[月別過去ログ] 2007年09月
« 2007年08月 | 最新のページに戻る | 2007年10月 »2007年09月28日
■ 意志決定の曖昧さ1
研究会予習シリーズも時間的にこれが最終回。私が座長をする予定になっている、東大先端研の渡邊克巳さんの講演に関して予習することにします。研究会要旨集で参考文献を挙げてくださってます。このへんから行ってみましょう。
- Watanabe, K. (in press) "Behavioral speed contagion: Automatic modulation of movement timing by observation of body movements." Cognition.
- Science 7 October 2005: Vol. 310. no. 5745, pp. 116 - 119 "Failure to Detect Mismatches Between Intention and Outcome in a Simple Decision Task" Petter Johansson, Lars Hall, Sverker Sikstr ö m, Andreas Olsson
- Dijksterhuis, A., Bos, M. W., Nordgren, L. F., van Baaren, R. B. (2006). "On making the right choice: The deliberation-without-attention effect." Science, 311, 1005-1007.
とりあえず2番目の論文から読み始めました。Science 7 October 2005: Vol. 310. no. 5745, pp. 116 - 119 "Failure to Detect Mismatches Between Intention and Outcome in a Simple Decision Task" Petter Johansson, Lars Hall, Sverker Sikström, Andreas Olsson。ファーストオーサーのPetter JOHANSSONという人はもともとスウェーデンの人だけど現在は渡邊研に在籍しているようです。
わたしたちは日常生活で意志決定をしてなにかを選ぶわけだけど、その意志決定と選択との関係がどのくらいあやふやかを示した論文です。
やってる実験はひとことで言えば、手品。被験者の前で実験者が顔写真が書いてある二種類のカードを左右の手に持って見せます。被験者にはどっちが好きか選んで指さししてもらう。被験者は実験者から選んだほうのカードを引いて、なんでそのカードを選んだかを答えてもらいます。んで、トリックが仕掛けられていて、たまに指さしたものとは反対の手のあったカードが被験者に手渡されます。ここが手品なわけですね。手に持ったカードを二枚重ねにしておけばいいわけです。
さて、被験者はカードがすり替えられたことに気付くか。これがなかなか気付かない。その場で気付くのはせいぜい5%とか。
では、気付かなかったときはその写真についてどうコメントするか。だって、こっちがいいと思って選んだはずなのに逆を選んでいて、しかもそのことに気付いていないんだから、なんでそれを選んだかコメントするのが難しいはずで。ひとつのパターンとしては後付けで説明を作ってしまう。つまり、現在手に持っている写真の顔についてあれこれこういう理由でこっちを選んだ、と言ってしまう(confabulation=作話)。もしくは逆にはじめに選んだときの写真の顔についてコメントしてしまう。たとえば、「なぜ選んだかというと、こっちの写真は笑っているから」みたいなかんじで。手元にある写真は笑い顔ではないのに。
そういうわけでわれわれはそういう状況において自分が選択したもの、選択した理由についてけっこう見逃している。けっこう曖昧だということがわかるというわけです。著者はこの現象のことを"choice blindness"と呼んでいます。
これに似通った現象はいろいろ知られています。目撃証言にかんする記憶の曖昧さに関する話がありますね。Schacterの本とかで出てるやつ。事件の目撃報告とかでは証言がいろんな要素によってゆがめられる。写真のリストから現場にいたと思われる人を選んだら、無意識レベルの偏見によって選択がゆがんだ。これもおそらくは本人は意識しているわけではないのですね。嘘をついているわけではなかった。
あと、ラマチャンドランの本で出てくる印象深い病態否認の例。右腕が動かないということを否認するために、いまは動かしたくない、とか動いてないのに動いていると言い張るとか。これもたぶん嘘をついているわけではないのですよね。
あと、論理的不協和理論とも大いに関係ありそうですよね。論理的不協和理論は言ってみれば「酸っぱいブドウ」理論です。「おいしそうなブドウがある」という認識(=内的条件)と「ブドウが手に入らない」という状況(=外的条件)があって、この二つが整合的でないために論理的不協和(cognitive dissonance)という状態を体験する。これを解決するために、変えようのない状況の方ではなくて、「おいしそうなブドウがある」という認識の方を変えてしまう。「あれは酸っぱいブドウだった。」というわけです。ところで、もはやクリシェとなった「勘違いしないでよ、べつにあんたのためにXXしたわけじゃないんだからねっ」も同じような分析が可能ですね。余談ですが。
このへんの関連性については著者は今回のScience論文の一番最後のパラグラフで
Classic studies of social psychology have shown that telling discrepancies between choice and introspection can sometimes be discerned in group-level response patterns (12) but not for each of the individuals at hand.
と書いていて、このreference 12というのがNisbett and WilsonのPsychological Review 1977 Vol 84(3) 231-259 "Telling more than we can know: Verbal reports on mental processes."(PDF)というやつでして、この世界の古典的論文です。ここに書いてある、「選択と内観の不一致」に関してはまた後述しますが、ともあれ、これまでの社会心理学的な実験で示されたことを個人のレベルでも示すことが出来た、という点がこの論文のノイエスであるようです。
Science論文とこのニスベットとウィルソン1977との関連については、Science論文の続編のConsciousness and Cognition Volume 15, Issue 4, December 2006, Pages 673-692 "How something can be said about telling more than we can know: On choice blindness and introspection"(PDF)でかなりくわしく扱われています。
ちなみにこれにたいするPatrick HaggardによるコメンタリがConsciousness and Cognition Volume 15, Issue 4, December 2006, Pages 693-696 Commentary on ‘How something can be said about telling more than we can know: On choice blindness and introspection’ James Moore and Patrick Haggardで、それに対する返答がConsciousness and Cognition Volume 15, Issue 4, December 2006, Pages 697-699 "Reply to commentary by Moore and Haggard"です。
さて、Science論文の意義を評価するために、もうすこしいろいろ調べてみましょう。(というか調べてたら面白くなってきた。これは大いにわたしのやっていることと関係します)。渡邊克巳さんはERATOの下條潜在脳機能プロジェクトの意思決定グループのリーダーでもありまして、この論文の意義も「潜在脳機能」という側面から見るとよくわかります。下條先生の「サブリミナル・マインド」の第1講「自分はもうひとりの他人である」ではまさに上記のニスベットとウィルソン1977に言及しています。
下條先生の「サブリミナル・マインド」を読み直して、さらに社会心理学の本なども読んでいろいろ調べてきました。次回は「フェスティンガーの認知不協和理論」、「ベムの自己知覚理論」、「原因帰属理論」といって「ニスベットとウィルソンの"Telling more than we can know"」までたどっていきます。
- / ツイートする
- / 投稿日: 2007年09月28日
- / カテゴリー: [生理研研究会2007「注意と意志決定の脳内メカニズム」]
- / Edit(管理者用)
2007年09月27日
■ 実りある議論のためのリソース2
「実りある議論のためのリソース」つづき。第2日目の講演者の方に関してです。第2日目は全体として「意志決定」のパートとなっております。
5. 「情動に基づく意思決定のための大脳基底核関連回路」中原 裕之(理化学研究所 脳科学総合研究センター 理論統合脳科学研究チーム)
中原裕之さんは計算論神経科学をされておりまして、情報幾何をスパイク列に応用したお仕事(Nakahara, H., & Amari, S. (2002). "Information geometric measure for neural spikes." Neural Computation. 14(10): pp2269-2316.)で有名ですが、実験神経生理学者とも多くのコラボレーションを行って成果を出されております。今回は大脳基底核についてお話をされますが、それ以外にもattentional modulationによるtuningに関する仕事(Neural Computation 2001やNeural Networks 2002)や上丘のsaccadic burstに関わるlocal circuitに関する仕事(Journal of Neurophysiology 2006)などさまざまです。
- Nakahara, H., Itoh, H., Kawagoe, R., Takikawa, Y., & Hikosaka, O. (2004). Dopamine neurons can represent context-dependent prediction error. Neuron. 41(2): pp269-280.
