[月別過去ログ] 2004年07月
« 2004年06月 | 最新のページに戻る | 2004年08月 »2004年07月31日
■ Nature 7/29
"Neurons compute internal models of the physical laws of motion."
Vestibular nucleiと小脳のrostral fastigial nucleusからの記録で慣性などの物理法則を内部モデル化している証拠を提出。
Vestibular系ってはずかしながら私なんにも知らないんですよね。VOR (vestibulo-ocular reflex)とかとくに重要であるにもかかわらず。というわけでこれまでNature and Scienceで関わってるっぽいもの。
- / ツイートする
- / 投稿日: 2004年07月31日
- / カテゴリー: [Paper archive]
- / Edit(管理者用)
■ Science AOP
"Distinct Ensemble Codes in Hippocampal Areas CA3 and CA1." Edvard I. Moser。
先週のNature("Comparison of population coherence of place cells in hippocampal subfields CA1 and CA3.")へのコメンタリかと思ったら違うようです。"We examined the function of this structural differentiation ..."なんて書いてある。CA3-CA1のfunctionのdissociationを出すのに競走をしていたようですな。しかも微妙にずれた出版日。Natureの方はreceive 1/8、accept 6/10で online publicationが 6/30、Scienceの方はsubmit 5/13、accept 7/8でonline publicationが 7/22、というわけでNatureの方が早かったけど、(その情報を察知してか、)Scienceのほうが追い上げたということらしい。Journal clubで採り上げる人はこの二つをセットにしてプレゼンしないと、あっちの方はどうなのと問い詰められること必至ですな、これは。
内容については紙で出版されてからにします。ていうかうちではAOPのpdf取れないんですよね。
Edvard I. Moserといえば私にとってはScience '98 "Impaired Spatial Learning after Saturation of Long-Term Potentiation."です。Barnes and McNaughtonの"Recovery of spatial learning deficits after decay of electrically induced synaptic enhancement in the hippocampus."みたいな、LTPを海馬全体に惹き起こしてやると空間学習がimpairされるというやつでした。なんかすげ―感心した憶えがあるのだけれど、どの点だったか思い出せない。SFNでこの内容を見たときは夫婦並んでポスターで、そろって美男美女だった印象がある、どうでもいいことだが。
Science '02 "Place Cells and Place Recognition Maintained by Direct Entorhinal-Hippocampal Circuitry."のほうはCA3 -> CA1の入力を取り除いてentorhinal cortex -> CA1の入力だけで何ができるか、という議論だから、今回のCA1-CA3のfunctional dissociationへ向かうのは当然の流れだったと言えましょう。
- / ツイートする
- / 投稿日: 2004年07月31日
- / カテゴリー: [Papers_unclassified]
- / Edit(管理者用)
2004年07月30日
■ 伝染するあくび("contagious yawning") つづき。
7/27のCorreggioさんへの返答から膨らませてみました。
Cognitive Brain Research '03 "Contagious yawning: the role of self-awareness and mental state attribution."(pdf)についてまとめます。
被験者にヴィデオであくびをしている人の映像を見てもらって、あくびをするかどうか数えます。この数(=あくびが伝染しやすいかどうか)とさまざまな心理テストとの相関を調べてやるわけです。
まず、自己意識の障害についてのテストであるSPQテストのスコア(自己意識に障害があるとこのスコアが上がります)と伝染したあくびの数は反比例します。つまり、自己意識をしっかり持っている人ほどあくびが伝染しやすいのです。
また、自分の顔を見つけ出すテストでも、このスコアが高い(自己意識を持っている)人ほどあくびが伝染しやすい。
それから、「心の理論」研究で使われるお話を聞いてその中に含まれるfalse belief(だれがなにを勘違いしているか)およびsocial faux pas(だれがどんな良くないことをしているか)を報告するテストをすると、social faux pasのほうのみcontagious yawningの数と相関があるというのです。著者はこのことから、他者の立場に立って考える能力がcontagious yawningに関わる、とします。(ここはなんかデータが汚いわけですな。)
ところでこの論文、被験者が普段からどのくらいあくびをするかについては記載がないのではないでしょうか。そうしたら、あくびが伝染しやすいのか、ちょっとしたきっかけですぐにあくびが出やすいのかわからないのだけれど。ほかの論文でも、コントロールとしてヴィデオの中で微笑む人をみて何回微笑むか(当然ゼロ)というのを出しているのを見たことがあるけれど、そうではなくて、コントロールとは、退屈なビデオを見ているうちに何回あくびをするか、ではないでしょうか。このへんどうしてるんだろ。
というわけで7/27のCorreggioさんへの返答につなげると、Imitationとcontagious yawningとはミラーニューロンシステムを共有している一方で、幼児のimitationにはまだ自他の分離(self-awareness)と他者の顔色をうかがうような社会性(心の理論を必要とするようなmental-state attribution)とが欠けていて(Cognitive Brain Research ’03)、それらがあってはじめてcontagious yawningのように高度な認知的行動が起こる、ということのようですね。そしてもちろん、ミラーニューロンシステムとはたぶん、ただのimitationにのみ関わるのではなくて、そのようなimitationを通して心の理論と共感とを獲得していくダイナミックな過程に関わっている、ということになるのでありましょう(Rizzolattiは読んでないけど、こういうことを言っているのではないですか)。
■ Empathy
せっかくなのでempathy、ミラーニューロンあたりで検索して関連論文漁ってみました。
- PNAS '03 "Neural mechanisms of empathy in humans: A relay from neural systems for imitation to limbic areas." Premotorやsuperior parietalだけでなく、insulaやamygdalaで差分が出る、というのがミソのよう。
- "The manifold nature of interpersonal relations: the quest for a common mechanism." Vittorio Gallese。"EARLY AND MATURE IMITATION"なんてセクションもある。
- "What imitation tells us about social cognition: a rapprochement between developmental psychology and cognitive neuroscience." アブストにこんなこと書いてある。"We marshal evidence that imitation recruits not only shared neural representations between the self and the other but also cortical regions in the parietal cortex that are crucial for distinguishing between the perspective of self and other."
