[月別過去ログ] 2021年05月
« 2021年04月 | 最新のページに戻る | 2021年06月 »2021年05月16日
■ 自由エネルギー原理FEP関連メモ
FEPの説明法を見直していた。期待自由エネルギーEFEにいきなり飛ぶのは難易度が高いので、VFE = Uncertainty + Bayesian surpriseの式を使ってきた。これは不正確な方便のつもりだった。でも、行動aからサンプルされる $s$ が一意に決まるときはEFEにせずに、VFEの式でよいと自信を持った。
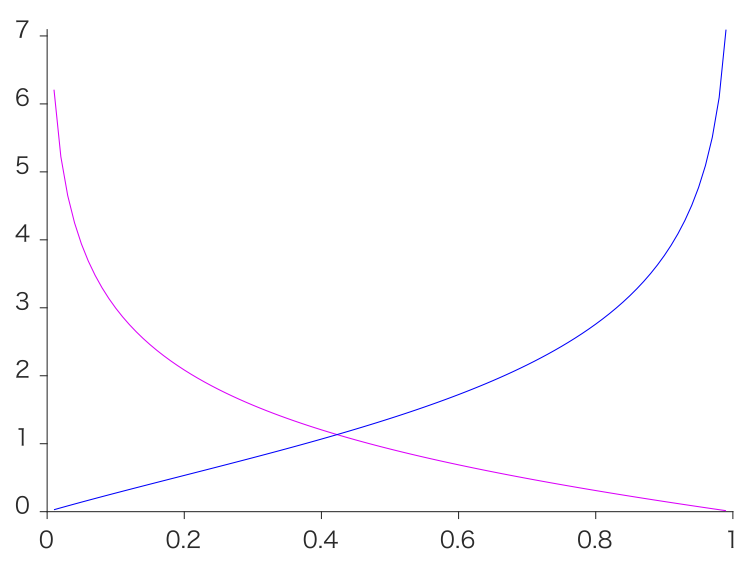
横軸に $p(s=1)$ ($s=1$:左下を見ていて蝶はぼやけてる画像; $s=2$:右上の帳を注視している画像)とおいて、posteriorは固定しておくと、$s=1$ でのUncertaintyはマゼンタ。つまり $p(s=1)<0.42$ で $s=2$ をサンプルする。つまりもし $p(s)$ を自由に動かせるなら、ありふれた $s$ を選ぶ。暗い部屋問題を再現できた。
今のはposterior $p(x|s)$ を固定して、それに $p(s)$ を掛けて生成モデル $p(x,s)$ を作っていた。このように $p(s)$ にアクセスできるなら、暗い部屋問題は起こる。しかし、実際は $p(s)$ は生成モデル $p(x,s)$ を周辺化したモデルエビデンス $p(s|m) = \sum_{x} p_m(x,s)$ なので、agentと環境との関係によって生成モデル $p_m(x,s)$ のほうが先に決まる。(ここでは、生成モデルの形を決めるパラメーターを $m$ と書いている)
そうか、「もっともありふれた刺激 $s$ 」とか言わずに、モデルエビデンス $p(s|m)$ の最大化、と言えばいいのだな。(FEPのような)BIC的なアプローチの代わりにAIC的なアプローチがありうる、って話まで持っていけそう。ぜんぶつなげて話すと、FEPの話ではなくて、ふつうにベイズ統計の授業だなこりゃ。
変分自由エネルギーFはモデルエビデンスの-logの下限(ELBO)なので、自由エネルギー最小化とは、$F_1-F_2 \approx -log(BF_1/BF_2)$ となっていて、ベイズ因子によるモデル選択と同じになる。知覚よりも行動選択の場合だと、$p_{a1}(x,s)とp_{a2}(x,s)$ という2つの生成モデルの比較がしっくりくる。
以前書いた気がするけど、「FEPはなんでも最適化主義ではないか問題」は進化における適応主義論争と形式的には同じで、「理不尽な進化」では適応主義という研究プログラム自体は否定し得ない(最適化というフレームワークで検証をすることの意義)ということでグールドが敗北したことになっているけど、カウフマンが外適応の概念を拾っているのを見て、いやエナクティヴィズム的には再考するお余地があると思った。
「理不尽な進化」も「脳がわかれば心がわかるか」も非常にデネット的なのだけど、グールド敗北で終わらない動機とか、回帰する擬似問題、といったところを残す点がこの2冊では共通してる。
この話への正確なツッコミとしては、「定式化しようと思えば定式化できる(定式化の妥当性、答えの有無に関わらず)」という意味であれば、それは無意味な言明で、それ以降の主張を支える役割を果たしてない。ここで必要な文章は端的に「制約付き最適化って重要だよね」ってことだけだと思う。
2021年05月01日
■ スチュアート・カウフマンの「生命はいかにして複雑系となったか」を読んだ
カウフマンの"Beyond Physics: The Emergence and Evolution of Life"についてのトークのスライドPDFがCERNでのトークの資料として入手できる。このスライドを使ったWisconsin Institute for Discoveryでのトークの動画。字幕はないけど。