- Nakahara, H., Amari, S., & Hikosaka, O. (2002). Self-organization in the basal ganglia with modulation of reinforcement signals. Neural Computation. 14(4): pp819-844.
6. 「社会的状況における意思決定のメカニズム」春野 雅彦(国際電気通信基礎技術研究所 脳情報研究所)
春野雅彦さんはMOSAICモデルの論文(Haruno M, Wolpert DM, Kawato M: MOSAIC model for sensorimotor learning and control. Neural Computation, 13 2201-2220 (2001))の著者として有名ですが、近年はhuman fMRIを使った理論の検証を進めておられます。今回発表される内容はこれまで発表されているものとは違うもののようですが、human fMRIのお仕事をリストしておきます。
- Haruno M, Kawato M: Heterarchical reinforcement-learning model for integration of multiple cortico-striatal loops; fMRI examination in stimulus-action-reward association learning. Neural Networks, 19, 1242–1254 (2006) .
- Haruno M, Kawato M: Different neural correlates of reward expectation and reward expectation error in the putamen and caudate nucleus during stimulus-action-reward association learning. Journal of Neurophysiology, 95, 948-959 (2006).
- Haruno M, Kuroda T, Doya K, Toyama K, Kimura M, Samejima K, Imamizu H, Kawato M: A neural correlate of reward-based behavioral learning in caudate nucleus: a functional magnetic resonance imaging study of a stochastic decision task. Journal of Neuroscience, 24, 1660-1665 (2004).
7. 「ヒトにおける金銭的価値の脳内表現 - 機能的MRIによる神経経済学的研究」筒井 健一郎(東北大学大学院生命科学研究科 脳情報処理分野)
筒井健一郎さんは日大酒田研時代のCIPでの3D surface codingのお仕事(Science. 2002 Oct 11;298(5592):409-12. "Neural correlates for perception of 3D surface orientation from texture gradient." Tsutsui K, Sakata H, Naganuma T, Taira M.)が有名ですが、現在は東北大に移動されて新しい仕事を始めておられます。今回の発表では、human fMRIでの神経経済学的アプローチでの成果をお話しいただきます。まったく新しい仕事ですので、参考文献は挙げておりません。
8. 「意思決定の適当さ」渡邊 克巳(東京大学先端科学技術研究センター(認知科学分野), (独)産業技術総合研究所, (独)科学技術振興機構)
渡邊 克巳さんは下條先生のところ、彦坂先生のところで仕事を出され、多彩な成果を出しておられます。ERATOの下條潜在脳機能プロジェクトの意思決定グループのリーダーでもありまして、意志決定に関する潜在脳機能にアプローチをされております。次回から予習シリーズで採りあげさせていただきます。以下は要旨にありました、参考文献のリストです。
- Watanabe, K. (in press) "Behavioral speed contagion: Automatic modulation of movement timing by observation of body movements." Cognition.
- Science 7 October 2005: Vol. 310. no. 5745, pp. 116 - 119 "Failure to Detect Mismatches Between Intention and Outcome in a Simple Decision Task" Petter Johansson, Lars Hall, Sverker Sikström, Andreas Olsson
- Dijksterhuis, A., Bos, M. W., Nordgren, L. F., van Baaren, R. B. (2006). "On making the right choice: The deliberation-without-attention effect." Science, 311, 1005-1007.
- / ツイートする
- / 投稿日: 2007年09月27日
- / カテゴリー: [生理研研究会2007「注意と意志決定の脳内メカニズム」]
- / Edit(管理者用)
■ 今日ネットで見た印象的なフレーズ
「シャワー浴びてると嫌な事思い出して壁パンチしたくなるよな?」
2007年09月26日
■ 実りある議論のためのリソース1
研究会予習シリーズですが、このペースでは全員回ることは不可能になってきました。残された時間で、私が座長をする予定の東大先端研の渡邊克巳さんの講演について予習を行うことにします。これはのちほど。
「実りある議論のためのリソース」と題して講演者のみなさまに関する参考文献などを挙げるコーナーを作成しました。まったくもってわたしの独断ですので、これで当たっているかどうかは保証の限りではありませんのであしからず。
まずは第1日目の講演者の方に関して。第1日目は全体として「注意」のパートとなっております。
1. 「脳損傷患者における注意と意思決定」 鈴木 匡子 (山形大学大学院医学系研究科 高次脳機能障害学)
鈴木匡子さんは今年から山形大の教授となられました。神経心理を専門とされていて、高次脳機能障害の患者さんの症例のレポートを多く発表されています。鈴木匡子さんは日本で数少ない、blindsightの患者さんのレポートをされた方でして(J Neurol Neurosurg Psychiatry. 2000 Jun;68(6):782-6. "Intact verbal description of letters with diminished awareness of their forms." Suzuki K, Yamadori A.)、わたしはその縁でお知り合いとならせていただきました。
-
BRAIN and NERVE 2007 (Vol.59 No.1) 「視覚性注意のしくみ」 鈴木 匡子
注意の分類、同時失認と半側空間無視についての症例、注意に関する脳部位、というまとめ方でとても役に立ちます。 -
Cortex. 2003 Apr;39(2):327-41. "Visuospatial deficits due to impaired visual attention: investigation of two cases of slowly progressive visuospatial impairment."(PDF) Suzuki K, Otsuka Y, Endo K, Ejima A, Saito H, Fujii T, Yamadori A.