- "Empathy for Pain Involves the Affective but not Sensory Components of Pain." Raymond J. DolanとChris D. Frith。2/25にChris Frithがまたらしい感じ(empathy)を扱っている、とだけ書きましたが、これがCorreggioさんの言及したFrithのfMRIですね。"their loved one"ってすごいなあ。でも痛みへのempathyだったら、自分の子供を使った方が強烈だと思う、って何を考えているんだ。
- "Both of Us Disgusted in My Insula: The Common Neural Basis of Seeing and Feeling Disgust." Giacomo Rizzolatti
# Correggio
心の理論は、大体4−5歳から獲得するという話しですが、自他の分離のついた模倣はもっと早いようですね。の辺は最後にリンクを張ってある論文に詳しいです。従って、ミラーニューロンシステムは、発達にしたがって、ステップアップしていくんでしょうが、心の理論はある時期のその表出を見ているということでしょうか。empathyについては、PNAS03の論文が、顔の表情の模倣をしたときのイメージングで、純粋にemotionの共有といえないのではないかという疑問があります。それに対して、Neuron2003ではRizzolattiたちが、実際に匂いをかいだときとそれをかいだときの表情の観察で、Isulaやcingulateが共通して活動するのを見ています。Frithのところからは、16ペアの恋人同士を被検者にして、その片方を痛めつけた時のもう片方のfMRIをとってます。
# pooneilCorreggioさん、ありがとうございます。関連論文、リストに入れておきました。自他の分離のつかない模倣、自他の分離のついた模倣、心の理論、という順番なのでしょうか。なんにしろ、ご指摘のPhilosophical Transactions、読んでみます。
# メシダ初めまして。
ryasudaさんのリンクから来ました。
私はこの「あくびが伝染する」という記事を読んだとき、
「年を取ると涙もろくなる」のも、同じメカニズムなんじゃないかと思いました。
どちらも、共感能力が高くないとできない(他人の気持ちのわからない、人生経験の少ない幼児にはできない)ことだからです。
ま~、これは私のただの思い付きです。
どうもはじめまして。お返事遅れました。「年を取ると涙もろくなる」、なんかさいきん実感するようになりました。発達(子どもから大人へ)や老化(大人から老人へ)だけでなく、なんかその中間でもいろいろ変化が起こってるんだろうなあと思います。
2004年07月29日
■ Nature Neuroscience 8月号
- "A morphological basis for orientation tuning in primary visual cortex." David Fitzpatrick。図だけしか見ていないけどこれすごいな。V1のlayer 2/3ニューロンのorientation selectivityがどうやってできるかを入力線維の走行パターンによって説明しようとするんだけれど、News and Views ("A shrewd insight for vision." Martin Usrey)のFigure 1にあるように、それがlayer 4からlayer 2/3への投射がねじれていることによって実現されている(分かりにくいから図を見てください)、ということを示したというのです。これはおどろき。普通そういったorientation selectivityは層を垂直に延びるaxonがdivergentに投射していること(そのときには今回のようなasynmetricalな構造は想定されていない)および層に平行な方向のlateralなconnectionとによって実現されるものだと思われていたわけですから。このことはV1のpyramidalニューロンが垂直方向に延びているとするコラム仮説の修正を要請するものだし、0-180degreeの方位を表象する領域がハイパーカラム的な構造をしていることを予言します。 また、これは重要なことだと思うのですが、V1の (optical iamgingで作る)orientation mapでのpinwheel center構造(NVのFigure.1での黄、緑、青、紫、赤に囲まれた領域。orientation selectivity map上での鞍点のようなもの。これもFitzpatrickやBohnhofferとかが明らかにしてきたもののはずですが)について、今回の論文で出てきたような構造(layer 4での回転する軸の中心に対応する部分がlayer 2/3でpinwheel centerになっていると考えればわかりやすい*1)から説明することもできます。 これまでの論文とどうconsistentなのかも気になるけれど、V1の局所回路についてずっとやってきたFitzpatrickに言われるのでは何もいえなくなってしまいます。
- "Prediction of immediate and future rewards differentially recruits cortico-basal ganglia loops." 7/3のコメント欄でmmmmさんが言及した、銅谷先生のところでのfMRIの論文。
- "Underlying principles of visual shape selectivity in posterior inferotemporal cortex."
