[月別過去ログ] 2008年07月
« 2008年06月 | 最新のページに戻る | 2008年08月 »2008年07月31日
■ 生理研研究会 オンライン参加申し込みを開始しました
生理研研究会 「認知神経科学の先端」ですが、今年は玉川大学の松元健二さんとともに「動機づけと社会性の脳内メカニズム」というタイトルで行います。日程は2008年9月11-12日。以前アナウンスしたエントリは20080609。
そこからなかなか進んでいませんでしたが、研究会のサイトをアップデートしてオンライン参加申し込みを開始しました。参加申し込みフォームのページ。参加申し込み要領をよくお読みになってからお申し込みください。多くの方の参加をお待ちしております。
6月の視覚研究会のときに配ったビラに書いた講演者紹介をコピペしておきます。(一部松元さんに増補していただいております。) それではここから:
講演者の方の紹介など。
さて、ここからはもっとくだけたかんじで講演者の方の紹介を書いてみます。Pooneilブログ出張版?とでもいいましょうか。
大まかに分けると「動機づけ」パートが南本さん、出馬さん、村山さんで、「社会性」パートが細川さん、守口さん、遠藤さんです。それぞれのパートに動物実験、ヒトでの実験、社会心理の方に入っていただきました。動機付け、社会性、ともにどうやって実験の形に持ち込めばいいか、ということがなによりも難しいところだと思います。そこで、実験を通してアプローチしている先生方に加えて、社会心理の先生方にトークをしていただくことで動機付け、社会性という概念の広さを踏まえた議論ができるようにと配慮しました。
南本さんは京都府立大の木村先生のところで視床CM核の仕事("Complementary process to response bias in the centromedian nucleus of the thalamus." Science 308: 1798-1801, 2005)を出版されました。そのあとNIMHのBarry Richmondのところに留学されて、rewardのスケジュールと行動の関係に関する仕事をされて現在は帰国し、放医研に在籍しています。放医研にだんだんシステム関係の人が集まってきてますね。今回の講演ではおそらくNIMHでのお話をしていただけるのではないでしょうか。まだ未出版かと思いますが、SFNのアブストと説明が木村研ラボのサイトで読めます。このあいだRichmondが生理研に来たときのトークで一部を聞かせていただきました。要は課題を行ってからrewardが出るまでの時間を振っておいて、それをcueで予測できるようにしておくと課題の成績がcueによって影響を受けるようになる、というものです。これまでの設楽さんや菅瀬さんが行ってきた一連の研究と併せて、動機づけの研究としてどう捉えたらよいか、というあたりが論点のひとつとなるのではないでしょうか。
出馬さんは生理学研究所の心理生理学、定藤先生のところに所属している博士課程3年生の方です。昨年の生理研研究会でポスター発表をしてくださった研究が今年の4月にNeuronに掲載されました("Processing of Social and Monetary Rewards in the Human Striatum" Neuron, Vol 58, 284-294, 24 April 2008)。他人に褒められたときに活動する脳部位と、お金を報酬としてもらったときに活動する部分とが重なっていた(ともに線条体に activationがあった)、というものです。報酬と動機づけに関わるだけでなく、社会性に関しても関連のある仕事であると思います。
村山さんは東大教育心理を出られて東工大に在籍してからRochester大学に留学されてAndrew Elliotの研究室に在籍していました。Andrew Elliotは達成動機づけ理論にかんする第一人者です。動機づけには報酬と絡めた意思決定のような外発的動機づけだけではなくて、内発的動機どけというものがあります。そういう人間らしい複雑な部分をどうやって捉えて脳研究に結びつけたらいいのか、ということについてコメントしていただけるかと思います。また、村山航さんのホームページは心理統計などに関する有用な資料があります。わたしは知らず知らずのうちにお世話になってました。
細川さんは霊長研出身で現在は神経研の渡邊正孝先生のところに所属していらっしゃいます。昨年の神経科学大会のシンポジウムでも発表されていた対戦ゲーム中のサル前頭連合野ニューロン活動についてのお話をしていただけるのではないかと思います。この研究はサルどうしの対戦という実験系からして社会性に関する研究としてわたしは理解していたのですが、かならずしもそれだけではなく、動機づけの研究という側面も持っているようです。つまり、相手と競うという条件であることが動機づけに影響を及ぼすのではないか、rewardそのものだけではなくて相手に勝つということが動機づけとなるのではないか、というわけです。ということで本研究会の両テーマに関わるトークとなるのではないかと思います。
守口さんは国立精神・神経センターに所属されていて、アレキシサイミア(失感情症)の患者さんについて研究をされています。アレキシサイミアの患者さんは自己の情動の同定・表象が困難な症例ですが、これを「心の理論」の障害と捉えて機能イメージングを行った論文が出版されました("Impaired self-awareness and theory of mind: An fMRI study of mentalizing in alexithymia" NeuroImage 32 (2006) 1472-1482)。これで活動するのもやはりmedial prefrontal cortex。東北大学グローバルCOE若手フォーラムでのトークについての報告がwebにありますが、これをみると、さまざまなコンテクストを絡めたお話をしていただけるのではないかと期待しております。
遠藤さんは東大教育心理の准教授をされています。専門はアタッチメント、つまり発達時に母子が親密であることがどのように重要なのかに関する研究ですが、これは社会性の基礎となる概念であります。また、アタッチメントの概念自体はヒトだけでなく、もともと動物の行動観察から見出されたものであります(いま wikipediaにそう書いてあるのを見つけた)。「心の理論」などと併せて、社会性に関する大きなパースペクティブからコメントをいただけるのではないかと思います。