Attention関連の症例報告。
2. 「注意のトップダウン制御原理 - 次元加重、課題構え、探索モード」 熊田 孝恒 (産業技術総合研究所 人間福祉医工学研究部門 認知行動システム研究グループ)
熊田孝恒さんは産総研で注意の心理物理をされている方です。サーチタスクのパラダイムを用いて、top-down attentionについて研究されていますが、脳損傷の患者さんを被験者とした研究も行っておられます。
-
Cognitive Neuropsychology, Volume 23, Number 3, May 2006 , pp. 401-423(23) "Deficits in feature-based control of attention in a patient with a right fronto-temporal lesion" Kumada, Takatsune; Hayashi, Mieko
こちらが患者さんでの研究の論文です。 -
Kumada, T. (2001) Feature-based control of attention: Evidence for two forms of dimension weighting. Perception & Psychophysics, 63, 698-708.
Feature searchで特定の刺激に重み付けするときにはWithin-dimension facilitation(WDF)とIntertrial facilitation (ITF)という二つのモードがあるという話です。
3. 「行動価値予測の誤差とリスク - 行動適応における前頭前野内側部の役割」 松元 まどか (理化学研究所 脳科学総合研究センター)
松元まどかさんのNature neuroscienceについては予習シリーズでとりあげました。今回の講演では、松元健二さんが神経科学大会のシンポジウムで話されたtop-down attentionの話が加わったものとしてお話しいただけるのではないかと思います。
- Nature Neuroscience 2007 "Medial prefrontal cell activity signaling prediction errors of action values" Madoka Matsumoto, Kenji Matsumoto, Hiroshi Abe and Keiji Tanaka
- Neuroscience Research 2007 "Effects of novelty on activity of lateral and medial prefrontal neurons"
4. 「注意が意思決定に変わるとき - 変換場としての頭頂連合野機能」 小川 正 (京都大学大学院医学研究科 認知行動脳科学)
小川正さんのお仕事についても予習シリーズでとりあげました。ちょっと尻切れトンボ気味でしたが。Visual searchのパラダイムを用いて、top-down attentionとbottom-up attentionがどのようなinteractしているのかということをニューロンの活動で示しています。V4, FEFのデータに関してはこれまでに発表されていますが、今回の講演ではLIPのお話も伺えるのではないかと思います。
-
The Journal of Neuroscience, 2004, 24(28):6371-6382; "Target Selection in Area V4 during a Multidimensional Visual Search Task" Tadashi Ogawa and Hidehiko Komatsu
こちらが初めに出てきた、V4からの記録によるもの。 -
Experimental Brain Research 2006 "Neuronal dynamics of bottom-up and top-down processes in area V4 of macaque monkeys performing a visual search" Tadashi Ogawa and Hidehiko Komatsu
こちらはFEFのニューロンのデータを付加して、ニューロンの発火パターンの時間経過を追ったものです。
次回は二日目、意志決定パートの方について書きます。
- / ツイートする
- / 投稿日: 2007年09月26日
- / カテゴリー: [生理研研究会2007「注意と意志決定の脳内メカニズム」]
- / Edit(管理者用)
2007年09月21日
■ 生理研研究会のサイトを開設しました・研究会について語ります
10/11-12は生理研研究会「認知神経科学の先端 注意と意志決定の脳内メカニズム」です。研究会のサイトを開設しました。プログラム、ポスターのリストなどを公開しています。ぜひごらんください。
近日中に要旨集のPDFを公開する予定です。ぜひあらかじめ読んで予習しておいていただければ。そういう意味ではこのブログも役立つはずです。
それから、研究会サイトでは[実りある議論のためのリソース]と題したページを作成中です。これにかんしてはまだ中身はありませんが、講演者の文献リストなどを作成する予定です。わたしの独断と偏見で、ということになりますが。
さて語る。もともとこの研究会は酒を飲みながら元 生理研 小松研の小川さん(現在 京大 河野研)といっしょに、仕事が出たら一緒に研究会をやろう!と約束していたのが実現したものです(まだわたしの仕事出てないけどorz)。
高次脳機能関連の研究会を行うにあたって、私たちが特色として打ち出したかったのは、横に広げるより縦に深く行こう、ということでした。ある分野に絞って(今回は「注意と意志決定」を選びました)、さまざまな実験アプローチの方に話をしていただこうと、小川さんとわたしとで演者になっていただく方にお願いして回りました。感激したのは、お願いした方みなさまに快く引き受けていただけたことです。
その結果、さまざまな分野の方に講演をしていただけることになりました。感謝しております。高次脳機能障害の患者さんを調べておられる鈴木匡子さん、計算論的神経科学でさまざまな実験神経科学者とコラボレーションしてこられた中原裕之さん、それからもちろんhuman fMRI, human psychophysics, monkey neurophysiologyの方にも入っていただけることになりました。もっとrodentsとか無脊椎動物とかin vitroとかのアプローチがあっても良かったかもしれない。そのへんは課題ということで。昨年のSFNで松元まどかさんの仕事を見て講演をお願いしたら、そのあとでタイミング良くNature Neuroscienceに掲載されるというニュースがあって、ガッツポーズをとったりしたものです。
深く行くというコンセプトのためにもう一つ考えていたのは、ディスカッションを重視することを形に見えるようにしたい、ということでした。生理研研究会の名物のひとつとして京大の金子武嗣先生の研究会(昨年のプログラム)っていうのがあるんですけど、これが半日*2で講演者が5人、割り当てがひとり1時間ということになっているんだけど、実際の講演時間は質問が尽きるまで延々続く、というものです。去年の狩野先生の講演とかも2時間くらいかかったのではなかったでしょうか。わたしがはじめにイメージしていたのはこの研究会でした。
ただ、ひとりで全員の演者に食らいつくというのは大変なことですので、ツッコミ役になっていただく方をお願いしよう、ということから「指定討論者」という枠を作って、いろんな方にお願いして回りました。今回公開されたプログラムを見ていただくとわかりますが、この指定討論者の方に座長になっていただいて、一人の講演者に対して二人の座長が付いている、という形になっております。そして、講演時間35分、議論時間20分をメドにプログラムを組みました。さあさて、これがうまくいくかどうかは当日のお楽しみ、というわけです。ぜひ見に来て(口調変わってるし)。
研究会のもうひとつの特色は、小川さんや私のような若手が企画・運営している点です。そういうわけで、若手の人に多く来てほしいし、ぜひ質問してほしいと思います。研究会や学会で質問時間があまりないと、エラい先生が質問してるのを聞いて終わってしまうということになりがちです。若手の人の質問を超encourrageします。「へんな質問かもしれないですけど」とか「基本的な質問ですけど」とかそういう枕詞はオミットでぜひ(エラそー)。
