大学院講義「記憶の脳内機構」[24] 大学院講義「記憶の脳内機構」の準備のために記憶障害の患者さんの症例研究に基づいて宣言的記憶と非宣言的記憶との分類、エピソード記憶と意味記憶との分類について最新の知見までフォローする。
- 第一次運動野は何をコードしているか[2] 第一次運動野は何をコードしているか、という基本的なことですら現在でも論争が続いているが、StrickとScottの仕事でほぼ決着はついたといえる。Grazianoの仕事はそれに新しい切り口を付け加えたといえるが、microstimulationの特殊性と限界とを踏まえる必要がある。
2004年12月22日
■ SFNレポートつづき
上の論文関連でSEF関連の二つの発表についてメモしておきましょう。
- 378.4 "Separate representations of eye - and head - centered goals in the macaque supplementary eye field" J.Park, M.Schlag-Rey, J.Schlag
- -378.5 "Microstimulation of SEF can change the order of spatial memories" M.H.Histed, E.K.Miller
E.K.Millerのほうはdouble-step taskで、二つ出た刺激の位置に順番にサッケードする課題でdelayの時間に微小電気刺激(subthresholdで)してる。結論としては電気刺激によってサッケードする順番を間違えるということでした。質問で聞かれていたけどSchlag-Reyの結果に照らし合わせるとどうかというと、goal-directed仮説の方を支持する結果になっていて、刺激部位もSchlag-Reyでのposterior spotのほう(goal-directed)を刺激していたんだろう、という話になっていました。
- 378.6 "Attention and stimulus contrast modulate saccade latency but not kinematics" J.C.Martinez-Trujillo, A.Z.Khan, J.D.Crawford
2004年12月14日
■ Van Essen '91
の有名な図(cortexの領野がずらっと並んで結合関係が図示されているやつ)が欲しかったんだけどきれいなのがないので、しかたなく昔のCerebral cortexのコピーを使っていたのだけれど、
"Neural correlates of consciousness in humans" Rees G, Kreiman G, Koch C Nat Rev Neurosci. 2002 Apr;3(4):261-70.
これにmodifyされたきれいなのがあるのを確認。pdfをillustratorで開けばピクセルではなくてベクターで情報を持っている。
というわけで必要なときのために保存しました。
2004年12月09日
■ 川人先生
のトークがあったので行ってきました。前半は小脳の話と基底核の話ですでに論文になっているものです。何回か聞いた話ではありますが、いろいろと発見がありました。後半は今やってること、これからやっていることで非常にエキサイティングです。こちらに関しては書けませんが。
ATRのホームページhttp://www.cns.atr.jp/~kawato/にパワーポイントのファイルがいくつかあります。前半に関してはhttp://www.cns.atr.jp/~kawato/Ppdf/JCERE.pptが近い感じです。
それで前半に関して聞いてて考えたのは、どのくらいあらかじめ既知の条件を取り込んでモデル化してよいものなのだろうか、ということです。たとえば、階層型強化学習を使ってロボットが起き上がることが出来るようになるhttp://www.kawato.jst.go.jp/xmorimo/robo_sym2000.pdfという話の場合、それぞれの関節で行われる運動自体を強化するのではなくて、もっと上位のサブゴール(姿勢の高さ)が強化され、それによってこんどは下位の関節角などが強化される、という話なわけですが、たしかにそれは理にかなっているし、生物っぽい感じはするけれども、なんかそういう行動の全体を把握したホムンクルスのようなものが入り込んでいるように思えるのです。起き上がるという問題を達成するためにいくつかの問題に分割しなければならないのに、それをあらかじめモデルの中に取り込んでしまってよいのでしょうか。そうやって問題を分割する部分は未知のものとして、学習の過程で創発的に出てこないと意味がないようにも思えるし。しかしそのような上位の強化システムはいろんな行動で共通して使われているのだからよいのかもしれないし。考えれば考えるほどにわからなくなってくるのです。同じような感じで学習で獲得したいものがはじめからモデルの中に入っているというパターンをいくつかの点で見たように思うのです。
たぶんいま書いていることが飲み込めないと、複数のモジュールがあってそれぞれが内部モデルを持っていたり(Haruno M, Wolpert DM, Kawato M: MOSAIC model for sensorimotor learning and control. Neural Computation, 13 2201-2220 (2001))、それぞれが強化学習される(Doya K, Samejima K, Katagiri K, Kawato M: Multiple model-based reinforcement learning. Neural Computation, 14, 1347-1369 (2002))といったスキームの意義がわかってないことになるのだろうと思うのですが、まだ飲み込めていないのです。
というわけで少しNeural Computation '02のイントロを読んでみると、
"The basic problem in modular or hierarchical RL (引用者注:reinforcement learning) is how to decompose a complex task into simpler subtasks.
...
However, when the prediction models are to be trained with little prior knowledge, task decomposition is initially far from optimal. Thus, the use of "hard" competition can lead to suboptimal task decomposition."
と書かれているので、やっぱりどのくらいprior knowledgeを使うのはというのはnon-trivialな問題のようにも思えるし。どう分割するかの問題なのではなくて、すでに分かれているモジュールのどれが選択されるか、という問題を解けばいいだけなのかもしれないし(それがまさにこの論文で行われていることのようだし)。ああわからない。
# mmrl
なんかまた呼ばれた気がする....。疑問を解消できるとよいのですが。もちろん、問題の分割が学習で創発的にでてこないとダメだというのは尤もなことです。MOSAICの肝は予測誤差を使っていくつかの部分問題に分割するという点ですが、それだけでなく、学習についても言及していて、予測誤差がより小さいモジュールを強化する(学習する)ことによって、はじめは似通ったモジュールが分化されることもありうるわけです。 特に、Doya et al 2002はイントロの一部だけでなく、中身も見てほしいわけですが、Figure 8(a)に書いてあるのは最初の50試行は1つの状況(振り子の長さが一定)であるのに後半は2つの状況が交互に出現する(振り子の長さが可変)のような場合、1つの状況を担当していたモジュールが2つの状況を分割できるように分化しているわけです。 こういうことができるモデルだからこそイントロでprior knowledgeが少ないような場合、それだけでばっさり切る``hard’’ competitionは危ないといっているのです。だからって、使えるpriorを捨てるのは得策ではないし、prior knowleadgeと学習によってposteriorに与えられる情報とをうまく統合して状況分割する良い方法は、というと``soft’’ competition であり、baysianであり、MOSAICではないかというのがこの論文の主張で、その次の節にそのことが書いてあるわけです。
# pooneilすばやい反応感謝します。最初はそのつもりではなかったのですが、エントリ書き上げてみるとこれはどうにも呼びつけてしまったとしか思えないですよね。どうもお手数かけてすみません。んでもって、なるほど、このfig.8でのモジュールの分化、というのは面白い話ですね。これが「創発的な問題の分割なのか」というのに対する答えだということですね。よくよく読んでみます。いちばん近い道筋を教えてくださったことに感謝します。あと後半のコメントに関して私が今のところわかったのはsoftmax functionを使うということがprior knowledgeがないときのベイジアン的にやることであって、これを使ってモジュールの選択をするのがMOSAICやmultiple model-based reinforcement learningである、とそういうことであって、この段階ではまだpriorは使わずにボトムアップ的にやっている(ディスカッションの最後の”Combination of this bottom-up mechanism with a top-down mechanism is the subject of our ongoing study.”)、ということですね?とんちんかんなことを言ってるのかもしれませんが、ま、答え合わせみたいなもんということでご勘弁を。
2004年11月06日
■ Science 11/5
に"Thinking About the Conscious Mind." Christof Kochによる"Mind: A Brief Introduction."by John R. Searleのレビュー。
via cogniさんのところの11/5。
Kochが言っているようなneuronal correlates of consciousness (NCC)的なプログラムに対してSearleは否定的だろうから、どんなこと言ってるのか興味があります*1。326ページあるということなので、"The Mystery of Consciousness"とかの一般向けの小さい本とは違った、"The Rediscovery of the Mind "以来のまとまった本、ということになるでしょう。これは読まなくては。
一方で、Christof Kochも新著 "The Quest for Consciousness: A Neurobiological Approach."を出してます(Kochのサイトからいくつかの章がpdfで読めます)。まだごッツいハードカバーなので、ペーパバックになったら読もうと思います。
*1:ちなみに、Searleによる「意識を科学的に研究するには」'98を私的に翻訳したことがあるんだけれど、そのときに作った要約が20000215にあります。
2004年11月05日
■ 順天堂の北澤先生
が岡崎でセミナーと大学院講義を。聞きにいってきました。待ち時間にはいろいろお話もうかがって、面白かったです。なんか自分の話とつなげる糸口を考えてみたり。内容自体は手の交差での時間順序の話(セミナー)と小脳のcomplex spikeの話(大学院講義)がメインでした。
2004年11月02日
■ SFNレポート
ま、いろいろ見てきましたが、印象を:Neural prosthesis増えてます、fMRIが多くなったどころか、もはやsingle-unit studyを演題数で追い越してしまっている感じ、学会が大きくなりすぎでPosterを見る暇がない、後半バテた。以前は最終日が半日だったのが、初日が半日になるようになってました(去年かららしい)。これで最終日が寂れることを抑制できていたので効果的だったと思います。んで、予想するに、これは将来的に初日も全日にする布石だと思いますよ。たいへんだけど、そのほうがまだゆっくり見ていられるかも。
印象に残ったセッションについて、ラボでの報告会での資料作りを兼ねて作ってみましょう。今回、ああ、これはNatureかScience間違いなくいったな、と私が思った演題は二つありました。
- 527.2 "RESPONSE OF NEURONS IN MACAQUE AREA LIP DURING A PROBABILISTIC CLASSIFICATION TASK" Shadlenラボ。タイトル見るとどうってことない感じなのだけれど、これは将来的には"neural correlate of arithmetic operation in macaque area LIP"とかそういう形で出てくることでしょう。簡単に言うと、nonhuman primateに足し算、引き算をさせると、その計算途中と計算結果に対応したニューロン活動がLIPから記録された、というもの。ま、数の表象があるんだから、計算の表象があってもおかしくないけど*1、タスクが凝ってて、いろんなimplicationがあって、すごい。
詳しく言うと、こんな話:被験者は図形を四コ続けて見たあとで、左右のどちらかにサッケードすると報酬がもらえる。図形のレパートリーは12個ぐらいあって、それぞれの図形は左右のサッケードの報酬量の比の情報をもっている。たとえば、正方形が出ると左右の報酬量の比は1:4とか。だから正方形が出たら右にサッケードしたほうが被験者にとってはお得。これだけでもGlimcherがtrialのblock単位で報酬比を変えてたのをtrial単位にできるというだけでかなりナイスアイデアかつ、トレーニングが大変そうなのだけれど、1trialでこういう図形が連続して四種類出る。被験者は四つの図形それぞれで決められている報酬比の情報のlog likelihoodの和を計算してやっと左右どちらにサッケードしたほうがお得か決めることができる。たとえば、図形1が1:4、図形2が8:1、図形3が2:1、図形4が1:16だったとすると、それぞれのlog likelihood(log(左/右))はlog(1/4)、log(8/1)、log(2/1)、log(1/16)で、合計するとlog(1/4)なので、右にサッケードしたほうが1:4で得となる、というわけです。で、実際行動はそうなるし、個々の図形のlikelihood ratioを把握していることを示す行動データもあります。さらに、LIPから記録すると、四個の図形が順番に呈示されるたびに、evidenceが蓄積していって、ニューロンの発火頻度が図形ごとに上下して、最終的にどちらにサッケードするかの最終決定を表す発火頻度になってサッケードする、というわけです。つまり、4つの図形が呈示されているあいだに関しては、完全にdecisionシグナルの蓄積過程(計算の途中経過)を表象していると言えて、その途中過程で左右どちらに行くかのdecisionの結果(右か左か)を二値的に表象しているわけではないことが非常に明確に示されているわけです。*2
このタスクの徹底的さ加減もすばらしい。二つぐらい足せば充分じゃんとか考えてると足をすくわれるわけで、こういう徹底的さには心当たりがある。
んでもってしかもいちばん最初に書いたように、これは足し算引き算のneural correlateであるとも言えるわけです。タイトルにもポスターにもそういうことはまったく謳っていなかったけれども(たぶんそう主張するには私が気付かないような障壁がいろいろあるのでしょう)、演者に聞いたらそうだと言ってました。その主張が文句なく通ればNature、文句ありでもScienceは間違いなく通ると見ましたが。
- 646.2 "TWO-PHOTON CALCIUM IMAGING OF VISUAL CORTEX: ORIENTATION MAPS WITH SINGLE-CELL RESOLUTION" Clay Reidラボ。id:kkitaさんも11/1に書いてますが、これは間違いなくすごい。おめでとうございます。つづきは明日書きます、迷惑にならないように気を付けますので。
*1:しかしposterior parietal cortexは時間もあれば数もあれば計算もあればマッチングの法則もあればナッシュ均衡もあればmental rotationもあるわけで、ここは量:quantityの領野と名付けよう。ちなみにventral pathwayの連合野は質:qualityなわけ。
*2:トレーニングの過程では多分じょじょに図形の数を増やしていったのだから、行動のselectionの結果を表象しててもおかしくはなさそうなのだけれど、そういうのはFEFでやっているのでしょう。
2004年10月13日
■ オブジェクト指向プログラミング
がMATLABではどのくらいできるか。
MATLABのhelpdeskを見ると「クラスとオブジェクト」という章があって、なにやらできるらしい。勉強しなくては。
2004年10月12日
■ PLoS biology
のフォーラムページ。
"Breaking Down the Stereotypes of Science by Recruiting Young Scientists ."