前述のスライドのp.3 “Thus “functions” are legitimate categories in science in the historical becoming of the non-ergodic universe above the level of atoms. “Functions” do not exist in physics, which cannot discriminate “functional subsets” of causal consequences.”
ツーソン会議2018ではこれの短縮版のトークの動画がある(35分くらい)。こちらは字幕がある。こっちを視聴した。
カウフマン「生命はいかにして複雑系となったか」、さっそく立ち読みしたら、思ったより薄くて、内容は前述のスライドそのままっぽいことがわかったので購入してきた。さっそく読みだしたら、まさに私が知りたいことだった。
まとめておくと、宇宙の歴史の中でまだ可能なタンパク質の配列のすべてが作られたわけではない、そういう非エルゴード的宇宙において、進化においてどのように生物がエコロジカルニッチを見つけて進化してゆくかはあらかじめ予見することはできない。
(物理での運動方程式が初期条件と拘束条件が必要なのに対して)そもそも生物の進化においては、そのような拘束条件自体が新たに作り出されてゆく。しかもそのような「応急処置」は多様性が増すと可能性が増える。
よって、これから何が起こるかわからない(コインの裏表)どころか、何が起こりうるかすらわからない(確率測度を定式化するための標本空間が決まらない)。
このような非エルゴード的世界を記述するには物理的な説明ではなくてお話としての説明しかない(これは以前私がブログに書いた話とは逆の帰結)。だから、生命のあるこの世界は物理学を基礎とはしているけれども、それに還元されるものではないのだ、というのが結論。
ここからは私の感想だけど、物理的な説明に対するお話としての説明(その生き物にとっての「意味」を通した説明)の優位を強調するのは驚きだったけど、追求する価値はあるなと思った。これこそが人文知が必要となる、必然性の高い道筋なのではないだろうか?これはVarelaの相互的拘束よりも強い要請だ。
そんなわけで、いますごく興奮している。以前からちょっと読んでは積んでた「自己組織化と進化の論理」、あと分厚くて敬遠していた「カウフマン、生命と宇宙を語る」これらを読み進めてみようと思う。
補足: まあこれまで私は、そういう「物理を越えている」部分を情報として捉えた上で、情報は熱力学的な意味で物理とつながる、っていう理屈を想定していたのだけど、一歩立ち止まって、「一回性の世界」をもっとよく見てみよう、というのが今の心境。
追記: あと、ダーウィンの「前適応」の概念よりもグールドの「外適応」のほうを採用しているし、生態学的ニッチの意義の強調も含めて、カウフマンは非常にエナクティブな視点に立っている。進化の意義を強調しても、デネット、ドーキンス的な方向以外があり得る、このことは重要。
「カウフマン、生命と宇宙を語る」借りてきた。ざっと見、新著とそんなに大きく変わっているわけではない。あるプロセスの結果が別のプロセスの拘束条件になるって話もある。ただし、それがループとして閉じるとなってないだけ。
あと、熱力学の第4法則として生物進化を(統計的に)予測するようなものがあると「カウフマン、生命と宇宙を語る」では議論しているけど、新著ではそういう予測はあくまでも統計的なものであって、生命はbeyond physicsなのだ、という立場に立っている。新著では熱力学的な仕事(カルノーサイクル)への言及は最小限だったが、旧著ではひとつの章を当ててる。
