[月別過去ログ] 2005年08月
« 2005年07月 | 最新のページに戻る | 2005年09月 »2005年08月31日
■ ハウルの動く城ペーパークラフトの無料ダウンロード今日まで
息子とハウルの動く城ペーパークラフトを作ってます。8/31まで無料ダウンロード。でもA4のPDFで26枚もあるもんだから、印刷代とインク代だけで2000円はかかるという罠。やるなエプソン。でも私はCANON派。しかもそうそう完成までたどり着けません。こちらのハウルの動く城 ペーパークラフト組立日記 プラスα(おすすめ、というかここを見ないと作れません)を見ながら作っているのだけれど、この方も20日かかってる。私の方はというと、げんざい、骨格がだいたいできたところ。ここからいろんなパーツをくっつけてゆくのですが、まあ、息子が飽きるところまではやってみるつもり。

以前は紙飛行機のペーパークラフト(よく飛ぶ紙飛行機〈Vol.1〉―切りぬく本 切りぬく本のシリーズ)など作って公園で息子といっしょに飛ばしてたりしたのだけれど、作りきってしまった。しかし、せっかく完成させても、扱いが荒いからか、一日くらいでちゃんと飛ばなくなってしまうのです。補強するテクを身につけてまたやってみようか。
なんにしろ、そういうわけで家に帰ると私にはなつやすみのしゅくだいがあるのでした。
というエントリを作って放置していたのだけれど、松代に行くときに思い立って、紙飛行機を作って持っていってみました。しかし、飛ばす場所がないのでホテルで飛ばしていたという私。いや、なんとなく高原をイメージしていたのだけれど、松代は古くからの城下町だった、というオチで。
2005年08月30日
■ The robot said, "this is what it is like to be a bat!"
■ モンテカルロ(2)
昨日のつづきを貼ります:
昨日のコードを使って、どのくらいのループ回数でどのくらい真の値に収束するか確認してみました。
mloop=1000;sloop=5;loop=10^sloop; data2=zeros(log10(loop),2*mloop); nummat=10.^(1:sloop); tic; for m=1:mloop n=randn(1,1);meanN=n;meanNp=n;ssN=0;ssNp=0;stdN=0; data=[]; for i=2:loop n=randn(1,1); meanN = (meanNp*(i-1)+n)/i; ssN = ssNp + (i-1)*(meanNp^2) -i*(meanN^2) + n^2; stdN = sqrt(ssN/(i-1)); meanNp=meanN;ssNp=ssN; if ~isempty(find(nummat==i)) data=[data;i meanN stdN]; end end data2(:,2*m-1:2*m)=data(:,2:3); end toc; k1=log10(abs(data2(:,1:2:size(data2,2)-1))); k2=log10(abs(1-data2(:,2:2:size(data2,2)))); figure hold on plot(log10(nummat),median(k1'),'ro-') plot(log10(nummat),median(k2'),'bo-')
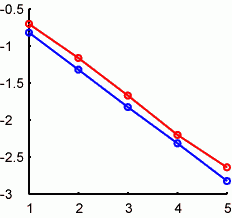
横軸がループの回数(log10(x))。縦軸は計算された値と真の値との差(絶対値)をlog10スケールにて。赤が乱数のmeanによるもので、abs(mean(data))で、青が乱数のsdでabs(1-std(data))です。昨日の計算を10000回繰り返して横軸の各点でのmedianをとっています(非対称分布なのでmean is not = median)。Meanとsdのどちらも直線近似できて、slopeは1/2なのです。これはたぶん差の絶対値だからで、差の二乗を取ればslope=1になったのでしょう。
以上の計算をしなくてもなんかのやり方で導出できるのでしょうけれど(そういうのってなにを読めばわかるのだろうか)、こういうことがわかりました:ループの繰り返し回数を10倍にすると真の値からのズレはsqrt(1/10)=1/3.16だけ小さくなる。
ところでこういうことやってよいか。とくに端数の問題、浮動小数点の問題。全データを用意しておいて、全データで一挙に計算したときとループごとに計算したときで誤差がどのくらいあるか検証してみる。
なんてことを考えて放置したままになっていたのでそのまま貼っておきます。
# ryohei
SE~SD/sqrt(N)になるのと同じ理屈ですね。Nこランダムな数字(が平均)の平均を何回も計算して平均をとると、もちろんが平均。分散はだいたい)^2> = =/Nとなります。(それぞれの事象が独立の場合、クロスタームがすべてゼロになる)。物理実験実習であたりで習った覚えがありますが、多分どの統計の本にも載っているのではないでしょうか。
# ryoheiしまった、< とか>を使うときに注意しなければいけなかったんだっけ。えーと、気を取り直してもう一度、Nこランダムな数字の平均を何回も計算してその平均をとると、もちろん<x>が平均。分散は<(Σxi/N - <x>)^2> = <(Σδx)^2/N^2 >=<δx^2>/Nとなります。(それぞれの事象が独立の場合、クロスタームがすべてゼロになる)。これでやっと書けている感じがします。上のできそこないのコメントは消しといていただけますでしょうか。
# pooneilそっかあ、「SE~SD/sqrt(N)になるのと同じ理屈」、これは明解でした。どうもありがとうございます。meanのmedianを取っているのだけれど、たぶん、数が大きいので、medianをとってもmeanをとってもほとんど変わらなかったのでしょう。
2005年08月29日
■ モンテカルロ(1)