若手の人に来ていただくためのひとつの方策として、ポスターセッションを併設しました。また、ポスターセッションの会場でひきつづき懇親会を行うことにして、懇親会の会費を低めに抑えました(4000円、学生は2000円)。ぜひ酒飲みながらポスター前で語っていただけたらと思います。
小松研が毎年行っている視覚研究会(今年のプログラム)は懇親会を所内の食堂で行った後に隣の部屋にある和室に移動してみんなで胡座で酒を飲みながら語るというのがありまして、これが毎年盛り上がるのです。やっぱ胡座重要。参考にしたいなと思っていたのですが、今回はポスター会場で行う、という方を重視しました。つーか食堂をポスター会場にするってのはどうだろう? これはもし来年もやるとしたらのネタとして記録。
第一回の研究会ですのでいろいろ至らないところはあるだろうと不安なのですが(運営方面でいまはあれこれ大変です)、ぜひ多くの方に見に来てくださればと思います。どうかよろしくお願いします。
……今回はかなりぶっちゃけぎみに書いてみました。もし支障がありましたらご指摘ください。本当はこういうことは研究会が問題なく終了してから書くほうがよいことなのですが、それもこれも多くの人に見に来ていただきたいからです。この、現在進行形のプロジェクトの目撃者になってみませんか? (と書いたらさすがに照れた。)
- / ツイートする
- / 投稿日: 2007年09月21日
- / カテゴリー: [生理研研究会2007「注意と意志決定の脳内メカニズム」]
- / Edit(管理者用)
2007年09月19日
■ 生理研研究会とASCONEが近づいてまいりました
宣伝です。生理研研究会とASCONEが近づいてまいりました。
はげしくいそがしくなって分けわからない状態に。さて乗り越えられるでしょうか。
10/11-12は生理研研究会「認知神経科学の先端 注意と意志決定の脳内メカニズム」です。ポスター申し込みは9/14で締め切りました。たくさんのご応募どうもありがとうございました。参加申し込みの方は当日まで受け付けておりますのでぜひいらしてください。まもなくプログラム・要旨集が公表されますのでしばらくお待ちください。
10/5-8のASCONEのほうも着々と準備が。というか形をなすかどうかビビリながら準備中。ASCONEのページに要旨が掲載されました。よければそちらもご覧ください。講義が終了したらweb上講義に再構成したいなんて考えてますが、以前の大学院講義スレッドも宣言したまま放置中ですので時間がかかりそうです。
- / ツイートする
- / 投稿日: 2007年09月19日
- / カテゴリー: [生理研研究会2007「注意と意志決定の脳内メカニズム」]
- / Edit(管理者用)
2007年09月18日
■ ぼくが好きな音
はるはあけぼの、みたいな。誤解した音響派、とか予防線貼ってみたりして。
井上陽水の「断絶」に入っている「ハトが泣いてる」で、イントロのギターCコードかき鳴らしが止まってドラムのフィルが始まる直前に、ドラムスを録ってるマイクのフェーダーを上げたのか、部屋の空気(アンビエンスっていうの?)が聞こえるところ。
トッド・ラングレンの「サムシング・エニシング」の"cold morning light"で、ひとり多重録音してるもんで、ドラムがへたってリズムがばらつくところ。同じようなネタとしてローリング・ストーンズの60年代の曲(忘れた)とかボブ・ディランの60年代の曲(忘れた)とかもあるけどこれらはバンドでやってるんでニュアンスがちがう。
ザ・バーズの「霧の5次元」に入ってる"eight miles high"で、イントロのベースのフレーズに合わせてバスドラが共鳴してるところ。これは前にも書きました。これがはてなでのはじめてのエントリーだったっていう。
サイモン&ガーファンクルの「ブックエンド」に入ってる「動物園にて」の一番と二番の間で、ギターのアルペジオとベースだけが鳴ってるところの静かなかんじ。手拍子が入ったりとか。
はっぴいえんどの「はっぴいえんど」に入ってる「12月の雨の日」で、歌い出しが低くて声が出てないところ。
ニール・ヤングの"Everybody knows this is nowhere"の"cowgirl in the sand"で、長いギターソロのあとで歌がはじまる直前にカウントだかなんだかかすかに合図をしているのが聞こえるところ。
ニール・ヤングの「今宵その夜」で、ラストの「今宵その夜・パート2」で演奏が崩壊寸前になってるところで、なんとか持ち直して間奏に入ろうと合図を送るピアノの連打。
フランク・ザッパの「いたち野郎」に入ってる「ガスマスク序曲」でロイ・エストラーダが絶叫からうなり声へ、そしてバカ笑いに移行するときのタメの部分。
井上陽水の「氷の世界」が「あかずの踏切」から「氷の世界」まで半音ずつキーが下がってゆくところの曲の切れ間。とくに「チエちゃん」がはじまる瞬間。
よしだたくろうの「元気です。」の「馬」でボーカルが片側のチャンネルに寄ってて、反対側のチャンネルにエコーが振ってあるところ。
ようするに何度も何度も聴いた曲にはそういうところが出来るわけです。
「音」というよりかは「間」かな。音は音でもっといろいろ挙げられる。カンタベリー系のオルガンとかね。今書いてるのはそういうのとはちょっと違ったニュアンス。
2007年09月15日
■ V1から上丘へ行く経路はなにをしているか 補足
Retinal ganglion cellはLGNだけでなく、上丘へも投射しているわけですが、RGCの細胞の形態と発火特性による分類というのは非常に進んでいるので、すこしこれについて補足を。
まず、元ネタはThe Cognitive Neurosciences III (2004)の"Origins of Perception: Retinal Ganglion Cell Diversity and the Creation of Parallel Visual Pathways" Dennis Daceyより。ちなみにこのDennis DaceyというひとがLGNのkonio layerに入力するblue-on cellがsmall bistratified cellという細胞グループであることをを示したNature 1994のファーストです。
LGNおよび上丘にretrograde tracerを注入して、retrogradeにlabelされたRetinal ganglion cellの反応特性を見たあとでdendriteの走行などのmorphologyを見てやる、というような仕事があって、macaqueのRGCは形態から17通りとかに分類されていて、メジャーなのがparasol cell (magno系)とmidget cell (parvo系)とsmall bistratified cell (konio系)で、これらでRGCの全populationの約75%を占めてます。これらはみんなLGNにしか行ってない。
上丘へ行ってるのは、recursive cellというやつで、これはdirection selective。SGSのニューロンはmacaqueではあまりdirection selectiveなものはないので、おそらくこのrecursive cellというのはSGSの応答のsourceとは考えにくい。もちろん、SGSへの入力にconvergenceがあればdirection selectivityがなくなってもいいんだけど。それから、べつのpopulationでmoderate monostratified cellというのがあって、これはあまり生理学的にはcharacterizeされていない。
以上の、上丘へ投射している細胞は全RGCのpopulationのうち約9%です。とても少ないと考えた方がよいでしょう。また、foveaかperipheralかによってもちがうでしょうし、上丘への投射は視野のnasal側かtemporal側か(鼻側か耳側か)によってassymmetricになっていることが知られています。(Rafalのhumanの仕事はこれを論文のロジックの重要なところに使っています。)