科学者、といえば白衣を着た髪ボサボサの老人(=マッドサイエンティスト)というステロタイプがあります。これを崩して、若い人たちにもっとリアルな科学者の姿を見てもらい、彼らをinvolveするにはどうしたらいいか、Thomas Jefferson University Science Outreach Programでの実践の報告。
Referenceで"An Emerging and Critical Problem of the Science and Engineering Labor Force."(The National Science Foundationの記事)へのリンクあり。
私自身は幸運なことに、実験というものにそんなに抵抗感を植え付けられずに済んできたという印象があります。小学校のときには区でやっていた科学プログラムに参加して寒天培地にカビを植えつけて培養した経験がありました(ベランダに置いておいたら、ノラ猫に食われてしまったのだけど)。中高では学校が実験に力を入れてくれていたこともあって、かなり手を動かす機会があったと思います。(高校の化学では、未知のイオン溶液をグループごとに与えられて、化学反応をもとにそれがなんであるかを当てさせる、という実習がありました。あれは大学の教養課程でやるようなレベルの実習ではなかったろうか。)それは恵まれていたことだったんだな。まあその一方で、世の中を金や物がどう回ることでこの世界が動いているのかについて充分リアルな形で学ぶ機会がなかったようにも思うのだけれど。
2004年10月05日
■ Richard Axel
がノーベル生理学医学賞。
nobelprize.orgのプレスリリース。
PubMedだとこんな感じ。
http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=PureSearch&db=pubmed&details_term=axel%20r%5Bau%5D
レビューを除くと、この10年間Cell,Nature,Science,Neuron,PNASにしか書いていない。恐るべし。追記:Nature Neuroscienceがないことに気付く。NeuronとNature Neuroscienceの選択だったら迷わずNeuron、ということなんだろうなあ。Cell pressだし。
ところで研究内容についてまったく触れてない私。
2004年09月21日
■ Back-to-back
- "Neural activity predicts individual differences in visual working memory capacity."
- "Capacity limit of visual short-term memory in human posterior parietal cortex."
*1:こんなこと調べちゃう俺に乾杯。
2004年09月14日
■ Neural correlates of time perception
古い順に。
- NeuroImage '01 "Cortical Networks Recruited for Time Perception: A Monkey Positron Emission Tomography (PET) Study." 尾上先生@神経研
- Neuron '03 "Representation of Time by Neurons in the Posterior Parietal Cortex of the Macaque." Matthew I. Leon and Michael N. Shadlen。
- European Journal of Neuroscience '04 "Stimulus duration in working memory is represented by neuronal activity in the monkey prefrontal cortex." 櫻井芳雄先生@京大
- Annual Review of Neuroscience '04 "THE NEURAL BASIS OF TEMPORAL PROCESSING." Michael D. Mauk and Dean V. Buonomano
- Current Opinion in Neurobiology '04 "The neural representation of time." Richard B Ivry and Rebecca MC Spencer
- JNS '04 "Climbing Neuronal Activity as an Event-Based Cortical Representation of Time." Yakovlev
2004年08月27日
■ サピア・ウォーフの仮説
に関する論文がScienceのAOPに出ています。
"Numerical Cognition Without Words: Evidence from Amazonia."
これの解説は今週のScienceで出てます。
"Life Without Numbers in the Amazon."
関連記事:
- "EP : end-point 科学に佇む心と体: サピア=ウォーフ仮説再考"
- "Study Of Obscure Amazon Tribe Sheds New Light On How Language Affects Perception."
id:cogniさん、すばらしい解説をありがとうございます。積んでた「言語を生みだす本能」を開いてみました。第3章「思考の言葉--心的言語」のところですね(3章の途中まで書き込みがあるので、どうやらここで挫折したらしい)。ぜひ参考にさせていただきます。強い主張と弱い主張、これもカテゴリカルエラーの問題ですね。
「言語がいかに強大な影響力を持つといっても、網膜にまで手を伸ばして、神経節細胞をつなぎなおすなどと主張したら、生理学者は目をむくことだろう。」(言語を生みだす本能 (上) p.83)
言語によって神経節細胞レベルでのwiringのつなぎ変えがおこることを主張されたら、たしかに生理学者としてそれはかなり疑わしいと思います。しかし、神経節細胞のつなぎ変えはred-greenやblue-yellowの軸を反転させるようなdrasticな変換であって、ここで問題になっているような色のカテゴリーがどう変わるか、という問題ではありません。また、色の知覚やカテゴリー化が経験(言語ではないけれど)によって変わるという報告について以前(8/5)わたしのところで採り上げたことがありますけど、このときもそこでつなぎ変えられているのは神経節細胞とかではなくて大脳皮質であると考えられています。だからPinkerのここでの論法は、議論されているものよりは極端な命題を持ってきてそれをrejectしているということのように見えます。
2004年08月25日
■ PNAS
ついでにPNASのtrack I-IIIについて説明しておきましょう。PNASというのは全米科学アカデミーの雑誌ですので、全ての論文は必ずアカデミーのメンバーの誰かによってハンドリングされます。
- Track III: タイトルの下に"Contributed by" と書かれているやつです。アカデミーのメンバーが自分自身の論文を寄稿するもの。以前言及したFrancis Crickのサーチライト仮説はこの範疇ですな。
- Track I: タイトルの下に"Communicated by" と書かれているやつです。アカデミーのメンバーでない者の論文をアカデミーのメンバーが受け取って、refereeに回したうえで、PNASのofficeに送るもの。しばしば身内というか弟子筋の論文がこの形で出版されます。
- Track II: タイトルの下に"Edited by" と書かれているやつです。上記の二つはいかにも内輪の占有になってて評判悪かったのでしょう。1996年からこのtrackができました。論文をPNAS officeに直接送って、アカデミーメンバーの誰がハンドルするのが良いかをカバーレターに書いておくと、editorial boardがハンドルするアカデミーメンバーを決めます。そのアカデミーメンバーがrefereeを決めてreviewプロセスを進めるというものです。
ちなみにSystems Neuroscienceのアカデミーのメンバーのリスト。
久しぶりにサイトのホームに行ってみたら、Classic PNAS articleなんてのができてます。John NashのNash平衡に関する論文はPNASだったんだ。"Equilibrium points in n-person games."が落とせます。たった2ページ!というか正味1ページですよ。"John F. Nash, Jr. - Prize Seminar" ここを見るかぎり、この論文がノーベル経済学賞受賞となった仕事の最初の論文らしい。
ところでsystem neuroscienceでPNASに載ったものでclassical paperになっていると言えるものはあるでしょうか。Hopfieldの論文なんかは分野がちょっと違うしなあ。Cytochrome oxidase stainingは最初はPNASだったっけ? Optical imagingとかでも初期の重要な論文はPNASだったかもしれません。
PNASに掲載されたsystem neuroscience系のclassical paperのリスト。ほかにご存知の方はお知らせください。
Cytochrome oxidase blob
- '82 "Thalamic inputs to cytochrome oxidase-rich regions in monkey visual cortex." Livingstone and Hubel。Fig.3見てるとV1とV2ってこんなにも構造が違うのかと印象深いです。
Grinvaldはたしかに90年代初期にPNASにいろいろ出していましたが、最初のintrinsic signalのoptical imaging paper はNature '86だったようで(Blasdelによるvoltage-sensitive dyeのNature '86の半年遅れ)、わたしの勘違いでした。
# ご隠居
小川誠二先生のBOLDの一連の論文もPNASですね.1990年と1992年ですが,もうclassical paperっていってもよいでしょう.
# pooneilすばやいっすね。なるほど、なんかimagingに強い印象がありますね。あんまりsingle-unitでがちがちの電気生理ってのは見かけないし。リストに追加しておきました。
2004年08月20日
■ Current biology論文関連
mdsさんからの書き込み(8/5)への返答をこちらにも書いておきます。
やはり単純な色の感覚と電磁波の波長とは一対一対応するものであり、恒常性のような「高次過程」のみが発達によって可塑的に変化するものなのでしょうか?
このへんが一番面白いところなのではないかと思います。線分なら単にコントラストがぼやけて見えるようなことが容易に想像できるわけですが、色の経験が変容されるとしたらいったい何が起こるんでしょう。色空間の位相的関係は保持されたままそれが縮んだりするんでしょうか。色と電磁波の波長とは一対一対応はしないのではないでしょうか。黄色の単波長光だろうと緑と赤の混色だろうと同じ黄色として感じますよね。そのような混色の関係が変わって色空間が歪むであろうことは予想できるわけですが。
ところで「先天盲開眼者の視覚世界」、ちょうど別件で読んでいたところです。重要ですよね。しかしこれ読みすすめててもやっぱりどう感じているのかはよくわからないんです。色の識別はだんだんできるようになるし、色名もどんどん付加されていって分化しているわけだけど、その分化する前に見えていた色(たとえば、名づけることができなかった緑)は、分化した後に呈示された同じ色と同じように見えていたのか。クオリア自体が変わったのか(暗くてよくわからないものからビビッドな緑へ)、それともその緑を緑たらしめる赤や緑とも相対的関係が変わっただけなのか(緑-赤の軸ができるまでは緑は青-黄または白-黒の軸からは分離されえない)。うーむ、もっと読み進めてみます。
2004年08月13日
■ しゃっくり(hiccup)
欠伸のつぎはしゃっくりということで。
- Clinical Neuropharmacology '01 "The Hiccup Reflex Arc and Persistent Hiccups with High-dose Anabolic Steroids: Is the Brainstem the Steroid-Responsive Locus?"
- Clinical Neurology and Neurosurgery '00 "Persistent hiccup as presenting symptom in medulla oblongata cavernoma: a case report and review of the literature."
- BMJ '99 "Intractable hiccups."