*1:いや、中心が対応するという言い方も正確でないか。点と点との一対一対応にはないわけだから。なんにしろ、すぐにモデルが作れそう。
■ Kinksを歌って泣く。
自分で歌って自分で泣くのです。
"Everybody's in Showbiz"のclosingの"Celluloid Heroes"をギターで弾き語りしながら。僕の人生がノンストップのハリウッド映画だったらよいと思う、だってフィルムの中の主役たちは痛みを感じることがないのだから。あーもう泣いた。子供たちには迷惑そうだ。
"Arthur - Or The Decline And Fall Of The British Empire"の"She's Bought a Hat Like Princess Marina"。彼女はマリーナ王女のような帽子を被り、公的行事(窓掃除や階段掃除)に参加する。彼はアンソニー・ギデンス(イギリスの有名な社会学者)のような帽子を被るがロールスもベントレーも買うお金がなくて中古のフォードを買う。んでブリッジ越えた三番で泣くのですな。やっぱり子供たちには迷惑そうだ。いいじゃん(<-酔っ払ってます)。
■ "フランツ・フェルディナント"
なんて書いておいたら。
"フランツ・フェルディナント"
アレックスのボーカルはキンクスのレイ=デイヴィスを彷彿させるメランコリックな声質(そしてメンバーのコーラス)。曲の転調の仕方さえもキンクスしている。(新訂カンタン系より。)
これは聴きに行かなくては。
2004年07月28日
■ さいとにふりがながつきます
ひらひらのひらがなめがね。 ソースはいつものARTIFACT@ハテナ系。
さっそくやってみました。
http://trans.hiragana.jp/ruby/http://d.hatena.ne.jp/pooneil/
うーん、キュート。英語の読みを付けてくれると完璧なのだけれど。
こぐにてぃぶ ぶれいん りさーち ぜろさん
Cognitive Brain Research '03
みたいに。あと、(笑)に「かっこわらい」というふりがなが付くのは笑った。
■ Vision Research 7月号
"Improvement of visual acuity by spatial cueing: a comparative study in human and non-human primates."
"Magnocellular involvement in flanked-letter identification relates to the allocation of attention."
- / ツイートする
- / 投稿日: 2004年07月28日
- / カテゴリー: [Paper archive]
- / Edit(管理者用)
- / ツイートする
- / 投稿日: 2004年07月28日
- / カテゴリー: [Paper archive]
- / Edit(管理者用)
■ Nature Neuroscience 5月号
"Visual and action cues contribute to the self-other distinction."
- / ツイートする
- / 投稿日: 2004年07月28日
- / カテゴリー: [Paper archive]
- / Edit(管理者用)
2004年07月27日
■ 伝染するあくび("contagious yawning") つづき。
関連記事を漁っときます。
まず原著論文:
- Cognitive Brain Research '03 "Contagious yawning: the role of self-awareness and mental state attribution."
- European Journal of Pharmacology '98 "The neuropharmacology of yawning."
- "Psychological Influences on Yawning in Children."
いろんな動物が欠伸をする画像。かわいいです:
その他いろいろ:
- "Neuroscience for Kids - Contagious Yawning."
- "ABCNEWS.com : 20/20: Are Yawning and Laughing Contagious?"
- "Why do we yawn when we are tired? And why does it seem to be contagious?"
- "USATODAY.com - Catching yawns keeps us together..."
- "Yawning: It Isn't About Oxygen Anymore."
- "Contagious Yawning in the Infant."
- "Ask Us @ U of T: Why do we yawn?"
- "Yawning may signal empathy - University Chronicle - Diversions."(グーグルのキャッシュ)
# Correggio
Adultやolder infantでは伝染するけれども、young infantでは、あくびはするけれども伝染しないというところがみそのようですね。赤ん坊の舌だしの模倣はかなり早くから起こるのだけれども、それとはちょっと違う。どちらもmirror neuronsystemが絡んでいるけれども、Rizzolattiたちのいうようにimitationにも異なるレベルがあるということになりますか。しかし、あくびの伝染は、抑制が利かない反射的な感じもしますね。この辺imitation behaviorとも似てますが。松沢さんの論文読まなければ。
# Correggioちょっと訂正。ヒトではolder infantどころじゃなくて5歳でも伝染しないみたいですね。
# pooneilありがとうございます。Imitationとcontagious yawningとはミラーニューロンシステムを共有している一方で、幼児のimitationにはまだ自他の分離(self-awareness)と他者の顔色をうかがうような社会性(心の理論を必要とするようなmental-state attribution)とが欠けていて(Cognitive Brain Research ’03)、それらがあってはじめてcontagious yawningのように高度な認知的行動が起こる、ということのようですね。そしてもちろん、ミラーニューロンシステムとはたぶん、ただのimitationにのみ関わるのではなくて、そのようなimitationを通して心の理論と共感とを獲得していくダイナミックな過程に関わっている、ということになるのでありましょう(Rizzolattiは読んでないけど、こういうことを言っているのではないですか)。あと、私も松沢先生の論文入手したかったのですが、うちのinstituteでは取っていないどころか、webcatで調べたら日本のどこの図書館もとってなさそうなので、別刷りかPDFを著者に請求した方が早そうです。
■ JNP8月号
- "Activity of Rostral Superior Colliculus Neurons During Passive and Active Viewing of Motion." Richard J. Krauzlis
- "Impact of Noise on Retinal Coding of Visual Signals."