あと、現在の文脈で重要なのは、Melanopsin-containing cellです。これはさいきん一挙に話題となってますが、ほかのRGCとちがって、自分でmelanopsinを持っているので光感受性になってるのです。ふつうのRGCはrodやconeが光を受容して、それをhorizontal cell, bipolar cell, amacrine cellなどで処理したあとの情報を受け取るだけなのです。それで、このMelanopsin-containing cellは視交叉上核(SCN、サーカディアンリズムを持ってる)を介して松果体に情報を送っていて、サーカディアンリズムとか、pupil reflexとかに関わっていると考えられています。
このMelanopsin-containing cellが自分のmelanopsinだけでなく、cone, rodからも入力を受けていて、L+M-SのシグナルをLGNに送っていることを示したのがDaceyのNature 2005 "Melanopsin-expressing ganglion cells in primate retina signal colour and irradiance and project to the LGN"です。この細胞は上丘へは投射していませんが、pretectumへは投射しているので、もしかしたらこれがextrageniculate pathwayでcolorの情報を担っているのかもしれません。
さて、おつぎの元ネタはAnnual Review of Neuroscience 2000 "The Koniocellular Pathway in Primate Vision" Stewart Hendry and Clay Reidです。上丘のSGSからLGNへの入力というのはLGNのkonio layerにのみ終止しているということが知られています。LGNのKoniocellularのlayerというのはmagno, parvoのlayerの隙間にあるわけですが、このうちいちばんventral側、つまりmagno layer側のkonio layerだけにSGSからの入力が来ています。
あと、"The ventral-most K layers, however, are innervated by types of γ and ε ganglion cells, similar in several ways to the types of cells that send their axons to the superior colliculus and pretectum."なんて書いてある。またややこしくなってきた。いったいどういう種類のRGCの入力が上丘に来てて、それがさらにLGN konio layerにどういう情報を送っているか、という問題ですね。上丘にkonio系の入力が来ているとは思えないのだけれど。
あと、こんなふうに言ってる。
"Zeki (1995) has proposed, however, that a direct geniculo-extrastriate path could account for the same phenomenon. That proposal is particularly attractive given the robust innervation of the foveal representation of V2 and the likelihood that this region has been little explored in studies of V1 ablation or cooling because of its proximity to V1. Thus, a relatively rich innervation of V2, with its feed-forward projections to most other extrastriate areas, as well as a much poorer direct geniculocortical innervation of those other areas, could well give the K layers a central role in blindsight."
Blindsightはfoveaだけによらないこと、それからnhp studyでは多くの場合V1 lesionによってV2もablationされていること、などからして賛成できませんが、residual visionを支えるpathwayがM, P, Kのどれか、という視点は大変重要であると思います。
今日はこのへんで。
- / ツイートする
- / 投稿日: 2007年09月15日
- / カテゴリー: [上丘、FEFと眼球運動]
- / Edit(管理者用)
2007年09月13日
■ V1から上丘へ行く経路はなにをしているか
今回もサーベイとJCを兼ねて。上丘への視覚入力にはretinaから直接来るretinotectal (retinocollicular)な経路と大脳視覚皮質を介してくる経路があります。V1からも来るし、V2からも来る。LGNからは来ないみたいだけど。Schillerの論文でJ Neurophysiol. 1974 Jan;37(1):181-94. "Response characteristics of single cells in the monkey superior colliculus following ablation or cooling of visual cortex."というのがあります。nhpのV1をcoolingもしくはlesionして急性で上丘の視覚応答を記録すると、SGSの応答はcoolingまたはlesionによって影響を受けないけれども、SGIの視覚応答は全くなくなる、というものです。つまり、SGSはretinaからの直接入力によってdriveされるけれども、SGIはV1経由の信号がないとvisual responseを引き起こすことができないのです。解剖学的にはV1のlayer5,6からSCへの投射があることが知られていますし、LIPやFEFを回ってくればいくらでも上丘へ行く経路はあります。ただ、麻酔下なんで、あんまり高次なところは回ってないんではないかと思うんですが。
ひきつづき彼らはJ Neurophysiol. 1976 Nov;39(6):1352-61. "Quantitative studies of single-cell properties in monkey striate cortex. IV. Corticotectal cells."にて、上丘(表面から0.5mmあたりと書いてあるので基本的にSGSからSoあたり)を刺激しながら、V1からunit記録をして、上丘の刺激によってantidromicに刺激されるunitを見つけます。こういうニューロンはV1のlayer 5,6にあって、antidromicの応答潜時は4.6ms。でもって、そういうニューロンの性質を調べると、complex cellで、orientation tuningはbroadで、direction selectivityがあって、spontaneous activityが高い。そういう意味ではSGSのニューロンの性質には似ていません。そこで彼らのdiscussionとしては、V1から上丘へ来る入力は「SGSの応答特性を作っている」というよりは、「SGS->SGIの情報の伝達をゲートしている」のではないか、と言っています。これは重要かつ面白い指摘です。
さて、このあとで彼らはJ Neurophysiol. 1979 Jul;42(4):1124-33. "Composition of geniculostriate input ot superior colliculus of the rhesus monkey."を発表します。これは上丘SGIからunit記録して、LGNのmagnolayerをlidocaineでblockすると応答が消えるけど、parvo layerをblockしても応答が消えない。どっちをblockしてもSGSの応答は消えない、という論文です。彦坂先生の仕事以降だとmuscimolになるのですが、この時代は局麻でblockしているのでpassing fiberもblockしてしまうという問題があります。ただ、SGSの応答は消えない、というのがcontrolで効いているので、だいたい妥当なのではないかと思うのですが。というわけで上丘はV1から受けている入力はmagno系だけだ、というわけです。色の情報とかはもらってない。