以上の6人の先生方で研究会を行います。指定討論者の先生方もまたあらためてお願いしますので、依頼された方はぜひ引き受けてガンガン質問、議論してください。今回も途中質問有りにしておきたいと思いますが、総合討論の場を作ってもっとgeneralな議論ができるようにします。でも、いわゆる学会のパネルディスカッションみたいにすると盛り上がらない。たんに補足質問タイムになってしまう。だから、なんか煽り気味のネタを用意したり、あらかじめ募集しておいたりしたらどうかと考えています。たとえば、「動機づけって概念はほんとうに必要か? すべては報酬による意思決定ではないのか」とか。ふつうだったら紛糾して時間切れになるようなネタを30分かけて議論できたらいいんではないかと思います。このへんの形式についてはまた計画ができたら発表したいと思います。
それでは、みなさまぜひお越しください。
- / ツイートする
- / 投稿日: 2008年07月31日
- / カテゴリー: [生理研研究会2008「動機づけと社会性の脳内メカニズム」]
- / Edit(管理者用)
2008年07月27日
■ 細胞外電極はなにを見ているか
夏が来れば思い出す・・・のは生理研トレーニングコースの準備です。ここ数年は当研究室では、「麻酔下動物での急性電気生理実験」というタイトルで、細胞外電極を使って麻酔下ratからLFPを記録して、視覚刺激に対する上丘の応答をみたり、ひげ刺激に対するbarrel cortexの応答を見るということをやっております。でもって、LFPで記録しているものがなにか、ということを毎年わたしがちょこっと講義しています。
問題なのは、適切な教科書がないということです。この問題については以前にも書きました(20050817および20050818)。細胞内電極やパッチクランプ用電極を使った細胞内電位の記録に関しては、ホジキン・ハックスレーあたりについて書いている生理学の教科書を読むことから始めて、"the axon guide"あたりまで読んでおけばだいたい入門できるかと思うのですが、細胞外電極を使った細胞外電位の記録に関しては良いものがありません。わたしが日々行っている、行動中のnhpのsingle-unit recordingがまさに、タングステン製の細胞外電極を使って、単一細胞のaction potentialを細胞外電位から記録しているものなのですから、よろしくない事態です。また、multiunit recordingやLFPなどが今後BMIなどの応用の面からさらに重要になってくるであろうことを考えると、このへんの基礎をきっちり押さえておくことは大切なことだと思います。
それで、今年もトレーニングコースの時期がやってきましたので、いそいそと準備しているのですが、しょうがないので自分でいろいろ調べて作ってみました。まだ自分で納得いけてないところがあるのでここで皆様の意見を聞きたい、というわけです。物理をわかっている方ぜひご協力お願いします。それではここから:
追記081108:リニューアル版のエントリを作りました。そちらもご覧ください。
(印刷用にPDFファイルを作りました。ファイルをダウンロード。数式のフォントがきれいでないのでhtmlを印刷した方がよいかも。)
(追記:いろいろ書き直してます。オリジナルのエントリはpdfファイルとして保管しておきます。)
[まとめ]
- 細胞外電極はなにを計測している? - 細胞膜を流れる電流。
- 細胞膜の間を流れる電流の正体は? - シナプス電流とaction potential。
- 低周波成分はLFPと呼ばれ、シナプス電流を反映している。
- 高周波成分はMUAと呼ばれ、action potentialを反映している。
- LFPは必ずしも電極の近傍で起こっている電流を反映しているとは限らない。離れたところで起こっている電流を反映している可能性がある。
- CSD解析では、さまざまな深さから記録した細胞外電位から、細胞膜の間を流れる電流の発生位置を推定する。
- 細胞外電極が神経細胞の非常に近傍にあるとき(<50µm)には、単一の神経細胞からの活動電位を反映したsingle-unitを記録することができる。
[細胞外電極はなにを計測している?]
ひとことで言えば、電極近傍の細胞膜を流れる電流を計測している。細胞膜電位ではない点に注意。
もっと正確に言うために、細胞内電極での記録と比較してみよう。図1(1)にあるように、細胞内電極は細胞内の電位Vintと無限遠に置いた不関電極での電位V∞との差を計測している。これはけっきょくのところ、細胞膜間の電位差(細胞膜電位)を計測していることになる。
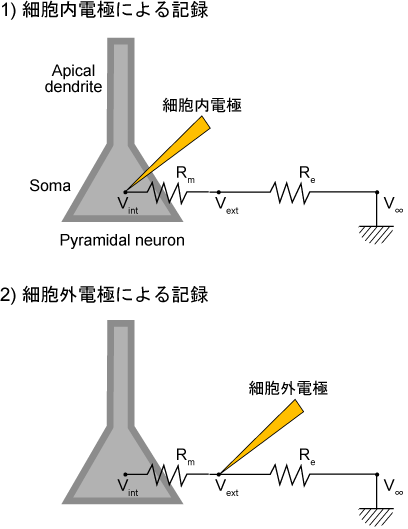
図1 細胞内電極(1)と細胞外電極(2)の記録の違い
Vint - V∞ ~= Vint - Vext (Rm >> Rextより、) = Vm ---(1)
細胞膜の両端にかかっている電位を計測しているという意味で、これはテスターである部分の電位差を測るのとまったく同じである。
一方で、細胞外電極(図1(2))は細胞間隙に置いた細胞外電極での電位Vextと無限遠に置いた不関電極V∞とのあいだの電位差(細胞外電位)を測定している。
Vext - V∞ ---(2)
しかし、ここで測定されたReの両端の電位差そのものには生理学的な意味がない。つまり、テスターでの計測をイメージするとおかしなことになる。
じつは、細胞外電極で記録される電位差は、細胞間隙に置いた細胞外電極の周りで起こった電流によって出来た電場を反映しているのだ。
そして、重ね合わせの原理から、細胞外電位は近傍で細胞膜の間を流れる電流量による電位変化の足し算である。電流の発生源から電極までの距離に反比例して弱くなる。数式で書くと、
Vext = R/(4*pi)*Σ(Ij / rj) ---(3)
ただし、Vextは細胞外電極で測定された電位、Rは細胞間隙の抵抗でだいたい200-300ohm/cm。Ijはある場所jで流れた電流の大きさ、rjは場所jから細胞外電極までの距離。Σはすべての電流源jでの総和。(Nunez and Srinivasan(2006)より)
だから、無限遠に置いた不関電極では、r=∞でVext = V∞ = 0となり、電流の影響が全くないところでの電位を計測していることになる。不関電極との差分を計測することで細胞外電極は電流の影響を計測していると言える。
上記の(3)式についてもうすこしくわしく書くと、クーロンの定理から
E(r) = R/(4*pi)*I/r2 ---(4)電流Iがあると、r離れた部分に電場E(r)ができる。後述の通り、電場E(r)はベクトル場。
電場ができるとそこには電位が生まれる。つまり、
E(x,y,z) = -grad(V(x,y,z)) ---(5)ある地点(x,y,z)の細胞外電位V(スカラー場)の勾配gradientを取るとその地点の電場E(ベクトル場)になる、というふうに電位が定義される。
以上の(4)(5)からEとqを消去して、sourceごとに足し併せると(3)式になる。
ここでつかうべき(4)の式はcharge sourceの式ではなくてcurrent sourceの式であるべきだからこれで良いと思うのだけれど、Nunez and Srinivasan(2006)には記載がない...