- "Functional Stability of Dorsolateral Prefrontal Neurons." Fraser A. W. Wilson
- "Decoding Continuous and Discrete Motor Behaviors Using Motor and Premotor Cortical Ensembles."
- "Optimal Impedance Control for Task Achievement in the Presence of Signal-Dependent Noise." Mitsuo Kawato
- "BOLD Repetition Decreases in Object-Responsive Ventral Visual Areas Depend on Spatial Attention." R. J. Dolan
- / ツイートする
- / 投稿日: 2004年07月27日
- / カテゴリー: [Paper archive]
- / Edit(管理者用)
2004年07月26日
■ あくびが伝染するチンパンジー
新聞に載ってた霊長研松沢先生の発表。たとえば
"チンパンジーもあくび“伝染”、人間以外で初めて確認"(Yomiuri Online)
件の論文は"Proceedings of the Royal Society: Biological Sciences, Biology Letters"に掲載された"Contagious yawning in chimpanzees."で、目次はこちら。"Proceedings of the Royal Society: Series B: Biological Sciences"の姉妹誌のようなものらしい。
アイちゃんにあくびが伝染するところを記録したビデオファイル(mpeg)はこちらから落とせます(フリーです)。ほかのチンパンジーのあくびをモニターで見てアイちゃんがあくびしてます。これがたんなるまねっこではなくて、本当にあくびが伝染しているのか、つまり、あくびのまねっこと本当のあくびではあくびの深さが違うわけで、そのへんまで併せて確かな知見のか興味があります。そもそも、人間のあくびが伝染するって確立しているのでしょうか。というわけでPubMedでcontagious yawningで検索するとありました。
Cognitive Brain Research '03 "Contagious yawning: the role of self-awareness and mental state attribution"
んで、そこにあったreferenceの
- R.R. Provine, Faces as releasers of contagious yawning: an approach to face detection using normal human subjects. Bull. Psychon. Soc. 27 (1989), pp. 211-214.
Abstract: 360 psychology students were divided into 12 experimental groups and participated in a single experimental session. The yawn-evoking potency of variations in a 5-min series of 30 videotaped repetitions of a yawning face were compared with each other and with a series of 30 videotaped smiles to determine the ethological releasing stimulus for the fixed-action pattern of yawning and to understand the more general process of face detection. Animate video images of yawning faces in several axial orientations evoked yawns in more Subjects than did featureless or smiling faces, and no single feature, such as a gaping mouth, was necessary to evoke yawns. The yawn recognition mechanism is neither axially specific nor triggered by an isolated facial feature.
- R.R. Provine, Contagious yawning and infant imitation. Bull. Psychon. Soc. 27 (1989), pp. 125-126.
Abstract: Suggests that the neonate's presumed ability to imitate the facial expressions or gestures of adult models may be the result of ethological fixed-action patterns released by sign stimuli. Contagious yawning (CY) of adults is a precedent for such a facial fixed-action pattern (i.e., a yawn) triggered by a facial stimulus (i.e., an observed yawn). The study of CY can provide insights into both the problem of infant imitation and the more general issues concerning the detection and processing of information about faces. CY also provides a reliable classroom demonstration of released behavior.
- R.R. Provine, B.C. Tate and L.L. Geldmahcer, Yawning: no effect of 3-5% CO2, 100% O2, an exercise. Behav. Neural Biol. 48 (1987), pp. 382-393.
- G. Schino and F. Aureli, Do men yawn more than women?. Ethol. Sociobiol. 10 (1989), pp. 375-378.