昨年くらいに途中まで作って放置してあったエントリを貼っておきます。しかも「間違ってる!」なんてメモまで付いてる。なんだったか忘れたので埋め草用に。しかも明日に続きます。
モンテカルロでなんかの値のmeanとsdとを計算するために10万回とかループを回して、全部の値を行列に収納してからそれのmeanとsdとを計算していたのですが、それではおっつかなくなってきたので、ループごとにmeanとsdとをアップデートしてゆくような漸化式の形を作るようにしました。そんなことすらサボってたわけです。
平均:MEAN; 変動(sum of square):SS; 標準偏差:SD; として、 MEANn = (MEANn-1 * (n-1) + newdata )/ n SSn = SSn-1 + (n-1) * (MEANn-12) - n * (MEANn2) + newdata2 SDn = sqrt(SSn / (n-1) )
たとえば、randnでmean=0、sd=1の正規分布の乱数を生成させるのを10^8回繰り返してその乱数のmeanとsdを計算してやると、
tic; nummat=10.^(1:8);data=[];loop=10^8; n=randn(1,1);meanN=n;meanNp=n;ssN=0;ssNp=0;stdN=0; for i=2:loop n=randn(1,1); meanN = (meanNp*(i-1)+n)/i; ssN = ssNp + (i-1)*(meanNp^2) -i*(meanN^2) + n^2; stdN = sqrt(ssN/(i-1)); meanNp=meanN;ssNp=ssN; if ~isempty(find(nummat==i)) data=[data;i meanN stdN]; end end toc; figure hold on plot(log10(data(:,1)),data(:,2),'ro') plot(log10(data(:,1)),data(:,3),'bo')
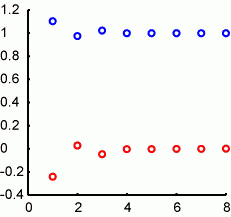
わたしのノートPCではelapsed_time =18975secで計算されます。図の通り、横軸がループの回数(log10(x))。縦軸がそれぞれのループの回数で計算された値で、赤が乱数のmeanで、回数が増えるほどに0に近づきます。青が乱数のsdで、回数が増えるほどに1に近づきます。10^5=10万回くらいやればだいたいよさそうなことがわかります。
ほんとうはもっと良いのは、matlabが得意とする行列での演算と組み合わせることで、
std(randn(10^4,10^3)); elapsed_time = 2.0160
なんてので10^4個の乱数のsdが10^3個計算できるので、それを前述の漸化式のように組み合わせたほうが10^7個の乱数のsdを計算するのはもっと速くなることでしょう。ちなみに直で計算させると
std(randn(10^7,1)); elapsed_time = 165.8200
メモリーが圧迫されるのでけっこう時間がかかってしまいます。さらに、うちのノートPCではもう一桁増やすと
std(randn(10^8,1)); ??? エラー: ==> randn メモリが足りません.HELP MEMORYとタイプしてオプションを確認してください
なんて出てきて、もう計算できません。
2005年08月26日
■ 論文いろいろ
Cerebral Cortex 7月号
Basal Ganglia and Cerebellar Inputs to 'AIP' -- Clower et al. 15 (7): 913 -- Cerebral Cortex
Localization of the Human Cortical Visual Area MT Based on Computer Aided Histological Analysis -- Annese et al. 15 (7): 1044 -- Cerebral Cortex
Brain 7月号
"Somatosensory activations during the observation of touch and a case of vision–touch synaesthesia"
Journal of Cognitive Neuroscience '04
"Neural Correlates of First-Person Perspective as One Constituent of Human Self-Consciousness"
以上の二つはvia Mochi's-Multitasking-Blog
- / ツイートする
- / 投稿日: 2005年08月26日
- / カテゴリー: [Paper archive]
- / Edit(管理者用)
2005年08月25日
■ Cannabis and the brain
"Cannabis and the brain" Leslie Iversen, Brain, Vol. 126, No. 6, 1252-1270, June 2003
THE MOLECULAR LOGIC OF ENDOCANNABINOID SIGNALLING Daniele Piomelli, Nature Reviews Neuroscience 4, 873-884 (2003); doi:10.1038/nrn1247
"Cannabinoid physiology and pharmacology: 30 years of progress" Allyn C. Howlett, Christopher S. Breivogel, Steven R. Childers, Samuel A. Deadwyler, Robert E. Hampson and Linda J. Porrino, Neuropharmacology, Volume 47, Supplement 1, 2004, Pages 345-358.