2004年08月12日
■ Faculty of 1000
Faculty of 1000すげ―気になる。でもうちのinstituteはsubscribeしていない。個人で一週間のtrialが可能なのだけれど、ちょっといまはタイミングが悪いのでまだ試してません。
すでにestablishされた人たち(Neuroscienceのfaculty membersはこちら)を数多く集めて、journalによらずによい論文を評価、発掘していくという趣旨は、(エラそーな言い方だが)言ってみれば私がこのサイトでやろうとしていることをものすごく強力にしたようなものであるわけだし。こうやって自力で論文をチェックしていると、どうしても採り上げる雑誌が有力雑誌に片寄ってしまう(EBR、EJN、neuroreport、neuroscience lettersまでチェックが行き届かない)という問題意識もあります(もちろん、私の仕事に直結する論文はどんなjournalに載っていようと洩らさず読むわけですが)。そういう意味で"Hidden Jewels"ってかなりすばらしいことだと思います。雑誌単位でのIFから論文ごとのF1000 factorへ、という流れができたとしたらその方が健全だと思いますし。
とりあえず誰がどんな論文を評価しているかだけでも読んでいると面白い。GoodaleがMilnerの論文を評価するってそれはないんじゃないの、とか。どうせEarl Millerは師匠のDesimoneの論文とか入れてるんでしょ、とか思ってEarl MillerのEvaluationsを見てみたら…いい奴じゃん<-都合良すぎ。
Subscribeできるようになったら、faculty memberのコメントにさらに突っ込みを入れる、なんてのもいいかも。
PubMedからもリンクされるようになったので、かなーり必須なサイトだと思うんだけれど、そういうわけでまだこれ以上コメントできません。
■ Brain Waves: What is Neurofinance?
Brain Waves: What is Neurofinance?
Neuroeconomicsのお次はneurofinanceだそうな。うーむ、うさんくさい。
2004年08月11日
■ 80年代バージョン
id:cogniさんのところを見て、8/2のやつの80年代バージョンを作ってみました。AIとコネクショニズムの時代であったことがわかります、というかそういう印象になるように選んでみました。。
- Hubert L Dreyfus "What computers can't do: The limits of artificial intelligence" '72
- Douglas R. Hofstadter "Godel, Escher, Bach: An Eternal Golden Braid" '79
- James J. Gibson "The Ecological Approach to Visual Perception" '79
- Douglas R. Hofstadter (Editor), Daniel C. Dennett (Editor) "The Mind's I: Fantasies and Reflections on Self & Soul" '81
- David Marr "Vision: A computational investigation into the human representation and processing of visual information" '82
- Jerry A. Fodor "The Modularity of Mind" '83
- John Searle "Minds, Brains and Science (1984 Reith Lectures)" '84
- Terry Winograd "Understanding Computers and Cognition : A New Foundation for Design" '86
- Rumelhart, McClelland, & the PDP research group (Eds.) "Parallel Distributed Processing: Explorations in the Microstructure of Cognition : Foundations" '86
- Marvin Minsky "SOCIETY OF MIND" '87
- Daniel C. Dennett "The Intentional Stance" '87
- Roger Penrose "The Emperor's New Mind: Concerning Computers, Minds, and the Laws of Physics" '89
2004年08月07日
■ Francis Crick 訃報つづき
Nature 8/5。
"From DNA to consciousness - Crick's legacy."
および"Nature web focus: Francis Crick"
しまった、Crickが脳科学で最初にやったことといえば、「サーチライト仮説」ではなくて、「REM睡眠で夢を見ている間に記憶が強化または消去される」というNature '83 "The function of dream sleep."(pdfファイル。フリーで落とせます)の方でした。この説はのちにMatthew A. Wilson(当時McNaughton研@University of Arizona、現在MIT)によってScience '94 "Reactivation of hippocampal ensemble memories during sleep"という形で結実します。
ところで、"From DNA to consciousness - Crick's legacy."
の最後の方で気になることが書いてあります。
"He thought the claustrum would be a good test system for the centre*," Murphy says. The centre will focus on the genes, proteins and neural networks that make the brain function. (*: 今年Salk instituteにできたCrick-Jacobs Centerのこと。)
The claustrum (前障)? Claustrumなんて考えたことなかった。Insulaとputamenに挟まれた薄いcortexで、いまだにその機能がぜんぜんわかってない脳領域です。Human fMRIとかだとinsulaのactivationと混ざってしまう模様。そういう場所なので、anatomyぐらいしか充分行われてません。
Subcortical structureという言い方がされますが、Trends in Neurosciences '98 "What is the amygdala?"をみるかぎりだと、発生的にはcortexで、amygdalaとかとも近いようです。
Connectivityに関してはmacaqueで最近の論文としては
- The Journal of Comparative Neurology '02 "Projections of the claustrum to the primary motor, premotor, and prefrontal cortices in the macaque monkey."
- Experimental Brain Research '97 "Projections from the claustrum to the prelunate gyrus in the monkey."
機能に関しては2-DGでcross-modal (visual/tactile) recognitionでの活性が認められること、丹治研の島さんによるsingle-unit study(JNP '96 "Neuronal activity in the claustrum of the monkey during performance of multiple movements.")でmovement-related acitivityが見つかること、ぐらいしかないようです(すくなくともnon-human primateでは)。
2004年08月05日
■ Francis Crick関連
Science And Consciousness Review
Nature Neuroscience '03 "A Framework for Consciousness"に関する議論がここから辿れます。
2004年08月04日
■ Figureのplot
ってみなさんどうやって作ってます? 私はmatlabの図をprint -depsc2でepsファイルにしてからillustratorで開くか、Excelで作った図をillustratorにコピペするかしてました。どちらも一回一回手作業なのでバカらしいのですが、けっきょく図をfinalizeする過程でplotの縦横比が変わったり、拡大縮小する際に線が細くなったりしてしまうので、illustrator上で作っておかないときれいにならないという印象がありました。Illustratorのなかでデータを持っておいて図を作る機能もあるのだけれどどうにもしょぼくて使うのを止めてしまった。本当はグラフ作成ソフトを使いこなす方がよいのだろうけど。
2004年07月23日
■ clustering
そうか、linear discriminant analysisとmultidimensional scalingとはsupervisedであるか否かの点で別物か。そりゃそうだった(情報提供from ガヤ)。というわけでメモメモ。
2004年06月21日
■ 「NatureとScienceに載った論文リスト」マイナーアップデート
せっかくPlatt and Glimcher論文のリンクを作ったので、6/17のテーブルにもタイトルとリンクをつけてみました。これからこの表にある論文をリンクすることがあったらそのついででアップデートしていくことにします。一挙にやるのは厳しいし。あ、テーブルから論文の説明へも飛べるようにしておくといいかも。
2004年06月18日
■ NatureとScience

つづき。掲載論文数の推移のグラフを載せときます。Natureが2003年からガタっと落ちていて、Scienceは逆にそのへんでいちどピークを迎えたことがわかります。
2004年06月17日
■ NatureとScience
私の分野でNatureとScienceに載った論文リストというのを以前作ったことがあるのですが、これをアップデートして載せておきます。Nature/Science主義の弊害みたいなのを踏まえつつ、ご覧いただければ。
1999年から現在までです。タイトルはなくしてしまいました。今のところ著者から推測するしかありません。単純な検索で作ったので漏れがあるかもしれません。私の分野でほかにご存知ならお知らせください。日本人による日本での仕事には水色を、日本人による海外での仕事には緑色を塗ってあります。
分野としてNatureに載ることがものすごく減り、2003年以降、Natureは日本人の論文をまったく採らなくなったことがわかります。(1988年あたりからのNatureについても作ったことがあるのですが、そのときのうろ覚えでは、そもそも以前は私の分野の論文は年に数本しか掲載されていなかったし、ここ近年はがものすごく増えてきて、それをNature Neuroscienceが発刊(1998年5月)されることで吸収されて、現在だんだん停滞している、というのが主な流れであるといえます。)
この表は私の分野においてはScienceよりNatureのほうが片寄っているというかへんなセレクションであることを示す証拠のひとつです。これはNatureはもはや行動中のsingle-unitを記録してneural correlateを見つけるというような主流でありつづけた研究はとらなくなった、ということでもあります(そこでneural prosthesisがもてはやされて軍の金が投入されることになったりする)が、その代わりに光るものを持ってきているかというとそれは疑問です。また、このことはNatureの編集者が怠慢か力不足で編集会議で他の分野を担当する編集者とのあいだでの戦いに負けつづけていることを示しているとも言えるかもしれません。すくなくともこの一、二年はScienceの方がよっぽど面白い論文を出していると私は思ってるのですが、いかがでしょうか?