"Contagious yawning and infant imitation"なんてのが扱われているということは、上記の松沢先生の論文でも、imitationなのかcontagious yawningなのか、という問題については扱われていそうです。
んで、松沢先生の論文のfull-textは入手できないんだけれど、アブストみるかぎり、この話はself-awarenessと関係してくるわけですな。自己意識、ミラーテスト、共感。このへんに関しては上記のCognitive Brain Research '03のメインテーマでもあります。
追記:"contagious"のつづりを間違えていたのを修正(ガヤサンクス)。
2004年07月24日
■ 武蔵書店
せっかく青山ブックセンターのことを思い出したので、ついでに、と言っては悪いが、武蔵書店のことも思い出してみた。私が茗荷谷に生息していたころ、夜中12時とかになってなぜか妙に本が恋しくなって、自転車乗ってお茶大の正門前から本郷通りと不忍通りの交差点を越え、新大塚の三叉路を越え、都電の踏切を越え、池袋駅の臭い地下道を潜って池袋北口を越えてゆるい坂を下って山手通りと交わる要町交差点の武蔵書店まで通っていたのでした。岩波文庫の品揃えが妙によくて、行くたびに徒然草の章を立ち読みしたり、背伸びしてベルクソンとか買ったりしていました。1980年代後半。なんかあのころ、ABCでも武蔵書店でも水戸街道を松戸へ進む道でも、その先に何かがあるような昂揚感があって、それでなんにもなくて、それでもめげないでまた同じ道を繰り返し進んで、それでも満足だったんです。私はロマンティシズムだけで生きていました。"I could dream all day"*1(by Posies). なんにもなくてもいいと思ってたんです。"I remember the good-old days"(by Neil Young)ですな。いいじゃん、べつにこういうのも。今日からこのサイトは私の生まれ育った東京での自転車での日々を追憶する日記となりました(<-ウソだって)。次回は広尾から南砂まで明治通り(環状五号線)全制覇ルートです(<-マジかよ)。その次は、外苑東通りとつながるはずだった幻の環状三号線の名残りの播磨坂、言問通り、三つ目通りな(<-もういいって)。
追記;やっぱりいくつか間違っていたので修正。
*1:追記:All dayに仮定が入っている仮定法だから、canではなくてcouldでした。「一日中だって夢を見ていられる」ですな。
2004年07月23日
■ Nature 7/22
- "Conceptual precursors to language."
- "Comparison of population coherence of place cells in hippocampal subfields CA1 and CA3." JAMES J. KNIERIM。海馬のCA1とCA3とでの違いを示しています。さらに関連論文として"A Double Dissociation between Hippocampal Subfields: Differential Time Course of CA3 and CA1 Place Cells for Processing Changed Environments."があります(情報提供fromガヤ、サンクス)。以前にも書いたように、area間のinteractionを見る、というのがsingle-unit recordingを越えるための最小限の一歩なわけですが、CA3-<CA1はひとつのシナプスしか結合してないわけで、そのあいだでの違いを見出すというのは凄いしすばらしい。もちろん、この機能局在は大いにありうることでありますが。</li>
- / ツイートする
- / 投稿日: 2004年07月23日
- / カテゴリー: [Paper archive]
- / Edit(管理者用)
■ 日記・つぶやき・思いつきは書かない
日記・つぶやき・思いつきは書かない
技術系サラリーマンの交差点より。
- 日記は書かない
- つぶやきは書かない
- 思いつきで書かない
■ あなたのブログはどのタイプ?
あなたのブログはどのタイプ?
ソースはARTIFACT@ハテナ系。
いつもどおり遅延を入れて今さら貼ります。
私がやっていることもオンラインにある論文に言及してそれを記録してゆく、という意味ではブログの定義に含まれるのでしょう。話題は専門的なところに限られているのだけれど。私がやっていることは、そういった読む価値のある論文をメモり、必要なら最小の説明を加えるだけでなく、論文をただ読んだだけでは出てこないような指摘を加え、その論文の本質に迫ることを目標としています。*1それはそうそう成功しないのだけれど、ガヤ論文とNewsome論文に関してはかなり迫ることができたのではないでしょうか。そういう意味では私がやっていることは日々の生活雑記ではないし(実名でやっている以上、それは抵抗があります)、あんまりweb日記、という感じではないのかもしれません。「日誌」ですな。それが私がconfidential matterを守りつつも有意義のあるやり取りを交わすにはどうすればよいかというひとつの案なのです。
そうやって考えれば私がやろうとしていることはエフェクター型のうちのエンハンサー型(「よそのネタ(=online publication)の考察を深める」)だし、こまめに日々出版される論文にまんべんなく要約をつけてゆくエフェクター型のうちのコンプレッサー型(「よそのネタを要約する」)を目標としているわけではないのですな。もちろんそれ自体は意味があるわけです。Journal clubのネタ探し用に使ってもらうならそれがよいことでしょう。そして怠けているときは単に論文へのリンクだけを列挙するサンプラー型(「よその情報をたくさん列挙する」)となることもあるわけですが、サンプラー型であるためには安定して情報提供をしなければならないわけで、そういうわけでたまにサンプラー型になるってのはあくまで自分のため、あとで深く考察する候補のひとつとして記録してゆく、という役割を果たしているのです。私のところでは。