[細胞膜の間を流れる電流の正体は?]
では、「細胞膜の間を流れる電流」とはいったいなにか。主に二つのものが考えられる。
- シナプス電流(EPSCとIPSC)。
- Action potential(に伴ったNa,Ca流入、K流出)。
これらはべつべつの周波数帯域と空間解像度を持っている。よって同じ電極からフィルタを使って両方の活動を分離することができる。
Local field potential (LFP):
- 周波数成分 1Hz-300Hzあたり
- 主に電極近傍のsynaptic current (EPSC, IPSC)による。
- synaptic currentはamplitudeは小さいが、time courseが長いので、近くの入力のspatial summationが起こる。
- 電極先端から0.5-3mm程度の範囲のシナプス電流が寄与する。(Logothetis (2008)より孫引き。) 電極の抵抗に依存する。
- あくまでシナプスの位置が重要であり、電極先端に細胞体があることを保証しない。このへんについては「Volume conduction、sinkとsource」にて後述。
Multi unit activity (MUA):
- 周波数成分 300Hz-5KHzあたり
- 電極近傍のaction potential による。
- action potentialはamplitudeは大きいが、time courseが短いので、近くの入力と<1msオーダーで同期しない限り、キャンセルアウトする。
- 電極先端から50-350µm程度の範囲のニューロンの活動電位が寄与する。(Logothetis (2008)より孫引き。) 電極の抵抗に依存する。
つまり、ざっくり言えば、一つの細胞外電極から記録される信号を周波数で分けることによって、LFPは入力(=シナプス電流)、MUAは出力(=action potential)、と違ったものを分けて見ることができる。
(なお、さいきんでは、action potentialのあとで見られるafterhyperpolarizationなどの遅いnon-synapticな成分がLFPに効いていることがわかってきている。けれども、それを入門のときに言うと混乱するので言わない。)
[Volume conduction、sinkとsource]
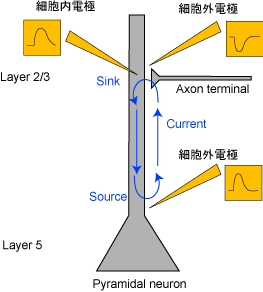
図2 興奮性シナプス電流
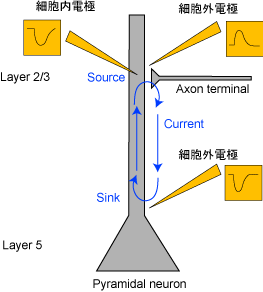
図3 抑制性シナプス電流
上の段で「細胞膜の間を流れる電流」には二つある、と書いたがそれは正確でない。上記のようなシナプス電流やaction potentialなどでactiveな電流が出来たときには、閉回路を作るためにpassiveな電流も流れる。よって、電極から記録されている現象が電極の近傍で起こっているactiveな電流によるものなのか、それとも離れたところで起こっている電流から作られたpassiveな電流によるものなのかは自明ではない。
たとえば、図2のようにシナプスで興奮性シナプス電流(EPSC)が起こっているとき、dendriteおよびsomaで細胞内電位の上昇が見られるが、細胞外電位はdendriteでは近傍の電流流入を反映した下向きのpotential(sink)が記録されるが、somaではpassiveな電流流出を反映した上向きのpotential(source)が記録される。よって、電極からsourceが記録されたとしても、かならずしも近傍の抑制性シナプス電流(IPSC)を反映しているわけではない。これは図3のように抑制性シナプス電流(IPSC)が起こっているときの図と併せて考えてみればわかる。つまり、下向きの細胞外電位が記録できたとしても、それは近傍での興奮性シナプス電流(EPSC)によるものかもしれないし、離れたところでの抑制性シナプス電流(IPSC)によるものかもしれない。
ともあれ、action potentialやシナプス電流によるイオンの流入流出よるactiveな電流にはpassiveな電流が伴う。これによってsinkとsourceのペアが生まれる。
このsink(吸い込み)とsource(湧き出し)という概念は、電磁気学の教科書で大昔たぶん習った、ラプラスの方程式、ポアソンの方程式で言ってることを数式を使わずに表現したもの。数式を使うと、電極の先端では電荷がないので、
div(grad(Vext)) = 0つまりラプラスの方程式となるが、この方程式を二つの境界条件、つまり、シナプスなどの電流源では
div(grad(Vext)) = -ρ/εポアソンの方程式が成り立つし、無限遠では
Vext = 0が成り立つ、というのを使って解くことで、Vextを計算することができる。
[Current source density (CSD)解析 その1]
上記の(3)の式にあったように、細胞外電極から記録される電位は電極近傍の電流の重み付き加算をしている。よって、一ヶ所の細胞外電位の記録だけではどこで電流流入が起こっているかを正確に推定することはできない。そこでさまざまな深さから細胞外電位を記録したデータをもとにして、電流の発生位置、つまりsinkとsourceの分布を推定する。これがcurrent source density (CSD) analysisだ。
結論だけ書くと、大脳皮質のように層構造を持っている脳部位で、深さ方向z(i) (i=1...n)で記録した細胞外電位をVext(z(i))とすると、それぞれの深さでの電流Im(z(i))は、
Im(z(i)) = -(1/R) * d2Vext(z(i))/dz2
となる(Mitzdorf 1985より)。つまり、深さz方向の軸で2階微分を計算すればよい。ただし、Rは細胞間隙の抵抗で、だいたい200-300ohm/cm。
深さ方向で等間隔に記録をしておけば、2階微分はある深さz(i)での細胞外電位Vext(z(i))とその隣り合った深さの細胞外電位Vext(z(i-1))およびVext(z(i+1))とから計算できる。
Im(z(i)) = -(1/R) * ( Vext(z(i+1)) + Vext(z(i-1)) - 2 * Vext(z(i)) )
よって、等間隔ということが非常にcriticalなので、一本の電極を進めてゆくよりかは等間隔で記録部分が並んでいるミシガン・プローブのタイプの電極を使って記録する方がよい。
くわしく導出仮定を書くと、
連続方程式より、 div(J) = Im ---(1)オームの法則より、
J = (1/R) * E ---(2)(ここでは1/R = σは均一の定数とする。)
前述の電場と電位の関係より、
E = -grad(Vext) ---(3)(2)と(3)よりEを消去すると、