| Journal | Year | Month | Day | Vol. | Page | Authors |
| Science | 2004 | May | 7 | 304 | 878-880 | Clarke HF, Dalley JW, Crofts HS, Robbins TW, Roberts AC |
| Science | 2004 | Apr | 9 | 304 | 307-310 | Roesch MR, Olson CR |
| Science | 2004 | Feb | 6 | 303 | 853-856 | Wang M, Vijayraghavan S, Goldman-Rakic PS |
| Nature | 2004 | Jan | 29 | 427 | 448 - 451 | Poremba A, Malloy M, Saunders RC, Carson RE, Herscovitch P, Mishkin M |
| Science | 2004 | Jan | 16 | 303 | 380-383 | Schwartz AB, Moran DW, Reina GA |
| Science | 2004 | Jan | 16 | 303 | 377-380 | Fitch WT, Hauser MD |
| Nature | 2003 | Dec | 11 | 426 | 668 - 671 | Chatterjee S, Callaway EM |
| Science | 2003 | Oct | 31 | 302 | 881-885 | Chen LM, Friedman RM, Roe AW |
| Science | 2003 | Oct | 3 | 302 | 120-122 | Ito S, Stuphorn V, Brown JW, Schall JD |
| Science | 2003 | Aug | 29 | 301 | 1246-1249 | Fujii N, Graybiel AM |
| Nature | 2003 | Aug | 7 | 424 | 674 - 677 | Krekelberg B, Dannenberg S, Hoffmann K-P, Bremmer F, Ross J |
| Science | 2003 | Jul | 11 | 301 | 233-6 | Ohbayashi M, Ohki K, Miyashita Y. |
| Science | 2003 | Jul | 11 | 301 | 229-32 | Matsumoto K, Suzuki W, Tanaka K. |
| Science | 2003 | Jun | 13 | 300 | 1758-63 | Sharma J, Dragoi V, Tenenbaum JB, Miller EK, Sur M. |
| Science | 2003 | Jun | 6 | 300 | 1578-81 | Wirth S, Yanike M, Frank LM, Smith AC, Brown EN, Suzuki WA. |
| Nature | 2003 | Jun | 26 | 423 | 937-8 | ASIF A. GHAZANFAR AND NIKOS K. LOGOTHETIS |
| Science | 2003 | May | 2 | 300 | 812-5 | Leventhal AG, Wang Y, Pu M, Zhou Y, Ma Y. |
| Science | 2003 | Mar | 21 | 299 | 1898-902 | Fiorillo CD, Tobler PN, Schultz W. |
| Science | 2003 | Feb | 14 | 299 | 1073-5 | Barbour DL, Wang X. |
| Science | 2003 | Jan | 24 | 299 | 568-72 | Poremba A, Saunders RC, Crane AM, Cook M, Sokoloff L, Mishkin M. |
| Science | 2003 | Jan | 3 | 299 | 81-6. | Bisley JW, Goldberg ME. |
| Nature | 2003 | Jan | 30 | 421 | 535-9 | Xiao Y, Wang Y, Felleman DJ. |
| Nature | 2003 | Jan | 23 | 421 | 370-3 | Moore T, Armstrong KM. |
| Nature | 2003 | Jan | 23 | 421 | 366-70 | Thiele A, Stoner G. |
| Science | 2002 | Oct | 18 | 298 | 572-6 | Adams DL, Horton JC. |
| Science | 2002 | Oct | 11 | 298 | 413-5 | Vanduffel W, Fize D, Peuskens H, Denys K, Sunaert S, Todd JT, Orban GA. |
| Science | 2002 | Oct | 11 | 298 | 409-12 | Tsutsui K, Sakata H, Naganuma T, Taira M. |
| Nature | 2002 | Oct | 10 | 419 | 616-20 | Ghose GM, Maunsell JH. |
| Science | 2002 | Sep | 27 | 297 | Ricaurte GA, Yuan J, Hatzidimitriou G, Cord BJ, McCann UD. | |
| Science | 2002 | Sep | 20 | 297 | Hoffman KL, McNaughton BL. | |
| Science | 2002 | Sep | 6 | 297 | 1708-11 | Nieder A, Freedman DJ, Miller EK. |
| Nature | 2002 | Sep | 12 | 419 | 157-62 | Fukushima K, Yamanobe T, Shinmei Y, Fukushima J, Kurkin S, Peterson BW. |
| Science | 2002 | Aug | 2 | 297 | 846-8 | Kohler E, Keysers C, Umilta MA, Fogassi L, Gallese V, Rizzolatti G. |
| Nature | 2002 | Aug | 8 | 418 | 633-6 | Cumming BG. |
| Nature | 2002 | Jul | 25 | 418 | 413-7 | Lauwereyns J, Watanabe K, Coe B, Hikosaka O. |
| Science | 2002 | Jun | 7 | 296 | 1829-32 | Taylor DM, Tillery SI, Schwartz AB. |
| Nature | 2002 | Jun | 27 | 417 | 938-41 | Gribble PL, Scott SH. |
| Science | 2002 | May | 31 | 296 | 1709-11 | Shidara M, Richmond BJ. |
| Science | 2002 | May | 24 | 296 | 1480-2 | Sommer MA, Wurtz RH. |
| Nature | 2002 | Apr | 11 | 416 | 632-6 | Buneo CA, Jarvis MR, Batista AP, Andersen RA. |
| Science | 2002 | Mar | 29 | 295 | Froehler MT, Duffy CJ. | |
| Science | 2002 | Mar | 29 | 295 | Thiele A, Henning P, Kubischik M, Hoffmann KP. | |
| Nature | 2002 | Mar | 14 | 416 | 141-2 | Serruya MD, Hatsopoulos NG, Paninski L, Fellows MR, Donoghue JP. |
| Science | 2002 | Feb | 22 | 295 | 1532-6 | Nakahara K, Hayashi T, Konishi S, Miyashita Y. |
| Science | 2002 | Feb | 15 | 295 | 1314-6 | Klier EM, Wang H, Constantin AG, Crawford JD. |
| Science | 2002 | Feb | 1 | 295 | 862-5 | Seidemann E, Arieli A, Grinvald A, Slovin H. |
| Nature | 2002 | Feb | 21 | 415 | 918-22 | Sawamura H, Shima K, Tanji J. |
| Nature | 2002 | Jan | 10 | 415 | 165-8 | Toth LJ, Assad JA. |
| Nature | 2002 | Jan | 17 | 415 | 318-20 | Sigala N, Logothetis NK. |
| Science | 2001 | Dec | 21 | 294 | Reich DS, Mechler F, Victor JD. | |
| Science | 2001 | Dec | 7 | 294 | 2127-30 | Kornack DR, Rakic P. |
| Nature | 2001 | Dec | 20-27 | 414 | 905-8 | Pack CC, Berezovskii VK, Born RT. "Dynamic properties of neurons in cortical area MT in alert and anaesthetized macaque monkeys." |
| Nature | 2001 | Sep | 13 | 413 | 161-5 | Scott SH, Gribble PL, Graham KM, Cabel DW. |
| Nature | 2001 | Aug | 2 | 412 | 549-53 | Schoups A, Vogels R, Qian N, Orban G. |
| Science | 2001 | Jul | 6 | 293 | 120-4 | Super H, Spekreijse H, Lamme VA. |
| Nature | 2001 | Jul | 5 | 412 | Waelti P, Dickinson A, Schultz W. | |
| Nature | 2001 | Jul | 12 | 412 | 150-7 | Logothetis NK, Pauls J, Augath M, Trinath T, Oeltermann A. |
| Nature | 2001 | Jun | 21 | 411 | 953-6 | Wallis JD, Anderson KC, Miller EK. |
| Science | 2001 | Apr | 13 | 292 | 297-300 | Yabuta NH, Sawatari A, Callaway EM. |
| Science | 2001 | Apr | 13 | 292 | 290-3 | Tian B, Reser D, Durham A, Kustov A, Rauschecker JP. |
| Nature | 2001 | Apr | 19 | 410 | 933-6 | Martin PR, Lee BB, White AJ, Solomon SG, Ruttiger L. |
| Science | 2001 | Feb | 23 | 291 | 1560-3 | Fries P, Reynolds JH, Rorie AE, Desimone R. |
| Nature | 2001 | Feb | 22 | 409 | 1040-2 | Pack CC, Born RT. "Temporal dynamics of a neural solution to the aperture problem in visual area MT of macaque brain." |
| Science | 2001 | Jan | 12 | 291 | 312-6 | Freedman DJ, Riesenhuber M, Poggio T, Miller EK. |
| Science | 2001 | Jan | 26 | 291 | 661-4 | Naya Y, Yoshida M, Miyashita Y. |
| Nature | 2001 | Jan | 11 | 409 | 191-4 | Tanaka M, Lisberger SG. |
| Science | 2000 | Dec | 1 | 290 | 1786-9 | Hasegawa RP, Blitz AM, Geller NL, Goldberg ME. "Neurons in Monkey Prefrontal Cortex That Track Past or Predict Future Performance" |
| Science | 2000 | Dec | 1 | 290 | 1782-6 | Graziano MS, Cooke DF, Taylor CS. |
| Nature | 2000 | Dec | 21-28 | 408 | 971-5 | Zhang M, Barash S. |
| Nature | 2000 | Dec | 14 | 408 | 857-60 | Stuphorn V, Taylor TL, Schall JD. |
| Nature | 2000 | Nov | 23 | 408 | 466-70 | Hoshi E, Tanji J. |
| Nature | 2000 | Nov | 16 | 408 | 361-5 | Srinivasan MA, Nicolelis MA. |
| Nature | 2000 | Oct | 26 | 407 | 1003-7 | Gamlin PD, Yoon K. |
| Science | 2000 | Jun | 16 | 288 | Janssen P, Vogels R, Orban GA. | |
| Nature | 2000 | May | 18 | 405 | 347-51 | Fuster JM, Bodner M, Kroger JK. |
| Nature | 2000 | May | 4 | 405 | Thier P, Dicke PW, Haas R, Barash S. | |
| Science | 2000 | Mar | 17 | 287 | Castner SA, Williams GV, Goldman-Rakic PS. | |
| Nature | 2000 | Mar | 9 | 404 | 187-90 | Steinmetz PN, Roy A, Fitzgerald PJ, Hsiao SS, Johnson KO, Niebur E. |
| Nature | 2000 | Mar | 2 | 404 | 77-80 | Orlov T, Yakovlev V, Hochstein S, Zohary E. |
| Science | 2000 | Feb | 25 | 287 | 1506-8 | Rollenhagen JE, Olson CR. |
| Science | 2000 | Feb | 18 | 287 | 1273-6 | Vinje WE, Gallant JL. |
| Science | 1999 | Oct | 15 | 286 | 548-52 | Gould E, Reeves AJ, Graziano MS, Gross CG. |
| Nature | 1999 | Oct | 14 | 401 | 699-703 | Tomita H, Ohbayashi M, Nakahara K, Hasegawa I, Miyashita Y. |
| Nature | 1999 | Oct | 7 | 401 | 590-4 | Prut Y, Fetz EE. |
| Science | 1999 | Sep | 24 | 285 | Kakei S, Hoffman DS, Strick PL. | |
| Science | 1999 | Sep | 17 | 285 | Moore T. | |
| Nature | 1999 | Sep | 16 | 401 | 269-72 | Sugita Y. |
| Science | 1999 | Aug | 27 | 285 | 1405-8 | Muller JR, Metha AB, Krauskopf J, Lennie P. |
| Nature | 1999 | Aug | 26 | 400 | 869-73 | Sugase Y, Yamane S, Ueno S, Kawano K. |
| Nature | 1999 | Jul | 15 | 400 | 233-8 | Platt ML, Glimcher PW. "Neural correlates of decision variables in parietal cortex" |
| Nature | 1999 | Jun | 10 | 399 | 575-9 | Treue S, Martinez Trujillo JC. |
| Nature | 1999 | Jun | 3 | 399 | 470-3 | Romo R, Brody CD, Hernandez A, Lemus L. |
| Science | 1999 | May | 14 | 284 | 1158-61 | Horwitz GD, Newsome WT. |
| Nature | 1999 | Apr | 22 | 398 | 704-8 | Tremblay L, Schultz W. |
| Science | 1999 | Mar | 12 | 283 | 1752-7 | Carpenter AF, Georgopoulos AP, Pellizzer G. |
| Nature | 1999 | Mar | 18 | 398 | 239-42 | Trotter Y, Celebrini S. |
| Nature | 1999 | Feb | 4 | 397 | 428-30 | Graziano MS, Reiss LA, Gross CG. |
| Science | 1999 | Jan | 22 | 283 | 549-54 | Usher M, Cohen JD, Servan-Schreiber D, Rajkowski J, Aston-Jones G. |
いろいろツッコミどころはあります。Earl Miller落ちぶれたなあとかNewsome意外に苦戦しているとか。Pack and Bornが一年に二度Natureに載る価値があったろうか、とか。最近話題にしていた人がたくさん載っていて、けっこうすでにいろいろ言及していること(ScottやらMooreやら)。Peter Thier小脳もやってたのか!とか。
ところでこの表の全部にタイトルとfull textへのリンクを付けて各論文の簡単な説明をつけたらかなり有用になることに気付いてしまった。
# mmmm
Newsome、満を持して出てきましたね。
# pooneil書いてたらいきなり来ましたね。>Newsome。しかもなんかはやりのゲーム理論っぽいやつ。ぜひ、読んでみたいと思います。とはいえ、この間のNature Neuroscienceのやつとか、急速に難解化しているんで読めるかどうか自信はありません。
# pooneilこの論文で使っているのはmatching theoryであってゲーム理論ではありませんでした。訂正します。数理的に難しくはなさそうだけど、conceptualにけっこう難しそうです。
2004年06月04日
■ 英語の脳科学関係のblog
を以前探したことがあるのだけれど、あまりありませんでした。とりあえず
私がやっているような論文チェック日記サイトを探しているのだが、そういうのは英語のblogにはないのでしょうか、ということでご存知の方、報告期待します。
もちろんこの問題は、日本語のわからない英米人には読まれないことを見越して書いているという私自身の日記の事情とカップルしていることはわかっているのですが。そしてこのことが、日本人の論文についてはコメントしにくくて、英米人の論文にはコメントしやすいということの理由であって、ある種、高みの見物を決め込んでしまっている、というこの日記の問題にも繋がります。論文の著者が読んでいることを知っているならばもっと力を入れて書く必要があるし(たとえば、ガヤのScience論文に対して私が行ったように)、論文の著者に読んでもらえるように書きたいのならば、英語でblogを作ってそこでやればよいのです。お、じつはこれはニッチなのだろうか。
脱線してきた!というかこっちが本題なのだけれど。
2004年05月17日
■ Reality Monitoring
NHKさん、ありがとうございます。今回のScale errorの方についてか、子供の意識の発生についてかちょっとわかりませんでしたが、後者の方かなと推測して書きます。
以前NeuroReport '02 "The role of the parahippocampal gyrus in source memory for external and internal events."これが出る前あたりにちょっとreality monitoringを調べた憶えがありますが、改めて見てみると、childhood amnesiaとepisodic memory(またはautobiographical memory)およびreality monitoring(またはsource memory)の関係というあたり面白そうです。"ヒトの意識が生まれるとき"、"赤ちゃんは知っている―認知科学のフロンティア"あたり引っぱり出してみようと思う。
検索かけてみた。
- Developmental Science '04 "Changes in reality monitoring and episodic memory in early childhood." むちゃくちゃ関係ありそうなやつ。子供の発達心理学。Reality monitoringの指標とepisodic memoryの指標とが4歳から6歳あたりの子供で相関していることを示し、さらにこられがprefrontal cortexの発達の指標とも相関していることを示した。
- Schizophrenia Research '03 "Cognitive basis of hallucinations in schizophrenia: role of top-down information processing."