"The endogenous cannabinoid system and the treatment of marijuana dependence" Daniele Piomelli, Neuropharmacology, Volume 47, Supplement 1, 2004, Pages 359-367. Nature Reviews Drug Discovery 3, 771-784 (2004); doi:10.1038/nrd1495
THE ENDOCANNABINOID SYSTEM AND ITS THERAPEUTIC EXPLOITATION Vincenzo Di Marzo, Maurizio Bifulco and Luciano De Petrocellis
"Long-term effects of exposure to cannabis" Leslie Iversen, Current Opinion in Pharmacology, Volume 5, Issue 1, February 2005, Pages 69-72.
"Emerging properties of cannabinoid medicines in management of multiple sclerosis" Gareth Pryce and David Baker, Trends in Neurosciences, Volume 28, Issue 5, May 2005, Pages 272-276.
2005年08月24日
■ 論文いろいろ
時間がない。以前メモって放置してたやつを貼っときます。
Neuron 6/2
Peter J. Bayley, Jeffrey J. Gold, Ramona O. Hopkins and Larry R. Squire, The Neuroanatomy of Remote Memory, Neuron, Volume 46, Issue 5, 2 June 2005, Pages 799-810. Wendy SuzukiがF1000で挙げてた。
John-Dylan Haynes, Jon Driver and Geraint Rees, Visibility Reflects Dynamic Changes of Effective Connectivity between V1 and Fusiform Cortex, Neuron, Volume 46, Issue 5, 2 June 2005, Pages 811-821.
Implicit Attentional Selection of Bound Visual Features
Nature 6/2
Oxytocin increases trust in humans
Human behaviour Brain trust
JNS 6/8
Neural Basis for a Powerful Static Motion Illusion -- Conway et al. 25 (23): 5651 -- Journal of Neuroscience 表紙にもなってます。
JNP 7月号
Statistical Issues in the Analysis of Neuronal Data
Neuron 5/19
Uncertainty, Neuromodulation, and Attention
- / ツイートする
- / 投稿日: 2005年08月24日
- / カテゴリー: [Paper archive]
- / Edit(管理者用)
2005年08月23日
■ JNP 9月号
"Chronometry of Visual Responses in Frontal Eye Field, Supplementary Eye Field, and Anterior Cingulate Cortex" Pierre Pouget, Erik E. Emeric, Veit Stuphorn, Kate Reis and Jeffrey D. Schall
Visual Response latencyをFEF,SEF,ACCで比較した。以前のJNP '98 "Signal Timing Across the Macaque Visual System"の続編みたいなもん。このJNP '98はLGN-V1-V2-V3-V4-MT-MST-FEFでvisual Response latencyを比較した、というもので、データは寄せ集めでしかないのだけれど、こうやって比較したものはほかにないので貴重な資料。Response latencyは視覚刺激が最適なものであるかどうかに大きく依存する、とかいろいろつっこみどころはあるにせよ。FEFのほうがV4よりもlatencyが早い、とか重要なことが書いてある。
前回のデータと照らし合わせてみると、SEFはFEFよりは遅くて、V2と同じくらい。ACCはSEFよりもさらに遅くて、V4と同じか遅いくらい、というかんじらしい。
- / ツイートする
- / 投稿日: 2005年08月23日
- / カテゴリー: [上丘、FEFと眼球運動]
- / Edit(管理者用)
2005年08月19日
■ 松代の統合脳班会議
松代の統合脳班会議に参加してきます。諸先輩方や同年代の若手研究者と熱く語ったり、飲み過ぎたりしてくる予定。
# nak
どうでしたか?