J = -(1/R) * grad(Vext) ---(4)(1)と(4)からJを消去するとポアソンの方程式ができる。
(1/R) * div(grad(Vext)) = -Im ---(5)これをx,y,z成分にばらすと、
(1/R) * (∂2Vext/∂x2 + ∂2Vext/∂y2 + ∂2Vext/∂z2) = -Im ---(6)大脳皮質のように、深さz方向に延びたニューロンがx,y方向で同じように並んだ構造をしているときにはx方向、y方向の2階微分は0になるから、z成分だけが残る。
(1/R) * ∂2Vext/∂z2 = -Im ---(7)(Mitzdorf (1985)より。)
[Current source density (CSD)解析 その2]
CSD解析で得られたImから、ある時間tでの深さ方向のsink-sourceの分布を推定することができる。しかし、上記の通り、このようなsink-sourceのペアは[activeな電流(channel開閉によるイオンの移動)によるもの]と[閉回路を作成するためのpassiveな電流]のどちらに対応するかを一意に決めることはできない。そこで、解剖学から得られた細胞構築のパターンなどを援用することによって、activeな電流の位置を推定する必要がある。
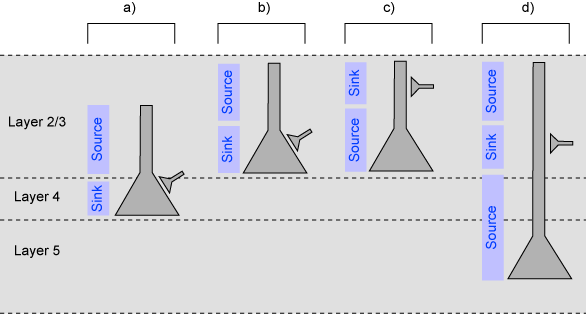
図4 CSD解析
かなり単純化した話だけれども、大脳皮質のおもなpyramidal neuronはlayer 3 (図4b-c)かlayer 5(図4d)にあって、apical dendriteを上に延ばしている。大脳皮質への入力はlayer 4 (図4a)にsomaのあるgranule cellに入力する。また、大脳皮質ではIPSCはあまりfield potentialに寄与していないことがわかっているので、EPSCだけに限る。(海馬とかだとこうはいかない。) また、sink-sourceの分布は細胞体を超えて広がることはない。
これらのことから、a)-d)のそれぞれ左にあるsink-sourceの分布パターンが見られたときには、a)-d)のそれぞれ右にあるようなニューロンへの興奮性シナプス入力が寄与していると推定することができる。
おわかりの通り、かなりざっくりとした推定なので、他の情報、たとえばsliceでの細胞内記録とそのあとのmorphologyの再構成とかと組み合わせていかないといろんなところに落とし穴がある。
このようなテクニックは1970年代後半から実用化した、けっこう古いテクニックではあるけれど、今後のLFPやMUAを使ったBMI研究においてinverse problemを解く際に、生理学的知見を用いたforward modelを作成するにあたって再び重要なテクニックとなるんではないだろうか、というのが私見。
[Single-unitの記録]
もし細胞外電極が神経細胞の非常に近傍にあるとき(<50µm)には、単一のニューロンからの活動電位を反映したsingle-unitを記録することができる。上記のMUAの特殊な例と捉えることができる。私自身はこっちが本業だけど。
教科書とかで見る細胞内電極で記録した細胞膜電位Vmはposiの山が一つで、ゆっくり落ちてくる。いっぽうで、single unitの波形Vextはそれより速くて、だいたいtriphasic。posi-nega-posi。このへんにはHenze et.al.(2000)とかの図を入れておきたい。
Single unitの波形Vextがなんであんな形をしているか。細胞内電極で記録した細胞膜電位Vmと関係づけたい。Henze et.al.(2000)では、
Vext = dVm/dt
時間ドメインでの1階微分としている。ただし、波形の後ろの方はafterhyperpolarizationとかそのへんの影響で、計算からかなりずれる。
[細胞膜電位と細胞外電位との関係の導出] (未完結)
これを導出するためには以下の手順が必要となる。(Gold et.al. (2006)より。) 複雑なのでだいたいNEURON simulatorとかが使われる。なんかもっと簡単な説明があると良いのだけれど。
- まず、Na channel、K channelの開閉によるチャネルコンダクタンスgNa, gKのダイナミクスとそれによる電流INa, IKを計算する。
- この電流によって起こる細胞膜電位Vmの変化をRallのcable方程式から計算する。
- この電流によって起こる細胞外電位Vextの変化をHolt-KochのLSA(line source approximation)によって近似的に計算する。
- 以上を(近似でいいから)解析的に解けばたぶん1階微分であることが示せるはず。
ただ、よくわからないのは、昔の本(Geddes(1972)とか)を読んでると、Lorente de No(1947)とかTasaki (1959)を引いて、2階微分だって書いてあるのがある。Axonでの記録だけど。(正確にはImとVmの関係についてだけど。) Llinas R, Nicholson C (1974)とかだと、Rallのcore conductor modelで近似できるような条件では、
the field potentials in the vicinity of extracellular sources or sinks are proportional to the first derivative of transmembrane voltage with respect to time.
とか書いてある。いっぽうで、excitable membraneでは、Imにはchannelによるcapacitanceの成分が入ってくるので、
the extracellular potential will be proportional to the second derivative of potential with respect to time
とか書いてある。たぶんこのへんを読めばわかるのかな。Lorente de Noまではさかのぼらずに済ませたいのだけれど。
そっか、基本的にはImとVmの関係さえわかればいいのだから、これは細胞外電極に限った話ではないな。Rallのcable方程式で、興奮性シナプスのある場合のバージョンを用いて、ImとVmとの関係の式にしてやればよいのだな。このへんを進めてみることにしよう。
[参考文献]
- Bedard C et.al., (2004) "Modeling extracellular field potentials and the frequency-filtering properties of extracellular space."(pdf) Biophys J. 2004 Mar;86(3):1829-42.