さて、この論文の意義を説明するためには視覚情報がRGC->LGN->V1->V2と進む情報伝達にはM,P,K経路の三つのチャネルがあるということの説明が必要です。図を作っておきました。
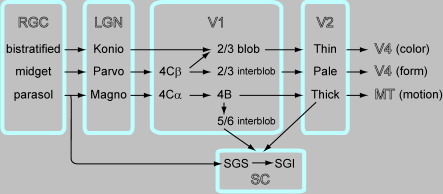
Schillerのだとcatの論文を引いて、X-type, Y-type, W-typeって話になるんでそこは翻訳しないといけないんですが、最新のレビューをもとにしてこれまでの知見を再構成してみました。図の元ネタはAnnu Rev Neurosci. 2005;28:303-26. "The circuitry of V1 and V2: integration of color, form, and motion." Sincich LC, Horton JC.および "The M, P, and K Pathways of the Primate Visual System" Ehud Kaplan in The Visual Neurosciences Ch.30です。図の表現上、一番上がkonio、真ん中がparvo、一番下をmagnoとしてあります。それぞれがcolor, form, motionのべつべつのチャネルとなってventral pathway, dorsal pathwayに行っていることはよく知られたことです。
それにさらに上丘への投射を組み込んでみました。LGNのparvocellular layerに終止しているRGCのX-type cell (midget cell)は上丘へは投射していません。LGNのmagnocellular layerに終止しているRGCのY-type cell (parasol cell)は上丘へ投射しています。あと、catではRGCにW-cellというのがあって、上丘に選択的に投射しているという話があります。(Klaus-Peter HoffmannによるBrain Res. 1972 Aug 25;43(2):610-6. "Very slow-conducting ganglion cells in the cat's retina: a major, new functional type?"およびJ Neurophysiol. 1973 May;36(3):409-24. "Conduction velocity in pathways from retina to superior colliculus in the cat: a correlation with receptive-field properties.") あと、conduction velocityが非常に遅い8m/secのはこのW-cellです。
ちょっと話が細かくなるのでここで改パラグラフ。CatのW cellがnhpになにに対応するかですが、koniocellularと考えた方がよいようです。Shapleyの80年代のTINSとかではそんな書き方をしています。そうすると、hnpではW-cellに対応するものは上丘へは投射していないと考えて方が良さそうです。ただ、Visual neurosciencesにもありましたが、W-cell、konioてのはどちらかというとmagno系、parvo系以外を全部入れたヘテロな集団と考えた方がよいので、これからの研究の進展によって話が変わってくる可能性はあります。なお、Schiller 1979では、W-like cellという言い方はintroでしているけど、それ以降ではまったく言及していません。また、それ以降の論文をしらべてみるとnhpでもW-like cellと表現しているのがあって、PNAS 2007 "Geniculocortical relay of blue-off signals in the primate visual system"とかではW-like = blue-off = koniocellularと書いてます。上丘にS coneからの入力が行っていないことはS cone isolating stimulusによる刺激による注意の効果の論文(Current biology)などから示唆されていますが、実験的には証明されていないようです。SGSからvisual response記録して、L-, M-, S-cone isolating stimulusで刺激するという実験をしたらいいと思います。きれいにS coneだけinsensitiveであることが示せたらJNSあたりいけるんではないでしょうか。
話を戻します。視覚皮質から上丘への入力はいくつかありますが、細かいことがわかっているのはV1,V2からのものです。
上丘にretrograde tracerを入れてV1で染まったニューロンの位置と、V1をCO染色してCOバンドとの位置とを比較したのがVis Neurosci. 1996 "The distribution of corticotectal projection neurons correlates with the interblob compartment in macaque striate cortex." Lia B, Olavarria JF.です。Layer 5/6のinterblobのニューロンが上丘へ投射しています。残念ながらinjectionは限局してないので、terminalはSGSなのかSGIなのかは不明。それ以前にもW. FriesがEBR 1985でlayer 5/6のMynert cellで上丘とMTの両方に投射しているものがあることを示しています(retrograde tracerによるdouble labellingを使用)。
同じグループが同様にしてV2で染まったニューロンの位置と、V2をCO染色してCOバンドとの位置とを比較したのがJ Comp Neurol. 1997 "Distribution of neurons projecting to the superior colliculus correlates with thick cytochrome oxidase stripes in macaque visual area V2." Abel PL, O'Brien BJ, Lia B, Olavarria JF.です。V2のthick band、つまりmagno系からおもに入力が上丘に来ています。
以上をまとめると図のようになるのでは、というわけです。
補足。ちなみに近年はganglion cellのレベルではなくて、coneのレベルでL+M-Sとかいったやり方で機能を描写するので昔の書き方を翻訳しないと使えません。このへんに関する最新の成果がEdward M. CallawayのNature 2003 "Parallel colour-opponent pathways to primary visual cortex"です。これを踏まえてconeからRGCまでの変換を書くと、parasol cellはcone全部の信号を足していて、L+M+Sで色に関わらない明るさをコードしていて、midget cellではL-Mでred/greenのシグナルを作っていて、bistratified cellではS-(L+M)でblue-onの受容野を持っています。そこでblue-offのニューロンを見つけてそれがV1のlayer 4Aに入力しているというのを見つけたのがNature 2003の仕事でした。これは図に組み込んでないけど、bistratifiedがblue-on cellでlayer 3Bあたりにメインで行ってて、それとはべつにRGCにblue-off cellがあって、これが4Aに終止していて、この二つを合わせてblue-on,offに関わるkoniocellular系を作っている、というのが最新バージョンではないでしょうか。
意外に充実してきたので今度のセミナーはこれにするかも。ちと専門的すぎるけど。