よろしければご感想をお聞かせください。
正味一日しか参加してない(計画班のプレゼンと公募のポスターセッション)のですが、忘れぬうちになにか書こうと思ってます。しばらくお待ちください(来週くらいになりそうです)。
■ CSSでmax-width設定
メインコンテンツの横幅が広すぎるというのも読みにくいものなので、CSSでmax-widthを640pixelに設定しました。FirefoxとOperaで有効でIEでは変化ありません。そのぶんサイドバーを広げたいのだけど、うまくいかん。とりあえず放置。(サイドバーのwidthは%ではなくてpixelで設定して、そのぶん本文のカラムは右のマージンを空ける、という方式にしてます。)2005年08月18日
■ extracellularで記録されるsingle-unitの波形つづき
昨日のつづきです。生理学の教科書ではホジキン-ハックスレーの仕事で細胞内記録と膜チャネルの動態の話はだいたい出てくるのですが、私がこのサイトで挙げている論文で使われるような、細胞外電極によるsingle-unitの記録の原理というものはちょうどいい教科書がない、という話でした。
詳しいことは置いといて、おおまかな骨組みだけまとめておきましょう。三段階の説明のレベルがあって、それらを関連づけられるようにすればよいのかな、と考えました。第一段階は膜電位のdipoleが作る電界によるsinkとsourceを計測する、という説明。第二段階はaxon方向の一次元での等価回路を使った近似。第三段階は電流密度の連続の方程式を使った三次元での取り扱い。
まずは一番プリミティブかつ直感的な説明は、Lorente de Nó 1947などを元にした、dipoleがaxonを伝播してゆくモデル。Action potentialが起こると、一つの細胞のaxonの中で膜電位の変化が起こり、細胞膜を通した電流の流入とその周りでの流出とでループが出来る。これは細胞膜にdipoleができてそこを中心とした等電位曲線が描ける、ということ。細胞外電極はこの電位を測定している。等電位曲線に垂直な電気力線が細胞内へ流入してるところが吸い込み:sink(オシロでは下向き)で、流出がわき出し:source(上向き)。脱分極と過分極の二つのdipoleを考えると細胞外記録でのポジ-ネガ-ポジの三層性の電位変化が再現できる。
第二段階のaxon方向の一次元での等価回路では、細胞外電極を置いたところでsinkが起こったのをキルヒホッフの電流保存の式を使って定式化。細胞外電位をaxon方向の一次元で二回微分するとその場所での細胞内への電流の流入、流出が算出できる。これがcurrent source density analysisの原理。膜電位変化を時間で二回微分すると膜電流変化になる(Tasaki I)、というのもここからの帰結。Rall Wのcable theoryもこのレベルでのこと。私自身もむかしじぶんで勉強したときはこのレベルまでだったと思う。細胞内膜電位を時間で一回微分すると細胞外電位になる、というのもこのレベルの話のはずなんだけど、元ネタ見つからず。
第三段階の電位ポテンシャルに関するラプラス-ポワソン方程式を使った三次元での取り扱い。電荷がない点では電流のsinkもsourceもないので電界の発散はない(divE=0)。膜電流のあるところでの電荷密度の時間変化は電界の発散に比例する(連続の方程式)。このレベルで、さまざまな仮定が第一段階や第二段階で置かれていることを明示的に扱えるようになる。現在のモデル屋さんはこのレベルを扱っている。ベクトル解析で電磁気学と流体力学の対応とか読んでるとおもしろいけど、生物系の教養課程ではそのへんまでたどり着いていなかったような。
んでもって、これらのレベル間での対応を付ける。各レベルでのsinkとsourceの扱いとか、第二段階でのキルヒホッフの電流保存の式と第三段階での連続の式との対応付けとか。 第三段階の式からcurrent source densityの式を導出とか。
とだいたいこんな感じになるのではないかと。前回書いたエントリでは、この第二段階が抜けていたからギャップを感じていたのですな。むかし勉強したことをすこし思い出してきました。
これを肉づけして、かんたんに説明できる資料が作れたらいいかな。ここで書いていることの完成版を作るのに私より向いている人はたくさんいるはずですし、どなたかぜひ。
Evoked field potentialとかLFPのoscillationとかを議論するにはまだ遠そうです。むかしラット海馬のslice cultureでevoked field potentialのLTPをやってたんですけど、そのときにはSchaffer collateralの入力層で記録すればfield EPSP(ネガ一層性)、pyramidal cell layerで記録すればpopulation spike(ポジ-ネガ-ポジの三層性)、と納得いってた気がするけど、somaでのcurrentのvolume conductionとか、dendriteへのaction potentialのpropagationとか考えるとじつはかなり深い。当時は疑問に思ってなかったけど、なんでfield EPSPにはsomaのspikeのvolume conductionがかぶってこないんだろう。Population spikeのポジにはdendriteでのEPSPのvolume conductionがかぶっているなんて理解してたけど、それでは対称性がない。それとも充分離れていれば、それこそextracellular mediumがinhomogeneousだからspikeのような速い周波数成分はさっさと減衰してしまうということか。海馬のような解剖学的に均一な構造ですらこんな調子なのだから、ましてやcortexとかは私にとっては複雑すぎ。