というわけでセレクションかけた論文を深く読み込む方向に進んでいこう、と確認したのでした。「pooneilの脳科学論文考察日誌」です。あー、また偉そうなこと書いた。前口上はいいからさっさとやればよいのだけれど、こういうことを語るのも大好きなのです。
*1:もちろん可能ならば関連論文に言及してさらに議論を一般化させることによってその分野がどこまで進み、どこまでいけるのか、そこまでいければよい、と思っています。そこまではまだ到達できていないし、本当はNewsome論文に関してはここから先が、neuroeconomicsの将来性、行動分析と神経科学との関係、といったところまで食い込めるチャンスなのですが、息切れしております。
■ clustering
そうか、linear discriminant analysisとmultidimensional scalingとはsupervisedであるか否かの点で別物か。そりゃそうだった(情報提供from ガヤ)。というわけでメモメモ。
2004年07月22日
■ Psychometric functionの書き方
 やっとわかった。こんなこともわからないでいたのです。Psychophysicsの基本中の基本にもかかわらず。
やっとわかった。こんなこともわからないでいたのです。Psychophysicsの基本中の基本にもかかわらず。
Psychophysicsではdetectionの閾値を決定するのにpsychometric functionを書きます。たとえば図のようなものです。たとえば、二つのターゲットのどちらかが点灯するのでどっちが点灯しているか当てなければならないとします。横軸はターゲットの明るさで、右が明るくて左が暗くなります。縦軸は正答率です。ターゲットのコントラストが明るいときには正答率はほぼ1ですが(右側の青丸)、暗くなるとどっちが点灯しているかわからなくなるので正答率はchance levelの0.5まで落ちます(左側の青丸)。これをlogistic regressionによってfittingしてやって、縦軸の0.75を通るところをその人の検出能の閾値としてやるわけです。しかしlogistic regressionというのは普通は縦軸が0-1の範囲を動くものなので、二択の時にはそれを0.5-1の範囲を動くようにmodifyしてやらなければならないのですが、そのやり方がよくわからなかったのです。
そこで参考サイト:"Matlab and Sas logistic"
というわけで、matlabでglmfitを使い、inline関数でfittingしたい関数の形を設定してやればよかったのです。
そこでまず、inline関数で2択(2AFC: two alternative forced choice)、四択(4AFC: four alternative forced choice)、yes-no question (正答率0-1の範囲)のそれぞれの場合のlink関数を設定してやります。
function mylink = choose_link(choice)
% choose link function
% 0:yes-no 0-1;
% 2: 2AFC 0.5-1;
% 4: 4AFC 0.25-1
switch(choice)
case 1,
mylink='logit';
case 2,
fl=inline('log( (max(0.5+1e-6, min(p, 1-1e-6))-0.5) ./ ...
(1-max(0.5+1e-6, min(p, 1-1e-6))))')
fd=inline('(1-0.5) ./ ( (1-max(0.5+1e-6, min(p, 1-1e-6))) .* ...
(max(0.5+1e-6, min(p, 1-1e-6))-0.5))')
fi=inline('(0.5 + exp(x)) ./ (1+exp(x))')
mylink={fl fd fi};
case 4,
fl=inline('log( (max(0.25+1e-6, min(p, 1-1e-6))-0.25) ./ ...
(1-max(0.25+1e-6, min(p, 1-1e-6))))')
fd=inline('(1-0.25) ./ ( (1-max(0.25+1e-6, min(p, 1-1e-6))) .* ...
(max(0.25+1e-6, min(p, 1-1e-6))-0.25))')
fi=inline('(0.25 + exp(x)) ./ (1+exp(x))')
mylink={fl fd fi};
end
これをchoose_link.mとして保存しておいて、
dat=[0.6021 4 9
0.6990 12 25
0.7782 8 14
0.8451 14 16
0.9031 11 11
1.0000 9 9
1.0792 11 11
1.1761 10 10];
% [x y Ny] variable success total
x =dat(:,1);
y =dat(:,2);
Ny=dat(:,3);
mylink = choose_link(2);
[b,dev,stats]=glmfit(x,[y Ny],'binomial',mylink);
xx=min(x):(max(x)-min(x))/100:max(x);
yfit = glmval(b,xx,mylink);
figure
hold on
h1=plot(x,y./Ny,'o');
h2=plot(xx,yfit);
set(h1,'markersize',10)
set(h1,'linewidth',2)
set(h2,'linewidth',2)
データ(dat)からlogistic regressionしてやると図のようなpsychometric functionが書けるという訳でした。
もう少し説明しますと、入力するデータdatとして、一列目がvariable (横軸として振った値)、二列目がsuccess trial数、三列目がtotal trial数という行列を用意します。これをglmfitに入れるわけですが、logistic regressionでは四番目の引数にリンク関数を入れる必要があります。これが'logit'なのが普通の0-1の範囲を動くデータでのlogistic regressionです。そこでchoose_link functionでこのリンク関数をinline関数mylinkとして設定してやるとregressionができて、coefficient bが出てくるのでこれでデータの範囲を100等分した新しい横軸のデータでglmvalを使ってfittingの値を得る、というわけでした。