- / ツイートする
- / 投稿日: 2007年09月13日
- / カテゴリー: [上丘、FEFと眼球運動]
- / Edit(管理者用)
2007年09月12日
■ 神経科学大会終了
発表の方は無事終わりました。まあ、練習に時間をかけた分くらいは伝わったのではないでしょうか。伝えることをメインに、かなり削りました。トークで詰まってしまうことがないようになんとかセリフ全部記憶してゆっくりしゃべるよう心がけました。
いつもできないのが、節目で一息つくとか、観客全体を見渡すとか、緩急を付けること。ようするに、スクリーンばっかり見っぱなしにならないようにしたいんですけどね。やっぱSteve Jobsリスペクト。SFNのときこそうまくやるつもりです。ちなみにSFNのトークもsaccadeのセッションです。Visionのひとの方が面白がってくれそうなのだけど、データは上丘ニューロンの活動なもんで、苦渋の選択です。
あと、オチをうまく付けたいんですけどね。"Thank you for your awareness of my talk, thank you even if it is a random choice for you"とか言ったらクドいですかね。
あとでポスター会場で福島研に所属しているガイジンさんに、おまえのトークを見たけどジェスチャーが激しいので日本人じゃないのではと思った、と言われて苦笑い。
んで、ラーメン二郎 横浜関内店と吉村家本店に行ってきたという出張でした。ラーメン二郎のほうは、「汁無し」を頼みそびれたので(店に入る前に指定する必要があったのに気付かず、ふつうの小ぶたになった)、明日また行って頼む予定。ラーメン博物館の方はいいや。
明日はplexonの講習会。まわりのひとはかなり導入しているようですけど、わたしはまったくわかってないんで基礎的なところからです。
2007年09月08日
■ VOCALOID 2
ニコ動とか見ましたけど、スゲーっすね! まさに日本ハジマタ。
詳細はIT mediaあたりへどうぞ。
CRYPTONってドラム音源のEZ drummerとか扱ってるところではないですか。もしかしたらVSTとしても使えるかも、そしたらSONARとFL studio持ってるんで同期できるんだけど、と思って調べてみると、できる! ただ、製品サポートのところに、SONAR, FL studioではVSTとしては使えない、なんて書いてある。SONARではrewireとしては使えるそうなので大丈夫そうだけど。
ドラム音源買おうとか思ってaddicitive drumのデモ版インストールしたりしてるところなんだけど、ドラム音源買う前にこっち買いそうってのもどうかと思うんですが、これは衝撃。第一代目VOCALOIDとデモソングを比べればレベルの上昇は歴然。日本ハジマタ(<-もういい)。もうこれで自分で歌わなくても歌詞入れできる! もうこれでギター入れるの以外は全部PC上で出来る! 打ち込みっぱなしではなくていろいろ調整必要そうだけど、それは他の音源だって同じですしね。
声は萌え系だからサイケデリックロックには合わないけどね!って書いて締めようかと思ったのだけど、じつはシューゲイザーでは合うかも。My bloody valentineの"Lose My Breath"がわたしの人生の一曲のひとつですが、これなんかきっと合うよ。
iPod touchも買いたいし。ちょっと物欲出過ぎてます。メシ代削るか。
神経科学者としてはこのような自然な日本語の発声を可能としているYAMAHAの「周波数ドメイン歌唱アーティキュレーション接続法(Frequency-domain Singing Articulation Splicing and Shaping)」に興味を持つべきなんでしょうけど、ググってもあまり資料が出てこないんで、とるものもとりあえず(なんか使い方へん)。
2007年09月05日
■ 上丘の細胞構築
もうすぐ神経科学大会ですね。私は一日目の午後に口演です。眼球運動のセッションの一番最後で、視覚1の直前。ちょうどいい位置に入れてもらいました。ぜひ見に来てください。
毎年7月末にあるトレーニングコースではラットの上丘、視覚皮質、バレル皮質などから急性で記録をしてあとでhistologyで記録部位を同定するということをやっております。わたしはだいたい電気生理の解析とhistologyの解析の担当をこの3年くらいやってます。上丘の細胞構築(cytoarchitecture)をきっちり把握しておく必要がありますので、元ネタにあたっておいたものをまとめておきます。
まず、上丘とはなんぞや、から始めてると時間がありませんが、上丘は中脳にあります。つまり、視覚皮質よりも腹側(ventral)で、視床よりも尾側(caudal)です。上丘は大きく分けて、retinaからoptic nerveを介して視覚入力が入ってくる入力層と、脳幹のサッケードジェネレーターに眼球運動の指令を送る出力層とに分かれます。おおまかには入力するoptic nerveが入ってくるSOと、メインの入力層であるSGSと、メインの出力層であるSGIとがわかれば充分です。
教科書的な図はありませんが、PNAS 2007 "Identity of a pathway for saccadic suppression"がopen accessなので、これの図を見てもらえると良いのではないでしょうか。Fig.3を見るとSGS,SO,SGIの順に並んでいることがわかるかと思います。さらにFig.2をみてもらうと上丘はさらに6層に分かれることがわかります。いちばん表面にある薄い層であるSZを入れると全部で7層構造です。ちょっと大脳皮質とは細胞構築が違う。ただ、SO=4層、SGIの大きい出力ニューロン=5層のpyramidal neuronくらいに当てはめてイメージすることはできるかもしれません。
解剖学の論文ってのは細胞構築、つまりニッスル染色でみた、細胞(主にニューロン)の大きさや密度などが層ごとにどのようなパターンを示しているか、の記述というのがずらずら並んでたりするので、それを読んで顕微鏡で標本を見てどこがどの層なのか「目を慣らす」という作業が必要となってきます。
Ratの上丘でのこのような細胞構築の分類の元となった論文は以下のJCN 1981です。これはさらにcatでの細胞構築JCN 1974 Kanaseki and Spragueを下敷きにしています。
J Comp Neurol. 1981 Sep 10;201(2):221-42. "Trigeminal projections to the superior colliculus of the rat." Killackey HP, Erzurumlu RS. PMID: 7287928
論文から抜き書きで7層についての記述を。
-
SZ: the stratum zonale
"a thin, loosely patched layer, which contains small neurons" -
SGS: the stratum griseum superficiale
"densely packed with cellular elements and is relatively thick" -
SO: the stratum opticum
"primary composed of fibrous elements with some scattered cells" -
SGI: the stratum griseum intermidiale
"band of large neurons readily detectable in Nissl preparations"
"densely packed with cells and is thick, like the stratum griseum superficiale" -
SAI: the stratum album intermediale
"characterized by patched of fiber bundles intermixed with a few cellular elements" -
SGP: the stratum griseum profundum
"has a layer of large neurons at its dorsal border" -