ところで私自身はこのへんのバックグラウンドが全然ないままにこれを書いているので、かなり恥をさらしています。つっこみ、訂正歓迎します。たとえば、ガウスの法則の微分形式と、ラプラス-ポワソン方程式と、連続の方程式の関係とか……本質的に同じなんだろうと思うのですが、系統的にそういうことを勉強した経験がありません。第二段階でのキルヒホッフの電流保存の式と第三段階での連続の方程式との関係とかも。物理のバックグラウンドを持っている方からすれば、教養課程レベルの知識もないことがモロばれでしょう。アンプを自作していた時代の生理学者から見れば嘆かわしいことに違いありません。まあ、なにより自分の勉強のため、ということで明日のために。
つづきがあります:20080727
2005年08月17日
■ extracellularで記録されるsingle-unitの波形
LFP関連ですが、研究所のトレーニングコースで、LFPのできるメカニズムを簡単にレクチャーできるようにと思っていろいろ資料を集めてたのですが、これが意外に難しい。そもそも教科書的にはホジキン-ハックスレーでイカのaxonからintraとかそういう話になってしまっていて、extracellularで記録されるsingle-unitの波形がどうできるかを説明している教科書というものはわたしが知る限りではよいものがありません。かたやsourceとsink、といった端的な説明の仕方(*1)になってしまうし、かたやラプラス・ポアソン方程式になってしまう(*2)。なんかその中間をいくようなうまい説明の仕方はないか思案中です。というわけで、いまさら電磁気学の勉強です。おすすめの資料がありましたら推薦歓迎します。
*1: たとえば、Roger Lemonの"Methods for neuronal recording in conscious animals"にあるように。
*2: Mitzdorfのcurrent source density analysisの論文 physiological review '85 "Current source-density method and application in cat cerebral cortex: investigation of evoked potentials and EEG phenomena"(pdf)とか、Alain DestexheのBiophysical Journal '04 "Modeling Extracellular Field Potentials and the Frequency-Filtering Properties of Extracellular Space"(著者サイトのpdfファイル)とか。
追記:STさんありがとうございます。推薦論文をリンクしておきます:
"Intracellular features predicted by extracellular recordings in the hippocampus in vivo." Henze, D., Borhegyi, Z., Csicsvari, J., Mamiya, A., Harris, K. and Buzsáki, G., 2000. J Neurophysiol. 84, 390-400.
"Electrical Interactions via the Extracellular Potential Near Cell Bodies". G. R. Holt and C. Koch. Journal of Computational Neuroscience 6:169-184, 1999
A Critical Reexamination of Some Assumptions and Implications of Cable Theory in Neurobiology.(pdf file) Holt, GR Ph. D. Thesis, California Institute of Technology, Computation and Neural Systems Program (1998)
上記のDestexhe論文ではKoch and Segevの"Methods in Neuronal Modeling" 1998がLFPの簡単なモデル(extracellular mediumがhomogeneousであることを仮定)の例としてreferされてます。
次回へ続きます。
# ST
最近の資料として以下のようなものはいかがでしょうか。
Henze, D., Borhegyi, Z., Csicsvari, J., Mamiya, A., Harris, K. and Buzsaki, G., 2000. Intracellular features predicted by extracellular recordings in the hippocampus in vivo. J Neurophysiol. 84, 390-400.
Electrical Interactions via the Extracellular Potential Near Cell Bodies. G. R. Holt and C. Koch. Journal of Computational Neuroscience 6:169-184, 1999.