何とかここまで来れましたが、要するにわたし、inline関数とかeval関数とかのような文字列を評価して計算する、というのがいまだによくわかってないのですね。
glmfitのhelpにたしかに関連することが書いてあるのですが、結局のところ、どのような関数を設定するかは人のサイトのほぼ丸写しでした。うーむ、敗北。
なお、参考ページを見ていただければおわかりのとおり、psychometric functionを書くためにはいくつかのやり方があります。たとえば、今回使ったようなprobit関数 (log(p/(1-p))によるのではなくて、gaussian分布の累積確率密度関数を使ってやるとか。というわけで細かく言うときりないようなのでこのあたりにて。
# ryasuda
データを(P-0.5)/0.5と0-1になるようにnormalizeするのもよくみかけます。
# pooneilなるほど、それはシンプルですね。Lest squareではなくてlogistic regressionのときに計算的に妥当なのかどうかはよくわからないのだけれど。
2004年07月21日
■ はてなアンテナのリンクはスケールフリーネットワークだった。
 ガヤ、サンクス。なるほど、scale-free(3/17,3/31,4/6にて言及あり)の話でしたねこれは。というわけで7/20の図1を確率密度分布に変換して(各月の総リンク数で縦軸を割る)、縦軸横軸を両方ともlogスケールにしたものが今日の図3の左です。横軸はあるサイトがアンテナに登録されている数k、縦軸は被リンク数kのサイトの全サイトに占める割合P(k)です*1。7月分重ねてありますがどれも同じ傾向ですので色分けしておりません。きれいな直線的関係(power law)を示しているのがわかります。
ガヤ、サンクス。なるほど、scale-free(3/17,3/31,4/6にて言及あり)の話でしたねこれは。というわけで7/20の図1を確率密度分布に変換して(各月の総リンク数で縦軸を割る)、縦軸横軸を両方ともlogスケールにしたものが今日の図3の左です。横軸はあるサイトがアンテナに登録されている数k、縦軸は被リンク数kのサイトの全サイトに占める割合P(k)です*1。7月分重ねてありますがどれも同じ傾向ですので色分けしておりません。きれいな直線的関係(power law)を示しているのがわかります。
ここで解説を。WWWのようなネットワークの個々の頂点vertexが他のvertexと何個繋がっているか(k)の確率密度分布(P(k))は以下のようなpower lawによって説明できることがわかっています。
P(k) ~ k-γ log10(P(k)) = log10(k) - γ (実際にはintersectの項が入るが、WWWでは0になるらしい)
参考文献:Science '99 "Emergence of Scaling in Random Networks."
なお、このpower lawはネットワークが(1)元からあるネットワークに新しい結合が付加されてゆく、(2)新しい結合はランダムに付加されるのではなくて、よりkの大きいところにより多く新しい結合ができる("rich-get-richer phenomenon")、という二つの性質を持っているときに成り立ち、ランダムな結合では成り立ちません。ですので、ryasudaさん、リンクの分布がpoisson-likeであると書いたのは私の早とちりでした。全員がランダムにリンクしたらポワソン分布になるのはその通りだと思います。実際にpoisson分布だったらどんなプロットになるか作ってみたのが図の右側です*2。縦軸横軸は左図と同じです。実際のはてなアンテナのデータではλ=3(kの期待値=2.8より)ですので、それでプロットしたのが赤い線、λを10.^(-10:0.1:1)でふってみたのが青線です。どのようなλであれ、poisson分布の時にはk>100であるような頂点はほとんどありえません(P(k=100) < 10^(-100))。k>100のようなサイトは"rich-get-richer phenomenon"によって形成されるのであって、偶然にはほとんど作りえないということがわかります*3。この二つの対比(左のpower lawと右のpoisson分布)こそがはてなアンテナの被リンク数が上記の(1)(2)の性質を持っていることの証拠となるでしょう。
左の図に戻ります。1月から7月までのそれぞれの月のデータはほとんど同じパターンを作っており、これはガヤが指摘するようなscale-freeであることの証拠といえるのかもしれません。また、各月のデータでlinear regressionして得たslopeからこのネットワークのγを推定することができますが、intersect=0の条件(横軸縦軸ともに10^0を通る)の元で、2.10±0.02 (mean±SD)となります。これは上記のScience '99でのWWWの例でのγが2.1±0.1であるのと劇的な一致を示します。実のところ、はてなアンテナの被リンク数とは、WWWのリンク数のような双方向的なものではなくて、はてなダイアリーを使ってないけどはてなアンテナだけは使っているという人もいるのにもかかわらず。自分で計算しておいて驚いてみたり。
*1:k=0のデータは除外して計算しております。
*2:ネットワークでは結合のない頂点はないものと見なされますので、結合数k=0のものは除いて、1<=k<∞でΣP(k)=1となるようにnormalizeし直してあります。
*3:なお、Watts and StrogatzのNature '98でのsmall-world networkはlocalに繋がったランダムなネットワークの結合のある一定の割合を遠い場所につなぎ直したものですので、この場合P(k)の分布はランダム結合の場合と同じであり、scale-free power lawは成り立たない、ということがScience '99に書かれています。Scale-freeとsmall-worldは似ているようで微妙に違ったものであるようです。