- G. Buzsaki (2006) Rhythms of the Brain: Oxford University Press (のうち、Cycle 4 "Windows on the brain")
- Geddes L.A. (1972) Electrodes and Measurement of Bioelectric Events (Wiley-Interscience, New York).
- Gold C et.al., (2006) "On the origin of the extracellular action potential waveform: A modeling study." J Neurophysiol. 95(5):3113-28.
- Henze, D. et.al., (2000) "Intracellular features predicted by extracellular recordings in the hippocampus in vivo." J Neurophysiol. 84, 390-400.
- Holt, GR (1998) "A Critical Reexamination of Some Assumptions and Implications of Cable Theory in Neurobiology."(pdf) Ph. D. Thesis, California Institute of Technology, Computation and Neural Systems Program
- Holt GR and Koch C. (1999) "Electrical Interactions via the Extracellular Potential Near Cell Bodies". Journal of Computational Neuroscience 6:169-184
- Kandel ER, Schwartz JH, Jessel TM, eds. (2000) "Principles of Neural Science", 4th ed. McGraw-Hill:New York, NY.
- Llinas R, Nicholson C (1974) "Analysis of field potentials in the central nervous system." In: Handbook of EEG and clinical neurophysiology (Stevens CF, ed.), pp. 61-85. Amsterdam: Elsevier.
- Logothetis NK. (2008) "What we can do and what we cannot do with fMRI." Nature 12;453(7197):869-78. (とくにsupplementary informationの"Neural Signals"のセクション)
- Mitzdorf U. (1985) "Current source-density method and application in cat cerebral cortex: investigation of evoked potentials and EEG phenomena."(pdf) Physiol Rev. 65(1):37-100.
- Nunez, P. L., & Srinivasan, R. (2006). Electric fields of the brain : The neurophysics of EEG (2nd. ed.). New York : Oxford University Press.
以上です。それで、いろいろまだ不備はあるのですが、なにがいちばん納得いってないかというと、volume conductionでのpassiveなcurrentという概念とdipoleの概念と電場の形成の概念との関係がまだ明確でないと思うのです。Dipoleによる電場の形成はdipoleの周り全体に起きることですが、passiveなcurrentの話は電荷保存の法則に従うもので、細胞膜のかたちによって影響を受け、ニューロンのサイズを超えて分布することはありません。よって二つは別ものなのだけれど、このへんをきっちりと書いている資料というものがあまりなくてわかりにくい。
それから、現在の説明(図2、図3)でもそうなのだけれど、field potentialそのものを使ってsinkとかsourceとか議論するのは正しくないわけです。しかし、われわれが慣れているのはそういう説明で、このへんが混乱の元になっているのではないかと思います。(正しい説明はたぶん、Vext = f(Im)という関係を示したあとすぐに、CSD解析でsource-sinkの推定をしたあとで、それが[activeなイオンの移動による電流]と[passiveな閉回路を作るための電流]のどちらなのかを解剖学的見地などから推測する、というものだと思われ。それはそれでわかりにくいけど。)
また、そうやって考えると、海馬で記録されるfield EPSPというのが、sinkそのものではないものを使って興奮性シナプス伝達を見るという簡便法である、ということも言えるのではないかと思います。
だいたい今年はこんなもんです。つづきはおそらくまた来年の今頃となることでしょう。ではまた。
# Ryohei
簡潔にまとまっていて、とてもよいですねー。PDFにして、講義ノートも公開してください!そういえば、もうすぐ岡崎でセミナーする予定です。お会いしたいですねー。
# pooneilどうもです。Ryoheiさん、Science出版おめでとうございます。
講義ノートの公開、いいですね。とりあえず現在のエントリのPDFファイルを作ってリンクしておきます。
内容ですけど、間違いはなかったですか? Vmの1階微分のあたりとか、それからdipoleによる電場の形成とleak currentの関係とか、直せるところがあったらぜひコメントしてください。
岡崎のセミナーは生理研ですか? 分子研ですか? ぜひお会いしましょう!
こんにちは。文献までコンパクトにまとめられていて、とても参考になります。
このエントリについて同僚と議論しているときに話題になったのですが、Single-unitのところではVextが時間微分になっているのに、細胞膜電位と細胞外電位との関係の導出のところでは空間微分になっているのはなぜなのでしょうか。
また一回微分、二階微分、2回微分という書き方が混在しているけど、どれが正しいのかというのも議論になりました。
僕は学生の頃一階微分、二階微分という風に教わりました。
なんだか指摘ばかりで申し訳ありませんが、
E(x,y,z) = -grad(V(x,y,x))
は
E(x,y,z) = -grad(V(x,y,z))
のタイポでしょうか。
# pooneilどうもです。Randy Pausch亡くなってしまいましたね。
>細胞膜電位と細胞外電位との関係の導出のところでは空間微分
しまった、ここでこの式を持ってくるのは間違いでした。とりあえず消しときます。Cable theoryあたりをもうすこし加えるy予定です。
>一階微分、二階微分
「2階微分」で統一しておきました。
>V(x,y,x)
直しておきました。
いちおうHodgkin-HuxleyのPropagationモデルでは∂2Vm/∂z2 ~∂Vm/∂tだから、その式でもよいのかも。というか、H-Hモデルなら、Vext ∝ Σi = -Cm dVm/dtでよい気がします。
# pooneilどうもありがとうございます。Im-Vmの関係だけ考えればよいというのに気がつきました。もうすこし考えてアップデートしてみます。
# Koidarman良い具合にまとまってて読んでて楽しかったです。もとは物理出身なのに、この手の数式をいじって細胞外記録データを記述するということをやったことがありませんでした。さすがです。
その昔、細胞外記録は電磁波を読み取っているんだ、という話を聞いたときに、ためしに電線にパルス電流を流して、それを近傍においた電極で読み取る計測をしてみたことがあります。
パルス波形をいろいろ変更しても、読み取った波形は結局アンプのフィルターのせいでか二相性や三相性の波形となることがわかって、パルス波形を適当にEPSPみたいな形にすると、いつも見ているような神経活動の波形が現れたので、それで納得していました。
# pooneil>ためしに電線にパルス電流を流して、それを近傍においた電極で読み取る計測をしてみた
どうもありがとうございます。さすが、これは物理やってる人の行動ですね。生理学実習でaxonを刺激して、みたいなのをやってるところだとこのへんまで勉強してるんでしょうね。
2008年07月21日
■ Saccadic suppression: 藤田昌彦先生からのコメント
以前のChronostasisのエントリ(20040609)に、法政大学教授の藤田昌彦先生からコメントをいただきました。どうもありがとうございます。読者の方の目に触れやすくするために以下に転載させていただきました。これではここから:
"saccadic suppression"の説明ですが,もしこれがVolkmann (1978) に基づくものなら,論文に記載された通りに,彼女の用いた刺激は極めてコントラストの低いものです.この抑制は存在しても極めて弱く,これで日常生活の"saccadic suppression"は説明できません.Saccade最中も視覚像は脳に伝わっており,むしろマスキングに基づくsaccadic omissionという説が確かだと考えます.