- Behaviour Research and Therapy '03 "Reality monitoring and metacognitive beliefs related to cognitive confidence in obsessive–compulsive disorder."
- Neuropsychologia '03 "Reality monitoring and visual hallucinations in Parkinson’s disease."
- Brain Research Reviews '01 "Spontaneous confabulation, reality monitoring, and the limbic system — a review."
- Journal of Experimental Child Psychology '01 "Reality Monitoring of Performed and Imagined Interactive Events: Developmental and Contextual Effects."
- Psychiatry Research '00 "Positive symptomatology and source-monitoring failure in schizophrenia — an analysis of symptom-specific effects."
- Johnson, M. K. (1988). Reality monitoring: An experimental phenomenological approach. Journal of Experimental Psychology: General, 113, 329-344
- Johnson, M. K., & Raye, C. L. (1981). Reality monitoring. Psychological Review, 88, 67-85
2004年03月21日
■ "The Mind of a Mnemonist" by A.R.Luria
がテレビで扱われていた。こういう番組を見るのは久しぶりだ、っていうかテレビあんまり見ない*1。私はかつて記憶の研究をしていたのだが、恥ずかしながらこの本を読んだことはない。ただ、共感覚について調べているとこの本は必ず出てくる話題で、共感覚についての記述は抜粋を読んだことがある。Webでexcerpt発見。で、この患者さんがどういう人生を経たのかははじめて知った。たしかゲイジも前頭葉を損傷したのちに見世物小屋に出るようなことをしていなかったっけか(たぶんダマジオの本に書いてあったと思う)。
もう一つの前向性健忘も有名な話だが、映像で見せられると迫力が違う*2。あれは無酸素になった状態(一酸化炭素中毒とか)で海馬CA1のpramidal cellが選択的に脱落した例(症例RBが有名)だが、テレビに出た人は家族が支えてくれていて、非常に明るく生活をしているのが印象深かった。
*1:テレビを見るのは子供と一緒のときに限られるので、見てるのはポケモンやハム太郎だったりする。
*2:ワイルダー・ペンフィールドの有名な実験も、写真ではなく映像で見たのははじめてだった。Dura上から刺激しているのだと思うのだけれど、大きくcraniotomyしているのでけっこう拍動していた。
2004年03月09日
■ Daniel Wolpert
Daniel Wolpertのwebサイト(reprintあり)。
Daniel Wolpertこそが内部モデル(のうちの順モデル)がhumanのarm movementの制御に関与していることをはじめて実験的に示した人だ。それ以降も川人先生といっしょに多重順逆モデルを提唱して、その理論を進化させている。
Nature Neuroscience '98 "Central cancellation of self-produced tickle sensation"
自分で自分をくすぐるとくすぐったくないが、人にくすぐられるとくすぐったくない。これに脳のどこが関わっているかをCris FrithといっしょにMRで調べて、小脳が自分をくすぐるときのcaccellationに関わっていることを示している。
Nature '04 "Bayesian integration in sensorimotor learning"
コンピュータカーソルを動かすタスクで、視覚フィードバックに不確定性を加えると、視覚と運動のそれぞれの不確定性をベイズ統計的に扱って学習してゆくことを示した。Prior probability (カーソルがどのくらい横にシフトしているか)と試行ごとの不確定性(視覚フィードバックがどのくらい与えられるか)とからposterior probability(カーソルとターゲットとのずれ)を人間は計算しているらしい。
2004年03月07日
■ 自明性
「ぬ」という字をずっと見つづけていると、この字は"nu"と読む字だったっけ、「め」と「ね」の中間みたいな新字だったっけ、と確信が揺らぐ瞬間がある(いや、あるよね、あるはずだ)*1。このとき脳で自明性を保証している活動に何らかの変化があるのだ。視覚入力はまったく変わっていないのに。よって、Logothetisがbinocular rivalryでやったのをKanwisherがhumanに移植したのと同じことができる。つまり、fMRIでこの二つの条件の差分を取ればよいのだ。もちろん、「ぬ」を見始めと変わってきたところとを比較するのではコントロールの取り方が悪い。そうではなくて、[見つづけて変わってきたtrial vs.変わってこなかったtrial]と[trialの前半vs.後半]とでinteractionがでるところを探してくればよいわけだ。よし、「自明性のneural correlate」のできあがり! だれか撮って。
もちろん被験者はこのような自明性の変化を恣意的にできるようにすることが要求されるし、そのために被験者は自分について現象学的に分析することに慣れておくことが必要となる。これこそがVarelaが言ったneurophenomenologyではないだろうか(冗談のつもりで書いていたがマジになってきた)。
*1:なんにしろ、「ぬ」を"nu"と読むことの自明性さえ壊れてしまえばよい。「ぬ」を"ne"と読める気がしてくるというのでもよいし、解読不能な古代の象形文字に見えてくるというのでもよいし、ジャイアンツの帽子についているあのキャラに見えてくるというのでもよい。
■ Neural correlate
は必ずしも外界の変化に対応した変化ではないかもしれない。もしneural correlateを環境世界とのループの中で捉えるのであるならば、[外界の変化に対して脳がまったく変化しない]ということが何らかの現象のneural correlateであるということがありうる。
# ガヤ
いつも不思議に思うのですがfMRIで引き算すると必須なものまで引いてしまってそこから誤解が生まれることとかってないですか?
あと「ぬ」に関係して、Neural correlateとは関係ないけれど、RumelhartのNecker Cubeモデルを思い出しました。
もう一個。Neural correlateは「無変化」もあり得るという話に賛成。茂木さんは「スパイクこそが情報の全てでそれ以外にはない」としていますが、私は「神経の沈黙(no spike)も情報になる」と考えています。竜安寺の石庭みたいだな(かんけーねーか)。そう考えるとRate vs. Temporal Coding対立も見通しが良くなりますね。
サンクス。Binocular rivalryってのがまさにNecker Cubeみたいなもん。右目と左目に別々の絵が入ってくると、右目の画像しか見えない時間と左目の画像しか見えない時間とが交互に訪れる。これも視覚情報として入ってくるものは同じだけれど、人間の都合で見えが変わる。これを発見。http://www.twcu.ac.jp/~asakawa/chiba2002/Necker/Necker.html 意図してなかったんだけど、昨日出た二つの状態が遷移するという話の続きになったね。MRについてはまた。
ガヤ日記3/5についてだけど、これはおもしろい。2/29に私がコメントアウトしたところに「その直前までの状態の来歴が必要なのか、それともそのシステムは記憶を持たずに済むのか(次の状態を決定する関数には現在の状態だけが必要なのか、それとも以前の状態も必要なのか。)はわからないが。」なんて書いてたもんだから、けっこう驚いた。ある時点iでのネットワークの状態D(i)がそれ以前の状態D(i-1),D(i-2)....のどのくらいに依存しているのか、これはまさにシステムはどのくらい過去(因果性)を持っているのか、という問題への答えだと思う。さらに言えば、それが外部入力(スライスだったら別の領野などへの刺激でもよい)によってドライブされるときにはそれがどう変わるのか、という興味も湧く。アトラクターの谷が入力によって変わり、その上を遷移していたある時点での状態D(i)は自発状態時とは違った方向へ進んでゆく。これがミクロレベルでの遷移ともう少し大きいスケールでの谷の変化とのinteractionという感じか。
経験の一回性または時間性というものは環境世界との関係においてシステムの何かが常にまったく同じではないことからきていると考える。そしてこれをネットワークのシステムの因果性、来歴への依存性から語ってみる。(飛ばしすぎ。)
MRについてはたとえば、あるタスクについて安静時からの差分でまずどこが活動しているかを同定してから、そのROIにおいて活動の大きさが条件によってどう違うか、activationなのか、disinhibitionなのかを議論する、というのがMRの手続きの一つです(ROI analysis)。なんにしろ、「必須なものまで引いてしまって」ということが起こらないように、いかにうまいコントロール条件を作るか、というところがMRでの研究での要点の一つである、というのが私の理解です。
あ、記憶、というのは人間の短期記憶や長期記憶、というつもりで使ったのではなくて、マルコフ過程であるか否か、ということのために出した言葉でした。なんにしろ、ある時間の状態、というやつをどのくらいのビン幅で見るかによって変わる話ではあるわけだけれど。
その意味でしたら答えは私のとった手法は明らかに非マルコフです。重マルコフ過程として捉えるとしたらその次元はおっしゃる通りBinningに依存してきますね。
# pooneilサンクス。重マルコフ過程と言えばよかったわけだ。3/8に続く。
2004年03月06日
■ Stochastic Resonance
は「Noise-induced order」で「もっと高次な非線形ダイミクス」なのかなあ。そのへんがやっぱりわからない。非常に素朴なartifactualな現象のようにも思えるし。で、以前に見つけたのは山本義春さんのページ(ちなみに高校水泳部の先輩)。それからイタリアのページ。
後者のページにあるレビューのfig.1なんかを見てると、たしかにガヤが言うようにBoltzmann machineをイメージするというのは正しいのかもしれない。ガヤがそれをイメージするのはわかる。UP stateとDOWN stateとの間をノイズが付加することによって行ったり来たりしている状態はたしかに二つの安定状態(アトラクタ)を持ったエネルギー曲線と捉えられるのだろうし。
なんにしろたぶん統計物理をわかったうえでないと使えなさそう。U.T.氏がGouldさんのところで書いていたように、素人は深入りは禁物かもしれない。どっかで関係してくる気はしていて、それが私の場合は"detection"という概念への寄与ではないかと考えてはいるのだけれど。で、前者のページを見なおしてみると、Collinsという人の論文*1でnear-thresholdでのdetectionへのnoiseの効果があるとのことだった。Tactileだけど。
追記:義春さんがPhysical Review Lettersに"Behavioral Stochastic Resonance within the Human Brain"というのを出しているのを発見。"The first evidence that stochastic resonance within the human brain can enhance behavioral responses to weak sensory inputs"だそうな。これは3/3に私がイメージしてたのに近い。
さらに調べてみたら、義春さんは上記のJJ Collinsという人といっしょに論文も書いてた。しかもこのCollinsが"Stochastic resonance without tuning"をNatureに出している人だった。つながってきたかも。
*1:Phys. Rev. Eだって。そこまでさすがに手は回らない。
# ガヤ
いつもながら、さすがは鋭い洞察力。このGammaitoniのレヴューはYusteラボで読まされました。しかも、まさにUP-DOWNの絡みです。お見事。一方、山本義春さんの方はむしろD. Fersterのthresholdingの概念に近いですね。
# pooneilサンクス。反応早すぎ。書き込んでから10分しか経ってないよ……おどろいた。
2004年02月29日
■ 昨日の*3の続き。
ここで想定しているのはNature '88 Lehky and Sejnowskiだ。<コメントアウト。ニューラルネットワークの機能の一意性について。>
2004年02月28日
■ ガヤ日記2/25の「分類すること」
ガヤ日記2/25の「分類すること」についてだけど、これは12/19コメント欄および12/20の日記あたりにdecodingの話をしたときと書いたことなのでつなげてみる。パターン識別の本を読んでいると「醜いアヒルの仔定理」というのがある(「認識とパタン」以外にもネットでいろいろ探したことがあるのだが、これのステイタスがどのくらいのものか私は知らない。それは別としていかにも濫用されやすそうな概念だと思う。)。これの帰結は分類には論理的に最適なものはなくて、どのカテゴリー分けに重きをおくか(たとえば、背骨があるかどうかと水中に棲んでいるかどうか。)という評価関数によらなければならない、ということだ。2/23のガヤのコメントでアルゴリズムを探し中と言ってたからきっと「No free lunch theorem」のほうもご存知だと思う。Pragmaticにはどのくらいインパクトがあるのか私にはわからないが。
というわけでこれが関係しているかどうかは別として、前にも言ったように、われわれが外から脳の神経活動をdecodeするときにも何らかの重み付けをすることがたぶん不可欠なんだろう。ただし、このことはdecodeの不可能性を意味しない。<コメントアウト。>ちょっとぱっとはわからないのだけれど、入力と出力が一対一で対応するようなネットワークを作ったときに、中間層への入力の重みだけを見てideal observerがすべての入力情報をdecodeすることは必ず可能なのだろうか?