カウフマンの"world beyond physicsで「束縛閉回路」という話が出ていた。つまり、火薬の爆発という非平衡なプロセスを砲弾を飛ばすという仕事にするためには大砲という拘束条件が必要となる。生物ではこのような拘束条件の連鎖がループして閉回路になっている、
つまり代謝の反応の連鎖で細胞壁を作って、これが拘束条件となって酵素による触媒反応が可能な程度に気質の濃度を高くする。このようにしてオートポイエーシスにおける作動的閉包をプロセスと束縛条件に分けて捉えている。
それで、カウフマンの本ではこれらが平衡から遠く離れた状態のプロセス、と読んでいたのだけど、物理化学の教科書とか読んでると、これはふつうに個々の反応はギブス自由エネルギーで取り扱えるような減少で、そのかわり、ATPとか生産物とかがその都度出入りしているけど、それは化学的平衡の速度と比べたらもっとゆっくりしたものであるということ。
そこまで見てみると、よくある「生物とは平衡からは遠く離れた状態を維持している」というフレーズが、時間の早いプロセスと遅いプロセスに分けることでかなり化学的な平衡の話で語れることであることを加味して理解しないといけないのではないかとか思った。
シュレディンガーの「生命とは何か」に出てくる「負のエントロピーを食べる」という話があって、いまそれも読み直しているけど、これの書いてある6章の注記(後日追加された)には、負のエントロピーとは正確には自由エネルギーのことであるが、一般聴衆にはエネルギーという言葉が誤解を与えるのでエントロピーという語を使ったと書いてある。そうするとここでの「秩序から秩序を作る」という話は端的にさっき書いたようなATPからADPに戻るときの大きなギブス自由エネルギーの減少ΔG1(<0)を使って、それとカップルしているリン酸化反応による秩序の形成(-ΔG1>)ΔG2(>0)を作れるという話と読める。
(もちろん遺伝子の情報がこのような回路を作るために働いていること込みで。) このような(カウフマンの言う)束縛閉回路については「エントロピーから読み解く 生物学-めぐりめぐむ わきあがる生命-」佐藤直樹に図があった。
ここはちがうか。化学反応での扱いは、系に対してその周り全体を環境として扱うから、全体としては孤立系として扱っているため、ということか。それに対して、生物でのローカルな環境(細胞の中のさらに局部)で物を見ようとすると、物質の出入りがあるから、非平衡として扱わないといけなくなる。
お勧めエントリ
- 細胞外電極はなにを見ているか(1) 20080727 (2) リニューアル版 20081107
- 総説 長期記憶の脳内メカニズム 20100909
- 駒場講義2013 「意識の科学的研究 - 盲視を起点に」20130626
- 駒場講義2012レジメ 意識と注意の脳内メカニズム(1) 注意 20121010 (2) 意識 20121011
- 視覚、注意、言語で3*2の背側、腹側経路説 20140119
- 脳科学辞典の項目書いた 「盲視」 20130407
- 脳科学辞典の項目書いた 「気づき」 20130228
- 脳科学辞典の項目書いた 「サリエンシー」 20121224
- 脳科学辞典の項目書いた 「マイクロサッケード」 20121227
- 盲視でおこる「なにかあるかんじ」 20110126
- DKL色空間についてまとめ 20090113
- 科学基礎論学会 秋の研究例会 ワークショップ「意識の神経科学と神経現象学」レジメ 20131102
- ギャラガー&ザハヴィ『現象学的な心』合評会レジメ 20130628
- Marrのrepresentationとprocessをベイトソン流に解釈する (1) 20100317 (2) 20100317
- 半側空間無視と同名半盲とは区別できるか?(1) 20080220 (2) 半側空間無視の原因部位は? 20080221
- MarrのVisionの最初と最後だけを読む 20071213