SAP: the stratum album profundum
"a fibrous layer that separates the superior colliculus from the periaqueducal dray"
細胞構築を決めてやるために必要なことは基本的にはこのような記述がすべてなのです。もちろん、immunohistochemistryとかでどこかの層がなんか特異なマーカー(CaMKIIだとかIh channelだとかcalcium binding proteinだとか)を持っているかを目印にしたりとか、トレーサーを注入して、投射関係などからさらにくわしく調べられ、consistencyの確認や、さらなる層の細分化が起こるわけです。たとえばの例として、LGNがmagnocellularとparvocellularに二分されていて、層のあいだの隙間はinterlaminarなんて言われていたところをCaMKII α-unitで染めることによってkoniocellularの層が確立する、ということもありました(Science 1994 Hendry and Yoshioka)。Koniocellularという言葉自体はその前からあったんですが。
ちなみに、ネットで探してみたらヒトでの上丘の細胞構築については「中脳蓋-上丘を主として-」を見つけました。
- / ツイートする
- / 投稿日: 2007年09月05日
- / カテゴリー: [上丘、FEFと眼球運動]
- / Edit(管理者用)
2007年09月02日
■ カポーティとかグラースサーガとかブローティガンとか読んでます
ども。元気です。論文コメントは進まず。
「ロング・グッドバイ」とか「グレート・ギャッツビー」の村上春樹訳とか読んだあたりからアメリカ文学にひさびさに回帰中です。あ、作品について言及するんで、ネタバレ有り、と書いておきます。
トルーマン・カポーティはオザケンつながりで(「犬は吠える」ね)、大学生の頃に原書まで買って気合い入れて"A Christmas Memory"とか読んだりして、いいかんじだったんだけどそれに入ってた"Breakfast at Tiffany's"にはたどり着けず放置。で、ついにあきらめてこのお盆に訳書で読みましたが、よかったです。ストーリー的には、わたしバブル世代なもんで恥ずかしながら、つい東京ラブストーリーのリカ(原作コミックの方ね。ドラマは知りません)とか想起してしまいましたけどね(いや順番逆だから)。訳が古くさいんで新訳が出るといいんではないかと思いましたけど、さっそく原書へ。もう50年前のものだから元の文章も古くさいんだろうけど("phony"とかね、Frank Zappaの歌詞とかでしか聞かないような)、表現ひとつひとつが読んでて楽しいです。つぎは「冷血」へ行けば映画の「カポーティ」が楽しめそうなのだけれど、私が好きそうなのは初期の方(まさにオザケンってかんじのこういう写真の時代ね)かなということで、「遠い声、遠い部屋」を選びました。まだ読んでるのはじめの方だけど、素晴らしいです。車のガラスのひび割れを「星が破裂したみたいな」みたいに表現したりとか。南部を舞台にしたものは前にフォークナーの短編とか読んで気が重くなっちゃったので向かないかなと思ったけど、意外にいけそう。
あと、佐藤友哉つながりでサリンジャーのグラースサーガを。「ナイン・ストーリーズ」は読んだはずだけど、もう忘れた。「フラニーとゾーイー」から。「フラニー」いいかんじ。こう、かみあわないかんじが良く書かれてます。解説書とか読んでると、レーン(Lane)を落としてフラニーを上げるような書き方をしているけど、フラニーの中にもレーン的な要素はあるし、ことさら貶めるようには書いてないと思うけどなあ。そもそもそんないい者悪者みたいな読みは「だれもがふとっちょおばさん("FAT LADY")だ」という啓示を踏まえていないと思うんだけど。
「ゾーイー」のほうはバディのもったい付けまくった文体にうんざりして断念しかけたけど、最終的には良かったです。わたしの読みが正しいかどうかわからないのだけれど、あの「FAT LADAYとは**(いちおう消しときます)のことなんだよ」というゾーイーの発言は本人も知らなかった、その場でひねり出した(シーモアとバディの力を借りて)、言葉だったんではないでしょうか。あらかじめ知ってたキメ言葉なんかじゃなくて。なんかわたし自身の人生の大事なタイミングでどう行動したかとかを反芻しながら考えるに、そういうものだった気がするんです。ゾーイーは言ったあとには「決まった!」とか思ったかもしれないけど、それをひねり出してくる過程はそれまでの、饒舌でなにもかもをダメにしてしまうところからほんのひととき離れることに成功していたのではないか、って思うんです。あー語った語った。
というわけでかなり楽しめたので、勢いづいて「大工よ、屋根の梁を高く上げよ シーモア-序章-」も購入。「ナイン・ストーリーズ」も原書で"A Perfect Day for Bananafish"とか読んでみました。でも、最後の方の"Hey!" "Hey, yourself"とか原文で読んでもニュアンスがわからんのでけっきょく訳書を頼ることに。英語英語。
図書館で「サリンジャー―伝説の半生、謎の隠遁生活」中公新書を読んだけど、なにこの上から目線。サリンジャー好きでないなら書かなけりゃいいのに。かなりがっかり。
というわけで、だれか本田透方式(バッファロー'66や宮沢賢治に非モテ主人公を当てはめて読む)でグラースサーガを捉え直すべきですよ。もしくはBananafishのSybilの会話を麻枝/久弥風にアレンジとか("Hey"=「わっ、なにするんだよっ」みたいな)。すでに「フリッカー式」でやってるってオチは無しで。……ぶっこわれてきたことを認めます。
あと、ブローティガンね。これまた大学生の頃に「ロンメル進軍」(高橋源一郎訳)とか「西瓜糖の日々」とか読んですごくよかったんで、原書で"Trout Fishing in America"と"In Watermelon Sugar"の合冊っつうのを買って持ってたんだけど、これも積んだままで。んで訳書で「アメリカの鱒釣り」がついに文庫化したのでさっそく読んでみたけど、さっぱり読めない。断念。なんか、こういうアメリカ風ユーモアみたいなのはダメみたい。もしくはこういうのが読めなくなっただけか、と「西瓜糖の日々」を再読してみたところ、こっちはイケる。かなりすばらしい。この独特の世界、すごく静かなのに常に不安に脅かされている。どこか現世ではないかんじ(近未来というよりはこの世でないかんじ)、にすごく惹かれたのだとわかりました。ようするに「えいえんのせかい」ですよ。ってだから俺が本田透式読みをしなくていいっつうの。
そんなかんじでお盆休みと札幌で本を読んでました。
{kind=link}
お勧めエントリ
- 細胞外電極はなにを見ているか(1) 20080727 (2) リニューアル版 20081107
- 総説 長期記憶の脳内メカニズム 20100909
- 駒場講義2013 「意識の科学的研究 - 盲視を起点に」20130626
- 駒場講義2012レジメ 意識と注意の脳内メカニズム(1) 注意 20121010 (2) 意識 20121011
- 視覚、注意、言語で3*2の背側、腹側経路説 20140119
- 脳科学辞典の項目書いた 「盲視」 20130407
- 脳科学辞典の項目書いた 「気づき」 20130228
- 脳科学辞典の項目書いた 「サリエンシー」 20121224
- 脳科学辞典の項目書いた 「マイクロサッケード」 20121227
- 盲視でおこる「なにかあるかんじ」 20110126
- DKL色空間についてまとめ 20090113
- 科学基礎論学会 秋の研究例会 ワークショップ「意識の神経科学と神経現象学」レジメ 20131102
- ギャラガー&ザハヴィ『現象学的な心』合評会レジメ 20130628
- Marrのrepresentationとprocessをベイトソン流に解釈する (1) 20100317 (2) 20100317
- 半側空間無視と同名半盲とは区別できるか?(1) 20080220 (2) 半側空間無視の原因部位は? 20080221
- MarrのVisionの最初と最後だけを読む 20071213