<コメントアウト。>
2004年02月24日
■ "Neural prosthetics"またはNeuroprosthetic
という言葉自体は前からあるみたい。Pubmed見るかぎりだと。同じ使い方してるかどうかはわからないかど。
■ 「論文など人の仕事を評価する時の姿勢」
「論文など人の仕事を評価する時の姿勢」
なるほど。私は2から3へ行こうとしているつもり。4は悟りの領域だなあ。論文とか読まなくてもよくなりそう。そうだ、レベル5と0に「まったく読まない」っていうのを入れてみるのはどうだろう。まったく同じように見えてぜんぜん違うの。レベル0はたんなる不勉強だが、レベル5はもはや論文を超越してしまって森羅万象を感じるだけで科学になってしまう(電波キター)。なんか修行の道っぽくなってきた。
2004年02月22日
■ ついでに、NatureやScienceに載ること
が科学的にどのくらい重要かという問題にはいろいろ言うべきことはあるが一つだけ。この問題はショートペーパーでどのくらいきっちりした論文が書けるかという問題も含まれる。NatureやScienceはどうしてもインパクトはあるが方法論的詳細が欠けててぜんぜん信用できない、なんてことになりがちだ(私の分野では)。
これに対する過去の人々の対処法は関連するfull paper(私の分野ではJNSやJNP)を書くことだった。前に言及したNewsomeのNature三つはどれも出版後にJNSでほぼおんなじネタを書いている。他にもたくさんある。さすがにここ近年はこういうのは少なくなってきたのだが、それでも最近でも2/19に挙げたSommers and WurtzとかBarashのJNPとかはこのカテゴリーに含まれる。新規性がまったくないんだったら許されるべきではない。データに厚みを加えること自体は科学にとって良いことではあるけれど、その厚みの分がどのくらい新規性をどのくらいあるかという問題。*1
最近はNatureもScienceもsupporting materialを付けることでデータの信頼性を挙げようとしているわけだけれど、それで最近のNature、Scienceは変になってると思うことがある。1/16に挙げたAndrew SchwartzのScienceなんてほとんどのmethodがsupporting materialに回っていて、本文だけではどういうタスクをやってるんだかほとんどわからなくて非常に読みにくい。Schwartzの場合はSOMと別論文*2とを読めばとりあえずわかるようにはなっているのだけれど、他にもいろいろある。(二年前には)Instructionには本文だけで内容が通じるようにすべきと書いてあったはずだけど、と思ってついでに調べてみたらけっこう変わってる!
Natureははじめから別のファイルとしてsubmitするようになってる。Guide to authorには本文にmethodを入れることを指示していて、多かったらpeer-review後に回す、と書いているが。
ScienceがSOAにすべきところを明示して最後尾につけるように、と明示したのはよいことだと思う。
というわけで「本文だけで内容が通じるようにすべき」と書いてあるところはざっと見ただけでは見つからなかった。マジっすか。
なんか脱線してるっぽいが、ショートペーパー集であるということがNatureやScienceが本質的に抱えている問題の一つであって、システムニューサイエンスのように完璧な結論というものがほとんどありえない分野においては詳細が書けないということがかなり足枷になっていると思う。というわけでこの話は「論文誌のブランド志向」という問題とはまったく別の話。
*1:ところでこれは程度を変えればみんな苦労していることだと思う。タスク変えたって、方法論変えたって、前に言ってたことと本質的に変わらんじゃん、という批判から逃れた論文を書くことは本当に難しいことだ。そういう論文こそがNatureやScienceになってほしいと思うし、私もそういうのを書きたい。私は重箱の隅などつつきたくない。いや、究極にはすべて程度の違いの話なんだけど。
*2:これがまたまったく同じタスクでScience論文の方法論的押さえとして働いている。
# pooneil
追記。この問題はまた、システムニューロサイエンスの方法論がどんどん複雑化していることの反映なのだろう。そのこと自体がこの分野が円熟した、または袋小路に入ってきた、ということの徴とも言えるかもしれない。革命的な論文はあっさり短くてreferenceも少ない、というのを夢想するなあ。
# GouldJNPもろくに読めない僕としては、pooneilさんとガヤさんの日記はバイブルのようなものです。できる事なら弟子になりたい・・・
# pooneil2/21日記見ましたけど、NatureでもJNPでも書ききれてないものを読まなければいけないという意味では(程度の差はあれ)同じなんだと思います。その意味では読むのはどれも難しいというわけで。結局前報とかをさらって著者のストーリーとコンテクストを見つけていかないとわからない意味はいろいろあるなあと思います。きりないし全部付き合う必要なんてないけど。(2/23日記へ続く)
2004年02月21日
■ 「ガヤの日記」の2/20
「ガヤの日記」の2/20にうちの2/20のコメント欄についての応答あり。サンクス。そこからわざわざ探して見にきてくれた人がもしいたらサンクス。ちなみに2/20の私のコメントはまるでガヤが[spontaを調べたら情報がわかる]と言っているのに反論しているように見えることに気付いた。べつにそんなことは言ってない。
で、ガヤ日記にあるように、時間的な繰り返しの構造(モチーフ)でspontaが作られているかもしれない、ということは、神経ネットワークのとる構造は実はそんなになくて(自由度は予想されるより少なくて)、そのいくつかの可能な構造のあいだを自発状態で遷移しているということなのかもしれない*1。ガヤがreferしたArieliの論文はそのような遷移しながらできる空間パターンが自発のときと刺激応答のときとで同じであることを示していた……
脳のネットワークの機能構造を調べる研究と入力出力のあいだでの情報変換を調べる研究とが合体するところまではもう近いと思う。たぶん究極は「行動する人間のすべての神経細胞の活動をモニターする」ということだと思うけど、そこまで行かなくてもすべきことはいくらでもある。
ところでそれでも意識と神経活動をつなぐのには問題があるかもしれない、といういわゆる「ハードプロブレム」のことをずっと考えているわけなんだけど(また話をややこしくする)。ここで「オートポイエーシスには入力も出力もない」とはいったいどういうことかを思い出してみる。
*1:これはたぶんCossartのNatureからのimplicationでもあるが「時間的繰り返し」というのがさらに重要なところ。
2004年01月10日
■ 宿題
ラマチャンドランがmirror neuronについて言っていることをちゃんとフォローする。
EDGE 69 — June 1, 2000あたりから。
2004年01月09日
■ Duhamel
が来た。カナダ生まれのフランス在住だけあって、どこかおフランスを感じた。Bullierもそうだったが、なんか微妙にカッコいい。実験セットアップを見てもらいながら一時間ほど話をしたのちに講演を聞いた。講演は基本的にJNS '02のデータの延長上。データ自身は決定的ではなかったと思うが、parietal-prefrontalのスキームは面白かった。とはいえ基本的に彼はGoldberg派であって、attentionへの重みが強いように感じられた。
2004年01月08日
■ 夢
夢がただの幻覚でないのは自分が行動している、という感覚を持つことができているからで、まったくの受身なわけではないから。その意味で、想像(imagery)はトップダウンコントロールによる感覚系の活性化だけで説明がつくが、夢や幻覚にはある種手触り、触れたり移動させたりすることの可能性(=アフォーダンス)がある。つまり夢や幻覚には遠心性コピー(=内部モデル)が戻ってくることが必要。そして覚醒時の意識にはそれ以上のものが必要になる。つまり、遠心性コピーではない、本物のフィードバックがあることがたぶん不可欠なのだ。前に言ってた、想像―夢―幻覚―覚醒時の意識で起こっていることの違いがたぶんここにある。いわば、それぞれのレベルでカップリング(またはバインディング)の仕方が違う、と図式化することができる。
幻覚は覚醒時の意識と共存できるという意味で夢とは違うが、両者の現象学(カッコつけすぎ)は多くを共有している。
自分の行動が環境世界に変化を与える、それを感覚器が受容する、というループ(byユクスキュル)がたぶん現実を構成するのに必要なのであって、これこそがいわゆる身体性であり、「世界は現実にある、という実感」のことなのだ(飛ばしてきた!)。クオリアはこの実感(というのは語弊がある、アプリオリとでも言えばよいか)に大きく依存するが、夢や幻覚にもクオリアがあるように、両者は同一ではない。Sensorimotor coordinationのループと意識とは同一ではないが、このループが意識を可能にしている。
いやいや、夢にもある種の身体性はあるか……止まらなくなってきたのでここまでにしとく。
# がや
私にはわかるようなわからないような微妙な線だけど、上の考え方にはよく共感できる。もし書く時間があったら是非ここで止めないで続けて下さい。
# pooneilサンクス。そのうち続けます。[Sensorimotorループ]と[意識]という別々のオートポイエティックなシステムがあって、その二つの輪が鎖のようにカップリングしている、というのが元イメージなのです。河本英夫氏にはきっぱりと否定されましたが。
# arational agentpooneilさん、新年あけましておめでとうございます。初めてご挨拶申し上げます。私、マインツ大学哲学科にrational agencyを主題とする博士論文を年明けに提出いたしました。その執筆の過程でpooneilさんのサイトを日々読み、おおいに助けられました。大変感謝しております。この夢に関する議論やMilner&Goodaleのtwo visual systemsに関して、pooneilさんのご高論を参考にさせて頂きましたので、お名前(「pooneil」さんではなくご本名)を注で挙げてしまいましたが、ご了承頂けないでしょうか。(提出するまで、論文を提出できるかどうか確信が持てなかったため、ご連絡するかどうか躊躇しておりました。)事後承諾をお願いすることになってしまったこと、ご容赦下さいますようひらにお願い申し上げます。pooneilさんのサイトからの助けがなければ、博論をいまだ完成できなかったと思います。誠にどうもありがとうございました。
# pooneilはじめまして。Rational agencyで哲学科に博士論文で、Milner and Goodaleも関わってくる、とはおもしろそうな話ではないですか。Rational agencyじたいはAIの分野での議論ですよね。どんなストーリーなのだか興味があるので、紹介してもらえませんか。たぶん、論文自体はドイツ語ですよね。直メールででも、コメント欄への書き込みでもけっこうですので、解説していただけるようでしたら、このサイトで紹介させていただきたいと思います。
承諾の方はなにも問題ございません。事実と反していないかどうかは確認しておいた方がよいかもしれませんが。
ご存じのとおり、Milner and Goodaleの議論は心の哲学ではいろいろな人に採りあげられている話題だと思います(すぐ思いつくところでAndy ClarkやAlva Noë)。このへんの話をすることが出来たら、とてもうれしいです。
あと、ここだと目立たないので、現在のトップページにもこのコメントを貼り付けておきました。
2004年01月07日
■ Duhamel @ CNRSと