確かHoltのdissertaionもWeb上にあったような。
STさん、ありがとうございます。さっそくリストに追加しておきました。とくにBuzsákiのほうは、in vivoの皮質でのintraとextraを比較して提示するのによい資料になりそうです。
2005年08月11日
■ Anticorrelated random-dot stereograms
重要なのだけど、とりあえず論文リストだけ作成。
- Nature "Responses of primary visual cortical neurons to binocular disparity without depth perception" CUMMING AND PARKER
- Perception '98 "Disparity detection in anticorrelated stereograms" Cumming BG, Shapiro SE, Parker AJ.
- JNS "Rejection of False Matches for Binocular Correspondence in Macaque Visual Cortical Area V4" Seiji Tanabe, Kazumasa Umeda, and Ichiro Fujita
- JNP "A Stereoscopic Look at Visual Cortex" Peter Neri
- Vision Research "Unconscious adaptation: a new illusion of depth induced by stimulus features without depth" Ryusuke Hayashi, Yoichi Miyawaki, Taro Maeda and Susumu Tachi
- Vision Research '93 "Depth in anticorrelated stereograms: effects of spatial density and interocular delay"
- / ツイートする
- / 投稿日: 2005年08月11日
- / カテゴリー: [Papers_unclassified]
- / Edit(管理者用)
2005年08月10日
■ 論文いろいろ
Current biology 7/26
"Blinking Suppresses the Neural Response to Unchanging Retinal Stimulation" Davina Bristow, John-Dylan Haynes, Richard Sylvester, Christopher D. Frith and Geraint Rees
"Vision: In the Blink of an Eye" David Burr
"Bodily Illusions Modulate Tactile Perception" Patrick Haggard
Neuron 8/4
"Dissociation between Ventral and Dorsal fMRI Activation during Object and Action Recognition" Ehud Zohary
"Action Insight: The Role of the Dorsal Stream in the Perception of Grasping" Melvyn A. Goodale
"Testing the Efficiency of Sensory Coding with Optimal Stimulus Ensembles"
JNP 8月号
"Delay-Period Activity in Visual, Visuomovement, and Movement Neurons in the Frontal Eye Field" Lawrence H. Snyder。去年のSFNに出てた。Recording siteをVan EssenのCARETを使ってunfoldして表示している分は新しく作ったらしい。
"Neuronal Activity Dependent on Anticipated and Elapsed Delay in Macaque Prefrontal Cortex, Frontal and Supplementary Eye Fields, and Premotor Cortex" Carl R. Olson
"Topographic Organization for Delayed Saccades in Human Posterior Parietal Cortex" David J. Heeger
"Topographic Maps of Visual Spatial Attention in Human Parietal Cortex" David J. Heeger
"Supplementary Motor Area Encodes Reward Expectancy in Eye-Movement Tasks" R. A. Andersen
"Differential Characteristics of Face Neuron Responses Within the Anterior Superior Temporal Sulcus of Macaques" Wania C. De Souza, Satoshi Eifuku, Ryoi Tamura, Hisao Nishijo and Taketoshi Ono
- / ツイートする
- / 投稿日: 2005年08月10日
- / カテゴリー: [Paper archive]
- / Edit(管理者用)
2005年08月09日
■ fMRIのBOLDシグナルと神経活動
Science 8/5でback-to-back。
"Hemodynamic Signals Correlate Tightly with Synchronized Gamma Oscillations" Wolf Singerたち。麻酔下のcatではBOLDシグナルはスパイクよりはLFPのgammma oscillationのpowerの方によく相関していた。