F. W. Campbell, and R. H. Wurtz: "Saccadic omission: why we do not see a grey-out during a saccadic eye movement.", Vision Res, 18, 10, pp. 1297-1303 (1978)
以上です。どうもありがとうございます。いま元エントリを再読してみましたが、わたしの"saccadic suppression"という言葉の使い方に混乱があったように思います。
ご紹介いただいたCampbell, and Wurtz (1978)は「サッカード中の網膜上の画像をわれわれが意識しないこと」を説明するメカニズムとして、サッカード中の視覚の閾値を上げるような"saccadic suppression"とサッカード前後の視覚情報でマスクする"saccadic omission"という二つの対立する仮説を取り上げて、"saccadic omission"の優位性を主張しています。また、Rossらによるレビュー(TINS 2001 "Changes in visual perception at the time of saccades")でもサッカード中の視覚の感度をsuppressする過程としてsaccadic suppressionという言葉が使われています。
いっぽうでさいきんでは"saccadic suppression"という言葉が「サッカード中の網膜上の画像をわれわれが意識しないこと」という現象のこと自体を指すようになっているように思います。(心理物理学者ではなくて神経科学者による言葉の濫用という側面はありますが。) たとえば、Science 2002 "Neural Mechanisms of Saccadic Suppression"では、
In normal vision our gaze leaps from detail to detail, resulting in rapid image motion across the retina. Yet we are unaware of such motion, a phenomenon known as saccadic suppression."
としていますし、また、Wurtz本人の最新のレビュー(Vision Research 2008 "Neuronal mechanisms of visual stability" Robert H. Wurtz)でも、
This high speed of the saccade sweeps the image of the visual scene across the retina producing a blur. Yet we are usually unaware of this blur. This lack of vision during saccades is referred to as saccadic suppression or, to emphasize the lack of awareness of sweep, as saccadic omission
と書いています。この意味においては、saccadic suppressionという現象において、「サッカード中の視覚の閾値を上げる」および「とサッカード前後の視覚情報でマスクする」というふたつのメカニズムがあって、それらがどのくらい寄与しているのか、どういう脳内メカニズムによって達成されているのか、という議論と捉えるべきかと思います。どちらの意味においても元エントリの"saccadic suppression"の言葉の使い方は良くなかったようです。ご指摘ありがとうございます。
ちなみに先述のWurtzのレビューではgating説(つまりサッカードの指令のcorollary dischargeがsuppressionに関わっているとする説)の方に関して、そのeffectはmodestとしていて、LGN関連であるとするデータを列挙しています。(Motionやdisplacementに関するdetectionに関してはShioiri and Cavanaghをreferしてます。) ただ、effectの起こっているsiteに関してはdiscussionのところで面白いことを言っていて、
One idea about a possible added source of that suppression, though a novel one, is that it could be provided by the SC superficial layer neurons, which show a CD based suppression, and that could project upward via the pathway from SC through pulvinar to cortex.
ということで上丘の関連を示唆しています。Wurtzの上丘中心主義もここまで来たかというかんじもありますが、これには伊佐研の仕事も関連しています。Ratの上丘sliceでの仕事なのですが、SGIからSGSへ投射するinhibitoryなconnectionを見つけたという論文(PNAS 2007 "Identity of a pathway for saccadic suppression")があります。これを元にしてWurtzのレビューでは、SGIでのsaccadeのシグナルがcorollary dischargeとしてSGS->pulvinar->cortexの回路をゲートするのが、サッケード中の網膜入力のsuppressionに関わるのではないか、というアイデアを提唱しています。(ちなみにPNAS 2007はcommunicated by Wurtzです。)
ちなみに藤田先生のグループからこの問題を直接扱った論文が出ています:信学論誌 「サッカード抑制説と脱落説の同一手法による検討」。
- / ツイートする
- / 投稿日: 2008年07月21日
- / カテゴリー: [サッカード中の知覚安定性]
- / Edit(管理者用)
2008年07月13日
■ 神経科学大会シンポジウム無事終了しました!
どもども。ここ最近出張しまくりの激動の日々を送っておりました。5/9-16でフロリダのNaplesでVSSに参加。6/19-23に台北で開催されたASSC2008に参加して、帰国したその日にそのままLAに飛んでUniversity of South CaliforniaのLaurent Ittiのところに2週間弱滞在してHFSP関連のコラボレーションの仕事をしてきました。そのあいだにカルテクにも見学しに行って、Christof Kochのラボでトークをさせてもらってきました。7/6に帰国してすぐに神経科学大会というかんじで、まだ時差ボケが抜けきってません。来週の火曜日(7/15)には京大医学部で開催される電子情報通信学会の7月のNC研研究会でトークです。ここまででひとくぎり。しばらく岡崎でおとなしくしているつもりです。お次のイベントはいよいよ生理研研究会(9/11-12)でしょうか。
神経科学大会のシンポジウムのほうですが、会場は立ち見の方も出て、熱気ある雰囲気の中で進めることができて非常に良かったのではないかと思います。質問もたくさん出たし。あれだったら総合討論の時間を入れても良かったかもしれませんね。私自身に関しては、座長としてのしゃべりとか、トーク自体とかけっこうひっかりまくってましたが、もう勢いでなんとかしたというかんじです。
また別にレポート書くかもしれませんけど、まずは来てくださったみなさま、どうもありがとうございました。以下にシンポジウムに言及してくださったブログをリンク:
* スウィングしなけりゃ脳がない!