さしで話ができるようなので、論文読みまくり。この人はneurologistでもあり、frontalからparietalへcorollary dischargeが来ている可能性をBrain '92に出している。また、neurophysiologistでもあり、LIPへのmuscimol injection、LIPのreceptive field mappingなどの仕事があり、かつてはGoldbergとScience '92を出し(これがいわゆるpresaccadic remappingの初期バージョンと言える)、GrafとVIPニューロンのreceptive fieldに関してNature '97を出している。Duhamelのサイトへのリンク。
2004年01月06日
■ 頭頂葉
の機能を考えるためには内部モデルについての理解が必要となり、川人先生とWolpertの話を理解しなければならないことが判明。たいへん。Duhamelがもうすぐ来るし、この辺を固めるチャンスなのかもしれない。
2003年12月25日
■ 宿題
- GlimcherのNatureとAnnual reviewを説明してPlattのNeuronにコメントする
- 松元さんのScienceについてコメントする
- Goldberg vs. Andersen
2003年12月20日
■ 19日のコメント欄
に書くことが長くなったのでこちらへ。まとまらない。
19日のJNS Fig.1,2を眺めているとこんなことを考える: それでも、encoderはdecodeされることを前提として作られなければならないはずで、encodeされる刺激とdecodeされたestimateを突き合わせて誤差を修正していく過程というのはdecoderだけではなくencoderにも必要なのではないだろうか。発達時に形成されながら、完成後も常に修正を受けながら存在しつづけるencoderとdecoder。
Decoderがencoderと別のレベルにあるようにわれわれには見えるのはわれわれが実験者として外からdecodeしようとしているからだけで、たぶん記述のレベルが違うのだ。Encodeするのは生物自身であって、decodeするのは実験者。Encodeの研究は、知覚刺激を生物自身がencodeしたスパイクというspecifyされた実体を実験者が記述するだけであるのに対して、decodeの研究では、生物がそれをどう利用(nearly equals decode)しているかではなくて、知覚世界像のような充分specifyされてないものを実験者がスパイクから作り上げようとする(= decode)のであって、それは記述ではない(>>このため、取捨選択が必要になる)。
動物や我々自身はspikeにecodeされた知覚情報を最終的な行動としてアウトプットするわけで、そのときには単なるdecodeをしているわけではないが、何らかの形でそのスパイクの情報を利用し、変換しているとは言える。この「内的な変換過程」みたいなものを見ようとするのが私が今まで書いてきた「内側からの視点」を計算論的に解釈したものといえるかもしれない。ああ、こんな曖昧なことしか言えない。
2003年12月08日
■ NYU
Center for Neural Science, New York Universityのメンバーは
Glimcher / Ledoux / Movshon / Simoncelli。
全員有名。Nature, Scienceはみんな持ってる。
Glimcher自身はNature以降reviewしか最近は書いてない。
2001年04月02日
■ 「記憶力を強くする」書評
池谷裕二君のブルーバックスの書評をbk1とamazon.co.jpに書く。正直なところ面白く書けているので熱烈に誉め、少し批判する。「脳と記憶の謎 遺伝子は何を明かしたか」山元 大輔著や「学習する脳・記憶する脳 メカニズムを探る」磯 博行著と比べて書くという計画は頓挫。1600文字では無理だ。
書評タイトル:記憶の脳内メカニズム研究の最新の知見を含んだ入門書。おすすめ。
書評:★★★★★
池谷裕二さんとは大学、大学院を通じての同僚なので、その分差し引いて読んでほしいが、この本は記憶に関する神経科学の最新の知見を興味深く読ませてくれる良い本だ。
まず、構成がよい。海馬が記憶に重要であることから始まって、最新の知見を散りばめ、スクワイアやタルビングの心理学的枠組みへとつなげる。エピソード記憶と意味記憶の関係とかは本当はいろいろややこしいのだけど、うまいことストーリーが流れている。教科書丸写しではなくて、よく消化してから書いている証拠だ。このあとに神経細胞、シナプスについての記述があって、LTPとは何かが説明される。いきなり本の最初から神経細胞の説明に入ったらうんざりだから適切な構成だ。そして6章の「科学的に記憶力を鍛えよう」に入っていく。
そしてこの6章が面白い。実際のところ、ここで書かれていることは先述のエピソード記憶、意味記憶、手続き記憶などの枠組みを使った話であって、「最新脳科学が語る」というほどのことではない。けれども池谷さんの経験と信念がにじみ出た人間味あふれる文章になっていて魅力的だ。たとえば、「どの科目でも優秀な成績をとることができる学業の優れた人は、一つの科目すらもマスターしていない人から見ると超人的な天才に見えますが、しかし、それは生まれつき頭がよいというよりも、むしろ、いろいろな科目の学習能力が相乗しあった結果なのです。(216、7ページ)」なんてのは家庭教師をしていた学生に教えてあげたいセリフだ。
ツッコミどころを探してみた。ベートーベンの「運命」とシータ波の関係にはヲイヲイって感じだし、ヴィトゲンシュタインの「語りえぬものについては沈黙しなければならない」という言葉の捉え方はおかしいぞと思ったし、記憶力の累積の効果はいいけど生物にはS字カーブもあるぞ、とも思ったけど、この本の良さを損なうものではない。
もう一点コメントしておくならば、7章の「天才ネズミ「や「記憶力を増強する薬」の可能性については私は懐疑的だ。老化などによる機能低下を抑えるようなものはありうるだろう。けれども正常な海馬全体に対する操作でできることは限られていると思う。神経細胞一つ一つがそれぞれに別の情報をもっていて、それが集団として働いているのが脳システムだ。これに作用を及ぼすためには、脳がどういう情報を扱い、操作しているかが明らかにならないとわからないのではないか、これが私の考えであり、私がいま生理学をやっている理由の一つでもある(ちなみに私は記憶の「再生」に関する研究をしているラボ(259ページ)に所属している)。まあ、とはいえ、歴史からすれば、メカニズムより先に薬が見つかるのなんてのはあたりまえだし(精神分裂症とレセルピンとか、他のほとんど全てについても)、面白いニュースを待ってます、というのがフェアな態度か。(それでも正常からの増強ってのはね、、、アンフェタミンやプロザックをその例としてよいだろうか。)
この本の重要さを一つ指摘しておかなければ。この本には2000年あたりの国際科学雑誌の報告がてんこもりだが、これらが日本語で紹介されている一般向けの本は私が知るかぎりこの本だけだ。しかも羅列的でない。題材の取捨選択と配置がうまいのだと思う。人やサルの研究に関する言及が少ないのは専門家としては不満だが、1冊の本に全てを詰め込むことはできないからちょうどいい線だと思う。
この本は、新しい報告がどんどん出る分野を扱っているがゆえにそのうち古くなってゆくだろう。だから池谷さんにはあと5年たったらまたアップデートした本を書いていただけたらよいと思う。この本を記憶の脳内メカニズム研究の現状に興味のある全ての人に薦めます。
2000年08月19日
■ グレゴリー・ベイトソン(Gregory Bateson)の「精神と自然」まとめ
Chap.2 学校の生徒だったらみんな知ってる
認識論についてのベイトソン流「基礎の基礎」。
-
科学は何も証明しない。
科学は過程を向上させたり反証を加えることはできるが、それ自身の正しさを証明することは出来ない。これはポパーの考え方と同じ。オッカムの剃刀でより単純な答えを正しいと思っているだけ。原文ではこういってる。"Science probes; it does not prove." -
地図は土地は別物。名前と名付けられたものは別物。
記号論。シニファンとシニフィエ。客観的経験というものはない。
経験とは全て主観的なものである。足を踏まれたときに経験するのは「足を踏まれたこと」そのものではなくて「神経報告を元に再構成された足を踏まれたことに対する私の印象」である。イメージの形成は無意識過程である。
我々は知覚の過程に入り込んでいくことは出来ない。我々が意識するのは知覚の産物だけである。錯覚の実験でわかるように、遠くにあるものは小さく、近くにあるものは大きく見える、という法則が意識しないところに組み込まれている。知覚された世界が部分に分かれるのは必然かもしれないが、その分かれ方は必然ではない。
ある多角形を記述する方法は複数ある。この「記述」は必然的に恣意性を持つ。このような「記述」を元に「説明」がなされる。[発散する連鎖]を予測することはできない。
全体の振る舞いを知ることは出来ても、個に関する言明は出来ない。カオスの発想。[収束する連鎖]を予想することはできる。
発散と収束は異なる論理タイプに属する。発散は個を扱い、収束は集団を扱う。確率法則とは、(大数の法則による収束の原理を使って)個と集団という異なる論理タイプに属するものを結びつけるものである。論理タイプ:ラッセルの考え。命題の論理タイプを考えることで自己言及によるパラドックスを回避しようとした。無からは何も生まれない。
a. エネルギー保存の法則。 b. パストゥール:生命なしに生命を生み出すことは出来ない。 秩序、パターンは情報なしには作り出せない。数と量とは別物である。
数はパターンとゲシュタルトとデジタル計算の世界に属し、量はアナログ計算、確率計算の世界に属する。量によってはパターンは決まらない。
量とパターンは別の論理タイプに属する。生物学に単調な価値はない。
どんな薬にだって致死量はある。小さいことはいいことだ、ということもある。
四倍体の馬の話。サイズには関連する他の変数によって決まる最適値がある。ゾウの時間、アリの時間、だっけ?論理は[因果のモデル]としては不十分だ。
サーモスタット、ブザーの話。論理には時間が含まれていないのに、因果には時間が含まれている。因果の方向が逆転することはない。
そのため、目的論的過ちを犯す。しかし、原因と結果が循環的であるとき、どちらも互いの原因になっているということができる。卵が先か鶏が先か。言葉は通常、相互作用の片面だけを強調する。
主語Aが述語Bという性質を持っている、という表現はAがその内的、外的及び観察者との関係によって規定されていることを無視しやすくする。[安定している][変化している]という言葉は記述のうちの一部分のみを表している。
生きているシステムにおいて、安定している、とは何か他のものが変化しているからであり、この言葉が属する論理タイプを明確にする必要がある。
Chap.3 世界の重なりを見る
複数の情報が組み合わされるとより多くの理解が得られる。
-
差異
差異の知らせ(=情報)は、二つ以上のものの相互作用に差異が内在するときに、その二つ以上のものによって作り出される。 -
両眼視
片目ずつの情報の重ねあわせによって奥行きという情報が生まれる。つまり、片目からの情報とは別の論理タイプに属する情報が生まれ、視覚に新しい次元が加わる。 -
冥王星の発見
ゆっくりとした動きを見付けるためには、時間上の異なった瞬間の観察を比べる、という作業が必要になる。冥王星の小さなジャンプは恒星の経時的な位置を基準にすることで発見された。 -
シナプス加重
ニューロンAとニューロンBとが同時に発火するときのみ、両者の下流にあるニューロンCが発火する。つまり、加重という言葉は適当でなく、むしろ論理積ANDを行っている。 -
「マクベス」の幻の剣
マクベスは剣が幻であることを触覚では血は付いていなかったのに視覚では血が見えることから見やぶった。二つの感覚を比較することでメタ情報を得ている。 -
同義表現
(a+b)2=a2+2ab+b2は幾何学的に、一辺がa+bの正方形の面積を考えるとよくわかる。しかしここでは新しいことが付け加えられているわけではない。しかしここで生徒は代数と幾何とが翻訳可能であることを発見している。 -
二つの性