"Coupling Between Neuronal Firing, Field Potentials, and fMRI in Human Auditory Cortex" Rafael Malach。去年出たintersubject correlationの手法を用いて、awake humanの聴覚野ではBOLDシグナルがspikeとよく相関している(r=0.75だと!)ことを強調しています。Gamma frequencyのLFPもBOLDとよく相関している、としているので前者の論文や以前のLogothetisらの論文とかと直接対立するものではなさそうです。
ちょうど1年前のエントリ(20040809)で関連する事項を扱っています。Logothetisの論文についてとか。ご隠居の最後のコメントで言及されているLauritzenらの仕事をリストしておきましょう:
- JP '98 "Modification of activity-dependent increases of cerebral blood flow by excitatory synaptic activity and spikes in rat cerebellar cortex"
- JP '00 "Temporal coupling between neuronal activity and blood flow in rat cerebellar cortex as indicated by field potential analysis"
- JNS '03 "Brain function and neurophysiological correlates of signals used in functional neuroimaging." (Review)
- / ツイートする
- / 投稿日: 2005年08月09日
- / カテゴリー: [Papers_unclassified]
- / Edit(管理者用)
2005年08月08日
■ 10万visitありがとうございます
昨年12月にsakura.ne.jpに引っ越してきてから累計で10万visitになりました。見に来てくださった方、どうもありがとうございます。
以前と比べて、訪問者数というのはあまり意味をなさなくなっていると思いますし(RSSやアンテナで見ている人もいる)、そもそもvisit数というのは正確なものではありませんし、ダメ押しで言えばこの数の中には私自身も含まれるわけですが、それでもなんかうれしかったりします。
かつてのNiftyでのサイトのカウンタが5年かけてやっと1万に達したとか、はてなダイアリーで書き始めた頃には10人/dayくらいしか読者がいなくて、目標は100人/day、とか書いていたことからすると感慨深いです。
今後も無理せずエントリを蓄積していく所存です。
と、いちばん上に来ないようにこっそりエントリを追加。
2005年08月05日
■ 三河ラーメン談義つづき
ローカルネタ。ひさびさにラーメンたじまに行ってみるとその日のメニューは「酢豚麺」とこれまたトリッキーなものだったので、すかさず頼んでみました。酢豚のあんがスープに混ざってもそんなに濃くならないようにバランスを取ってあって、スープも飲めてしまう。豚もおいしかったのでけっこうおいしく食べることが出来ました。でも前に食べた、すじ肉入りのラーメンをまた食べたいなあ、と思ってカウンター正面を見ると、「好評のため、再登場 すじラーメン 650円」とか書いてあるではないですか。おお!私以外にも好評だったんですか!ということでこんどまた行こうと思います。
……んでもって、今回もまたガラ空き。というか入ってから出るまで(8時半から9時まで)客は私一人だけ。ホント、つぶれないでほしいです。
関連する前回のエントリはこちら:20050512
追記:愛する妻の友人から「たじま」の場所の問い合わせが来る、というオモシロ展開に。ということで場所などの情報はこちらへどうぞ:「ラ王への道」での記述
2005年08月04日
■ Science Anosognosia(疾病否認)論文
そういえば昨年のエントリ(20040708)でPassinghamのScience '04 "That's My Hand! Activity in Premotor Cortex Reflects Feeling of Ownership of a Limb"に言及しましたけど、これも関係してきますな。自分の腕に対するfeedling of ownershipがあるかどうかはpremotorが関わってくる、というのは今回のScience論文とつなげて考えると納得のいくかんじがあります。つまり、半側空間無視と疾病否認との関係にさらに「身体失認」を入れて考えるということですな。あと、PassinghamのScience '04のPerspectivesが"Probing the Neural Basis of Body Ownership"にあります。
追記:最新のJNSにも関連論文あり。"Awareness of the Functioning of One's Own Limbs Mediated by the Insular Cortex?" Hans-Otto Karnath。例の、hemineglectの原因部位はparietalではなくてtemporalだとするNature論文の著者です。(参考:Spatial hemineglectスレッド)
Patients with anosognosia for hemiplegia/hemiparesisとhemiplegic/hemiparetic patients without anosognosiaとを比較して、その損傷領域の差はright posterior insulaであるとしてます。
- / ツイートする
- / 投稿日: 2005年08月04日
- / カテゴリー: [Papers_unclassified]
- / Edit(管理者用)
2005年08月03日
■ Chapterhouseのbreather聴きてー