* ◎脳研究のタマゴ◎
* 大「脳」洋航海記
shokou5さんとは前日の懇親会でもお話を。私からは、フジロックのマイブラ行きたいけど行けない!とかこれからは萌えとロボティクスだとか(またかYO!)。
vikingさんも見に来てくれてシンポジウム後にお話ししました。
(前回のエントリにコメントしてくださったatsushiさん、Hikaru.Tさんのコメントは内容を考慮してこちらに移動させていただきました。)
# atsushi
初めまして!いつもblog拝見しております。
神経科学大会の口演お疲れ様でした。
もうほとんどあのセッションがメインで行ったので、とっても楽しかったですし、visual awarenessのセッションは今大会で一番盛り上がってた気がします。
僕は学部生でまだ脳について勉強し始めたばかりなので全然知識もないですが、視覚意識にすごく興味があるのでこちらのblogでもかなり勉強させて頂いてます。
お話できる機会があればよろしくお願いします♪
あ、もし両側V1破壊したらvisual awarenessは片側のときと同じようにdSCを使ってretainされるんでしょうか?
pooneilさん Neuroscience2008、お疲れ様でございました!atsushiさんと同じく、僕も今回のシンポジウムを一番楽しみにしておりました。想像以上に興味深く拝聴させていただきました。本当にありがとうございました!!
# pooneilatsushiさん、Hikaru.Tさん、どうもありがとうございます。この分野に興味を持っておられる方がたくさんいることをシンポジウム会場でも実感しました。来年の神経科学大会は開催地が名古屋で、当ラボの伊佐教授が大会長です。Plenary lectureのひとつはChristof Kochが確定しています。おそらくは来年も意識関連の話題を扱うことができるのではないかと思います。ご期待ください。
atsushiさんの質問ですが、両側の条件を扱った研究は大昔に行われていますが、この条件でのdSCの寄与を直接検討した研究はないと思います。おそらくは片側のときと似たものが見つかるのではないかと予想していますが。ちなみに両側の条件を扱った研究で有名なのは1970年代にニコラス・ハンフリーによっておこなわれたものです。単行本で「赤を見る―感覚の進化と意識の存在理由」ニコラス ハンフリーというのが出版されていて、これの中にけっこう記述があります。ご興味ありましたら読んでみるとよいかもしれません。
吉田さん、
土谷です。
視覚意識のシンポジウムは結構評判よかったみたいっすね。リンク先のページも見てきたら、俺よりもさらに若い世代の人らが興奮してたみたいで、よかった。これなら、来年も何かネタを考えて、やっても良いかも、と思い始めました。クリストフが、トークだけじゃなくて、シンポジウムみたいなものも、やりたい、みたいに言ってたし。
ただ、何人かの感想だけだと、フィードバックとしては物足りないですね。ASSCみたいに学会後アンケートがないので、これらの意見が客観的に評価できないのが問題です。そういうアンケートの集計とかって、おそらく外部に頼む必要があるとか、色々大変だと思いますが、将来学会を改善していくには非常に有効だと思いますので、来年の時には是非実施してください!
総合討論もいいかとは思うんですが、あんまり質問とか出て来ないとサムイので、TICSみたいに、"outstanding questions" を各演者が用意しといてそれをたたき台に最後に20分ぐらい時間を取るとかすれば良かったなと思います。
あと、来年の会場は会場内で移動しやすいところが良いです。マジで自分のシンポジウムに遅刻しそうだったので...
# pooneil土谷さん、シンポジウムではどうもありがとうございました。
> クリストフが、トークだけじゃなくて、シンポジウムみたいなものも、やりたい、みたいに言ってたし
これいいですね。ぜひ実現できるように各方面に働きかけたいと思います。
フィードバックに関してですが、学会後のアンケートは今回もありました。(要旨集の冊子に挟まれてる。) ただ、特定の講演に対する感想というのはありませんでした。ですので、たとえば現状のアンケートに「いちばん面白かった講演(シンポジウム・ポスター)はなにですか?」というようなものを作って集計・発表するとかいうやり方はあるかと思います。
総合討論に関してですが、じつは今度9月に行う生理研研究会(「動機づけと社会性の脳内メカニズム」)では総合討論の時間を取ることにしているのですが、まさに土谷さんが言うのとまったく同じことを計画しています。「TICSみたいに、"outstanding questions" を各演者」に用意しておいてもらうといい、と松元さんにもメールしていたのですが、TICS、というところまでまったく同じなのですごいびっくりしました。(それとも前にこの話聞きましたっけ?) さらに、あらかじめ若手の人から総合討論でのお題を募集しておいて、面白いお題を出してくれた人にトラベルアワードを出す、ということも計画してます。このくらいやればそれなりに盛り上がるんではないか、というわけです。研究会の方はそんなかんじでそれなりに準備が可能ですけど、学会でやるのは難しいかも。
来年の生理研研究会も秋にやることになるかと思いますが、もしかしたら神経科学大会(9/16-18)に近接して設定して、シンポジウムの招待講演者にそちらにも出てもらうとかそういうやり方ができるかもしれません。この場合は「国際研究集会」という枠組みになると思いますが。もうすこし考えてみます。
来年の神経科学大会の会場はもう決定しています。今年ほど複雑ではありませんが、建物の右と左を行ったり来たりしないといけないかもしれません。ちなみにフロアマップ。
# vikingどうも、シンポジウムは大変興味深く拝聴いたしました。nhpには詳しくないもので、質問が冗長になってしまったかもしれません。今後はもう少しかっちり論点をまとめてから伺うようにいたします。
自分のblogでも書きましたが、東京国際フォーラムは思った以上に使いにくかったですね・・・名古屋は5年前に行ったことがあって使いやすかった記憶(でも周囲に食事できるところが全然なかった気も)があるので、期待したいと思います。
# pooneilvikingさん、どうもありがとうございます。ブログをリストに追加しておきました。
そうです、名古屋は昼ご飯が問題なのです。金山まで出るとそのまま戻ってこなかったりとかして。宮下研のときの横浜での神経科学大会みたいにフードコートとか作るとかそういう手が必要かもしれません。
匿名問題ですが、私自身は名前を出していることからもおわかりのように実名派です。わたしの場合、実名で情報発信することのメリットはむちゃくちゃ大きかったと思います。自分が名前を出してさらけ出したからこそ、安心してコメントを書き込んでくれて、自分が何者であるかも明かしていただけた方も多かったのではないかと思います。ここを元にして出来た人の繋がりに感謝しております。
とはいえ、実名であるが故に、ポスドク問題など、荒れる問題には意識してタッチしないようにしたりとか、それなりに気を遣っているのも事実です。(言いたいことはあるんだけれど、言葉尻を取られないようにきっちり書かないといけないかと思うと気が重い。) これはわたしのnhpという言葉遣いを理解しているvikinさんならご存じではあるかと思いますが。