生殖は単一の性で行われる場合と比べて、二つの性の分裂と融合による方法は個体の偏差を抑えつつ、遺伝子の組み合わせを変化させることを保証している。 -
うなりとモワレ
二つのリズムパターンが重ね合わされるとその差が第三のパターンを生み出す。二つの音を重ねると、二つの周波数の差がうなりとして聞こえる。二つの縞パターンを重ねると、二つの空間周波数の差がモワレとなる。 -
「記述」「トートロジー」「説明」
説明は記述以上の情報を与えることはないはずなのに、それ以上のボーナスがある。「説明」は「トートロジー」の上に「記述」をマップすることであり、「記述」と「トートロジー」との重ねあわせによって「説明」はより豊かになっている。
Chap.4 「精神過程」であることの条件
精神の過程mental processが他の物質的出来事とどう違うかを論ずる。
-
「精神」とは部分が相互作用してできる集合体である。
この書は全体論的であるが、まともな全体論はみな、部分の相互作用を基盤とする。 -
「精神」の各部分での相互作用は差異によってトリガーされる。
相互作用はエネルギーの受け渡しではない。 -
「精神過程」は付帯的なエネルギーを必要とする。
2.からエネルギーが必要でないといっているのではない。それぞれの部分がそれぞれの部分でエネルギーを必要としており、準備してある。 -
「精神過程」では決定要因が循環的に連鎖している。
フィードバックによって互いの決定関係の記述が一周すると、論理タイプが変化している。 -
「精神過程」では、「差異の効果」とは「それに先立つ差異をコード化したもの」のことである。
コード化のうち、「部分が全体の代わりコーディング」では部分を知覚してそこから全体を推測する。我々の生活での知覚がそうだ。 -
これらの情報の変換プロセスを記述し分類するとしたら、その現象の論理タイプの階層構造を明らかにすることになる。
論理タイプを移り渡れるかどうか。パブロフの犬、イルカの例、ダブルバインドの理論。
Chap.5 関係の重なりを見る
Chap.2に続いてこんどはそのメタの部分同士の重ね合わせを考える。佐藤訳ではそこは訳出できてない。ちなみにchap.3の原題はmultiple versions of the world、chap.5の原題はmultiple version of relationship。つまり、chap.7で出てくるprocessとformという形式をchap.3-6で模している。(追記20070630: 改訂版では「重なりとしての関係」となっている。)
-
「汝自身を知れ」
「汝自身を知れ」という言葉に従うことで新しい情報が加わる。また、自分の状態を知っていない方が問題解決がうまくいくこともある。「遊び」という例ではシステムAとBが内部の情報のみから学習をしている。 -
トーテミズム
「人間社会のシステム」と「自然をも含んだ生態システム」との重ね合わせで捉えること、これがトーテミズム。 -
アブダクション
例:カエルの体構造と他の動物の構造との同型性に気づく。これは複数の出来事の多重な記述をしていることになる。
Chap.6 大いなる「確率的過程」
「進化」と「学習」は本質的におなじ「確率的過程」であり、両者の違いはその過程の基盤が属する論理タイプの違いに過ぎない。
-
ラマルク説の誤り
ラマルク説の誤りとは、「獲得形質の遺伝」で受け渡される遺伝情報についての論理タイプの誤りである。 -
用・不用
用・不用の問題では、体細胞変化と遺伝的変化という別の論理タイプを渡っている。そのためには体細胞変化と遺伝的変化とはそれぞれがお互いの柔軟性が保てるように拘束しあっていることを考慮すべきである。 -
遺伝的同化
Conrad Waddingtonの実験。個体の体細胞変化と周りの生態系との共進化を考える。ここで、個体、個体群、共進化、という違った論理タイプが現れることになる。 -
遺伝的変化による体細胞変化のコントロール
体細胞変化の制御は「変化の能力」「変化の能力を変化させる能力」とメタ化することができる。(神経の場合、神経間の伝達を変える「神経の可塑性」に対して、「神経の可塑性」を変える「メタ可塑性」というのがある。)しかしこのメタ化は数段で遺伝子レベルに達する。そういう位置に遺伝子変化はある。 -
無からは何も生まれない―発生ヴァージョン
カエルの受精卵での極性の発生は精子の突入による。非対称であるということは対称であることよりも多くの情報を必要とする。「ベイトソンの法則再考」。 -
相同
トムソンのカニの形の例。動物の相同には違った種類のものがある。また、系統発生的相同はその中でより安定な擬似トポロジカルパターンによるものである。 -
適応と耽溺
「耽溺」とは破滅的結果をもたらす「適応」のことである。例として、短期的に見て好ましいことが、長期的には破滅的である例。これは論理タイプ間での価値の違いによる誤算が関係している。 -
「確率的過程」「発散的過程」「収束的過程」
遺伝的変化の発散的、確率的過程は発生の収束的システムによってバランスをとっている。 -
二つの「確率的過程」を比較し、組み合わせる
1)遺伝子変化にランダムな要素があり、それが生物の内的ストレスによって選択されてゆく。2)表現型と環境との関係に予測不能性=ランダムな要素があり、これは適応によって選択されてゆく。無からは何も生まれない。
この二つを組み合わせてわかることは、1)の遺伝子変化が抽象的、質的量的どちらでもありうるのに対して、2)の体細胞変化は直接的で全て量的である。またこの二つの「確率的過程」と「精神過程」を比較すると、「精神過程」も二つの「確率的過程」によっていることがわかる。1)「創造的思考」はランダムな要素をもっている。これは論理的一貫性のフィルターによって選択される。2)環境との関係のランダムな要素があり、これは学習などによって適応的に選択されてゆく。
さらにこの二つの「確率的システム」は「分類」と「過程」という形で組み合わされる。これはchap.7にて。
Chap.7 「分類」から「過程」へ
記号化する、とは「名づけられるもの」から「ものの名前」へと論理タイプをジャンプするものだ。形態と過程の関係はトートロジーと記述の関係に一致する。
-
「ナヴェン」の例
バリ島民の行動の記述(過程)は男女気質の類型化(形態、分類)にまとめられ、これは類型間の相互作用(過程)にまとめられる。このジグザグによって論理タイプを登っていくことになる。 -
エアコンの調節の例
調節される気温(量的変化)、エアコンの設定温度(閾値)、人が寒い/暑いと感じる(量的変化)と論理タイプを登る。 -
賢者は輪郭を見る
こうして過程のみを重視するものと分類のみを重視するものとは時間を捉えなおすことによって統一的に解決される。時間が関わるとき、不連続性が現れる。感覚と組織とコミュニケーションの世界はこのような不連続=閾値の存在なしには考えられない。
Chap.8 それでいったいなんなの?
野暮な物質主義を逃れる道は美だ。そして美と意識と神聖さの領域、これらを考える前の問題として、精神、トートロジー、差異、の問題を論じたのがこの本だ。
2000年01月10日
■ 書評:金沢創著『他者の心は存在するか』(金子書房:1999)
まずは非常に面白かったです。最終章での著者の世界観を一通り作り上げるところまであっという間に持っていかれて、話の展開には必然性を感じました。
構成についてひとつ言うと、2章は知っていたことも多いし、1章での問題提起、そして本の題名といったところで問題追求型の話の進め方をしているはずなのにまだるっこしいと思いました。強く言ってしまえば、この章はなくても話は通じるし、その方が訴える力は強かったと思います (この章でいいたかったことが後の章にかかっていることは承知しておりますが)。逆にいえば、この本はいろいろ詰め込んであるような印象があるが、ここを除くと結構一本道で進みやすいか、と。
大切な5章の結論についてですが、主観的世界が始まりであり、進化的には、そこから外部世界が出来上がり、他者が出来上がり、自己が出来上がる、というのは私にもスムーズに受け入れられます。スタート地点は主観的感覚世界なのは確かで、そういう内部の視点を大切にする、という考えが私がオートポイエーシス論に共感する理由でもあります。しかし、こういう立場はここから先を進むのに苦労するんじゃないかと思います。それは現象学しかり、オートポイエーシスしかりで。なぜ、他者が同じような認知構造を持っていて、実際コミュニケーションができてしまうのか、というあたりに心身問題は視点の逆転によって隠された形で入っていると思うのですが、ここについて著者が[この一歩を踏み出してはいけない」(p.219)というとき、単に出発地点に問題を押し込んだように見えてしまいます。こういうのは「哲学」がまた始まる場所だと思うんです。
違ったレベルの話ですが、結局著者は「進化による説明」(2章より) に終始して、他者のメッセージがどうやって自分に信憑を与えるのか、などについての機構、機能のレベルでの説明が足りない(誰もできているわけではないと思いますが)のではないかと思います。このあたりが私は「心身問題は偽問題」という主張が証拠不充分だと思う理由のもうひとつです。これらの説明のためには、ニューロン、脳レベルでの説明が必要になるのではないかと思います。試しにひとつ考えてみたのですが、カエルの目を手術で180度回転させておくと、カエルは餌の位置に対して180度回ったところに舌を伸ばすだけで、外部世界と脳内表現の対応を修正できないらしいです(スペリーの実験など)。これに対して、ヒトやサルでは逆さメガネの実験(下條信輔氏の本など)で順応できてしまうことからわかるように、外部世界と脳内表現の関係を修正することができる。だから、カエルはレベル0の外部世界を持っていない状態、ヒト、サルはそれを持っていて、少なくともレベル1以上であるようです。そしてこのことはおそらく感覚―運動連関のfeedbackの有無などの形で機構として明らかにしていくことができるのではないだろうか、と思います。このような機構による説明を隅々まで行き渡らせることができたら、問題はまた違った形を見せる感じがします。
著書の締めが複数の宇宙、となっていて、どうも閉じた感じが気になって指摘したかったのですが、p.217での、「感覚情報を元に別のリアリティーを構成しうるような体系を作り出すことができれば、その枠の外に出られるのかもしれない」という点がqualia-ML #1689 で強調されていたのを見て、結構納得いってしまった感じです。
結論としましては、主観的感覚世界からスタートすることには共感を憶えました。そしてその結論が必然性を持っていると思いました。その先がデッドエンドな感じが正直言ってするのですが、ぜひここから先を進んでいくところを見せていただきたい、と期待を込めて思いました。
お勧めエントリ
- 細胞外電極はなにを見ているか(1) 20080727 (2) リニューアル版 20081107
- 総説 長期記憶の脳内メカニズム 20100909
- 駒場講義2013 「意識の科学的研究 - 盲視を起点に」20130626
- 駒場講義2012レジメ 意識と注意の脳内メカニズム(1) 注意 20121010 (2) 意識 20121011
- 視覚、注意、言語で3*2の背側、腹側経路説 20140119
- 脳科学辞典の項目書いた 「盲視」 20130407
- 脳科学辞典の項目書いた 「気づき」 20130228
- 脳科学辞典の項目書いた 「サリエンシー」 20121224
- 脳科学辞典の項目書いた 「マイクロサッケード」 20121227
- 盲視でおこる「なにかあるかんじ」 20110126
- DKL色空間についてまとめ 20090113
- 科学基礎論学会 秋の研究例会 ワークショップ「意識の神経科学と神経現象学」レジメ 20131102
- ギャラガー&ザハヴィ『現象学的な心』合評会レジメ 20130628
- Marrのrepresentationとprocessをベイトソン流に解釈する (1) 20100317 (2) 20100317
- 半側空間無視と同名半盲とは区別できるか?(1) 20080220 (2) 半側空間無視の原因部位は? 20080221
- MarrのVisionの最初と最後だけを読む 20071213