夏ですな。摂氏35度とかになってましたよ。こう暑いときはガンガンに冷房かけた車でシューゲイザー系のノイジーなギターを大音量で聴きたいなあ、というわけで、Chapterhouseのbreather聴きてー。エビ喰いてー。しかし実家に置いてきてしまって聴けない。で調べたら中古が値上がりしてるとか言うてるではないですか。いろいろリンク。
- Chapterhouse「Whirlpool」の熱いレビュー。
- シューゲイザーコーナーの一つとしての記述。
- なにげなファンサイト。Breatherの歌詞聞き取りがあるけど、穴だらけ。いや、シューゲイザー系ってのは歌詞が聴き取りにくいように歌うんで仕方ないんですが。
- Amazon.comの長文レビュー。
- SHOEGAZING概論(たぶん)。
ところでシューゲイザー系とはなにか。上記のサイトをごらんいただければわかりますが、私なりに: 自己表出をしたいという欲望と、ショービジネス的エンターテイナー的な表現はしたくないという自己意識とのせめぎわいを解消するために、髪を伸ばして目を隠して下を見てうつむきながら(shoegazer=靴を眺めてる)大音量でギターを弾いて、ドリーミーかつ起伏の少ないメロディーを歌詞が聞き取れないようにギターの音に埋没させるようにして歌う、というスタイルをとったイギリスのギターバンドたち。1990-92年あたりに最盛期を迎えた。なんか書いてて最近の無_断リ_ンク_問題(昔から言われているやつではなくてね)に通じる自意識問題の存在に気づいてみたり(とか書いて下書き状態でこのエントリを放置しておいたらもうその話題は終わっていた!)。(さらにこの「髪を伸ばして目を隠して」とadvゲームの「主人公問題」とを重ね合わせることも可能。)
2005年08月02日
■ Nature 7/28
"Robust habit learning in the absence of awareness and independent of the medial temporal lobe" Larry R. Squire
Medial temporal lobeを両側で損傷している患者E.P.さんと患者G.P.さんはdeclarative memoryを保持することが出来ないにもかかわらず、concurrent discrimination taskを繰り返すことで成績がよくなってゆく。
患者E.P.さんに関してはこれまでもSquireグループが研究してきました。以前の「記憶の脳内機構」スレッドにも関連記事があります。
また、しげさんのところの20050728のエントリに詳しい記載がありますので、そちらへどうぞ。
- / ツイートする
- / 投稿日: 2005年08月02日
- / カテゴリー: [内側側頭葉と記憶システム]
- / Edit(管理者用)
2005年08月01日
■ 線条体とQ-learning
20050730のエントリにmmmmさんからコメントがついています。ATRの鮫島さんのポスターに関してですが、mmmmさんは線条体がActor-CriticではなくてQ-learningであることを生理学的に証明したことを評価しています。
鮫島さんが学会で発表されたことについては大脳基底核-強化学習モデルによる線条体神経細胞活動の記録と予測(pdf)で一部公表されているようなのでこれに基づいて議論しますが、この論文ではこの点が明確に書かれています。つまり、Actor-Criticだと線条体で行動選択そのものがrepresentされていると予言するのに対して、Q-learningでは線条体で行動選択肢の報酬予測がrepresentされると予言するというわけです。なるほど。
ニューロンのデータに基づいた議論はつづきの論文が出てから、としたほうがよいでしょう。
ところで、そもそもわたしはActor-CriticとQ-learningの概念がぜんぜんわかっていないので、参考になりそうな資料をメモっておきます。
ATRの鮫島 和行さんによる関連論文:
- 大脳基底核-強化学習モデルによる線条体神経細胞活動の記録と予測(pdf)
- 強化学習と大脳基底核(pdf)(実はこの論文は以前にも20040624でリンクしてます。)
- 行動決定系列からの学習系の内部状態/パラメータ系列の推定(pdf)
- 複数の予測モデルを用いた強化学習による非線形制御(pdf)
Sutton and Bartoの"Reinforcement Learning: An Introduction"のhtml版から:
東工大の木村 元氏によるもの:
- 強化学習の基礎(pdf)
- 平成13年度 (2001年度) 7月6日講義資料(PowerPointファイル)
- 強化学習システムの設計指針
同志社大の中村 康昭氏によるもの:
- / ツイートする
- / 投稿日: 2005年08月01日
- / カテゴリー: [行動の価値 (action value)]
- / Edit(管理者用)
お勧めエントリ
- 細胞外電極はなにを見ているか(1) 20080727 (2) リニューアル版 20081107
- 総説 長期記憶の脳内メカニズム 20100909
- 駒場講義2013 「意識の科学的研究 - 盲視を起点に」20130626
- 駒場講義2012レジメ 意識と注意の脳内メカニズム(1) 注意 20121010 (2) 意識 20121011
- 視覚、注意、言語で3*2の背側、腹側経路説 20140119
- 脳科学辞典の項目書いた 「盲視」 20130407
- 脳科学辞典の項目書いた 「気づき」 20130228
- 脳科学辞典の項目書いた 「サリエンシー」 20121224
- 脳科学辞典の項目書いた 「マイクロサッケード」 20121227
- 盲視でおこる「なにかあるかんじ」 20110126
- DKL色空間についてまとめ 20090113
- 科学基礎論学会 秋の研究例会 ワークショップ「意識の神経科学と神経現象学」レジメ 20131102
- ギャラガー&ザハヴィ『現象学的な心』合評会レジメ 20130628
- Marrのrepresentationとprocessをベイトソン流に解釈する (1) 20100317 (2) 20100317
- 半側空間無視と同名半盲とは区別できるか?(1) 20080220 (2) 半側空間無視の原因部位は? 20080221
- MarrのVisionの最初と最後だけを読む 20071213