とりとめが無くなってきたのでこのあたりで。
>pooneilさん
そうですね。どちらを選択するかは悩ましいところですが、まぁそれぞれの考え方があっても良いかなと僕は考えております。もっとも、僕のblogはあまりにも業界内では有名になりすぎていて、素性を隠す理由がもはやなくなりつつありますが(汗)。
pooneilさん、大変お忙しいスケジュールの中、質問に丁寧に答えて頂きありがとうございました。m(__)m
リンクも貼っていただき、ありがとうございました。
今たまたま京大にいるので明日の講演も拝聴させて頂くと思います(ストーカーみたいですが笑)。楽しみにしています。
では。
いつも楽しく拝見させて頂き、大変勉強させて頂いています。意識に興味があってサルの電気生理を始めた僕としては、先日のシンポジウムは大変刺激的で興奮してしまい、一枚脱いでしまう程でした(実際会場自体に熱気がこもっていたのもあるかもしれませんが)。
僕が知らなかった事が多すぎたせいかもしれませんが、日本の意識研究があのセッションで確固たるものになりつつあることを示された、そんな歴史的な瞬間であるように思われました。
神経科学者SNSのイベントで学会の時に飲み会があるようなので、是非これから先、参加させて頂こうと思いますので機会がありましたらよろしく御願いします。
# pooneil桑原さん、どうもありがとうございます。
歴史的な瞬間、とはさすがにびっくりですが、これからもいろいろ仕掛けていくつもりです。
よろしくお願いします。
いま過去のエントリを見てたら昨年自分で書いてた>>「TICSみたいに、"outstanding questions" を各演者」(20071019 http://pooneil.sakura.ne.jp/archives/permalink/001119.php)
2008年07月08日
■ 内側前頭皮質のセルアセンブリは行動に依存して動的に変化する
Buzsákiラボへ留学中のしげさん(ブログ(休止中)へのリンク)の論文がNature Neuroscienceに掲載されてます。おめでとうございます。
Nature Neuroscience 11, 823 - 833 (2008) "Behavior-dependent short-term assembly dynamics in the medial prefrontal cortex" Shigeyoshi Fujisawa, Asohan Amarasingham, Matthew T Harrison & György Buzsáki
Figure 4がpositionによってcross correlogramが変化するというデータなのだけれど、これがなんとmonosynaptic connectionなんです。1-2msあたりに鋭いピークがあって。これは印象的。
- / ツイートする
- / 投稿日: 2008年07月08日
- / カテゴリー: [セルアセンブリ (cell assembly)]
- / Edit(管理者用)
# しげ
こんにちは。ご無沙汰しております。論文を紹介していただき、ありがとうございます!紹介していただいたとおり、この論文では「cross-sorrelogram の t=2ms あたりに sharp peak がある」という結果だけから「monosynaptic connection が存在する」と主張しているわけですが、extracellular のユニット記録だけでそれが主張できるのか、という点に関してはやはり弱く、いろいろとクレームもつきました。
それで、われわれの主張の論拠としましては、「cross-sorrelogram に sharp peak がある理由として、もし細胞A、Bが monosynapse でつながっているのではなく、単に別の細胞から common input を受けているだけだとしたらどうか?その場合、発火タイミングの精度は荒くなるだろうから、CCG の peak 幅はもっと太くなるはずであろう」という仮説のもと、細胞Bの spike time に人工的に微少な "jitter" (<5ms) を加えて、CCG のピークの sharpness な構造が崩れるかどうかを統計的に検定する、という統計的な reasoning を用いた、というものです。(共著者の真ん中の二人は統計学者です。)
(まあどちらにしても "inferred" monosynapstic connection であることには変わりはないのですが。。)
ところで、夏に一時帰国する予定なのですが、そのときちょうど生理研に寄る用事ができそうなので、そのとき pooneil さんにお時間があればお話などさせていただけたら、と思ってます。ではでは。
# pooneilレスポンスどうもありがとうございます。
斜め読みでのコメントで恐縮です。(ところでいま、publisherのサイトからPDFが落とせなくないですか?)
神経科学大会でガヤに会ったときにこの論文について少し教わりました。統計学者が二人入っているというところがミソのようですね。
「extracellular のユニット記録だけでそれが主張できるのか」、これについてはシステム関連では外山先生の論文やC. Reidの論文などで同様な論法が使われているのですが、さすがにいまどきのrodentでのmultiunitの研究だとそのへんまできっちり対処しないといけないのですね。
「生理研に寄る用事」の話はホストする方は決まっていますか? 必要でしたらお声をかけてください。お力になれるかと思います。
ともあれ、またそのときに。
お勧めエントリ
- 細胞外電極はなにを見ているか(1) 20080727 (2) リニューアル版 20081107
- 総説 長期記憶の脳内メカニズム 20100909
- 駒場講義2013 「意識の科学的研究 - 盲視を起点に」20130626
- 駒場講義2012レジメ 意識と注意の脳内メカニズム(1) 注意 20121010 (2) 意識 20121011
- 視覚、注意、言語で3*2の背側、腹側経路説 20140119
- 脳科学辞典の項目書いた 「盲視」 20130407
- 脳科学辞典の項目書いた 「気づき」 20130228
- 脳科学辞典の項目書いた 「サリエンシー」 20121224
- 脳科学辞典の項目書いた 「マイクロサッケード」 20121227
- 盲視でおこる「なにかあるかんじ」 20110126
- DKL色空間についてまとめ 20090113
- 科学基礎論学会 秋の研究例会 ワークショップ「意識の神経科学と神経現象学」レジメ 20131102
- ギャラガー&ザハヴィ『現象学的な心』合評会レジメ 20130628
- Marrのrepresentationとprocessをベイトソン流に解釈する (1) 20100317 (2) 20100317
- 半側空間無視と同名半盲とは区別できるか?(1) 20080220 (2) 半側空間無視の原因部位は? 20080221
- MarrのVisionの最初と最後だけを読む 20071213