- / ツイートする
- / 投稿日: 2004年07月21日
- / カテゴリー: [ネットワークの科学(small-world, scale-free)]
- / Edit(管理者用)
# ガヤ
いま実験の待ち時間中。いや、これまたやってくれますね。しかもきれいな図になりました。K=1を見ていなかったので、もしかしたらSingle-Scaleになるかと思いましたが見事です。ところで、あまり詳しく覚えていませんが、上記のScience論文では“rich-get-richer”の部分で、Linearモデルを使っていた(つまりK=100はK=1よりも100倍の「客寄せ効果」がある、といういう単比例で計算していた)と思うのですが、この仮定がおそらく「正しくない」だろうということは、今回の図でもプロットが末広がりになっている(しかも、Kが大きいほど予想される直線よりも右方向にバラけている)ことから想像できます。つまり、K=100はK=1よりも100倍以上は魅力的なサイトだってことですね。この方が我々の直感に合っているかと思うわけです。
# ryasudaなるほど、面白いですね。Journal citationとかも似たような感じなのかな。
# ガヤryasudaさん、さすが鋭いですね。PNAS 97(2000)11149-11152には「the network of citations of scientific papers are scale-freethat is, they have a distribution of connectivities that decays with a power law tail」という記述がありますよ。
# pooneilなるほど、「予想される直線よりも右方向にバラけている」、これ面白いですね。まず、この辺の値の全体に占める割合は非常に小さいため(< 10^(-4))、どうしてもバラけるのは本当でしょうね。で、ガヤの言う通りに線形性がないとするとプロットは右曲がりになるというわけですね。ちょっと誤差が大きすぎて本当かどうかわからないけれど、その右方向にバラけているやつはじつは主にid:hatenadiary、つまり<A HREF=”http://d.hatena.ne.jp/hatenadiary/”>「はてなスタッフのはてなダイアリーに関する記述」</A>なのです。よってそのような特殊かつ中心的なステータスにあるサイト(しかもネットワーク形成の一番最初からあったっぽい)がネットワークの中でどんな動態を示すのか、というふうに捉えられるのかもしれません。ほかにもScale-freeの例として俳優の共演ネットワークがScience ’99には挙げられていました。これはsmall worldの方でも挙げられている例です。そうなると、俳優の共演ネットワークはscale-freeなのか、small-worldなのか、それとも両立しうるのか。Scale-freeになるような(1)と(2)の性質を持って自己組織化したネットワークはsmall-worldなのか、というあたりに興味が出てきます。どっかですでに扱われているのでしょうけど。
2004年07月20日
■ "はてなアンテナ被登録数"
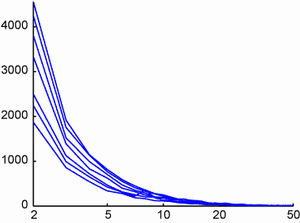
Figure 1
"はてなアンテナ被登録数"の統計が出ていたのでヒストグラムを書いてみました(図1)。横軸ははてなアンテナ被登録数(logスケール。0と1はomitしてあります)で、縦軸はそれぞれの横軸に該当するidの数です。それぞれのプロットは1月から7月までの毎月の計測データで、色分けしてませんが、だんだん増える方向にあります。注目する点としては、分布の形自体は変化がないのですね。
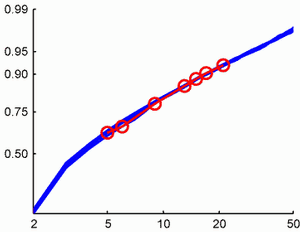
Figure 2
というわけでこれをcumulative plotにしたのが図2です。横軸は図1と同じで0と1は計算から除外しています。縦軸Pは累積データの全体のデータに占める割合です。スケールはprobitスケール、つまりP-valueをZ valueに変換したものです(MATLABでいうところのnorminv(P,0,1))。もし分布がgaussianならこのプロットは直線になります。じっさいにはpoisson-likeですが。青線は各月のデータですが、ほとんど重なっていることがわかります。赤丸を赤線でつないだid:pooneilは順調に被登録数を上げていき、7月現在で上位10%まで来てます。専門的なサイトにしては健闘していると言えるでしょう。
# ガヤ
おっと?これは思わず書き込みせずにはいられない統計処理(笑) 0と1が省略されているのでなんとも言えませんが、2エッジ以上を見る限り期待通りスケール・フリーっぽいですね。各月の折れ線が重なっているのにも、その特徴が現れているかと。
# ryasudaPoissonになるってのは面白いですね。全員がランダムにリンクしたとしても、ポアソン的になりそう。
# pooneil>>ガヤ なるほど、scale-freeの話でしたねこれは。というわけで7/21のところでやってみました。けっこうきれいにいきました。>>ryasudaさん、はじめまして。日記見にいってます。アメリカでラボを持つことに関する貴重な資料だと思っております。これからもよろしくお願いします。それから、分布がpoissonだというのは私の早とちりでした。全員がランダムにリンクしたときには今回の分布のような100以上のリンクを持つサイトができることはない、というあたりも7/21で検証してみました。よければご覧ください。
- / ツイートする
- / 投稿日: 2004年07月20日
- / カテゴリー: [Paper archive]
- / Edit(管理者用)
■ JNS 5/26
"Encoding-Specific Effects of Social Cognition on the Neural Correlates of Subsequent Memory."
- / ツイートする
- / 投稿日: 2004年07月20日
- / カテゴリー: [Paper archive]
- / Edit(管理者用)
■ PNAS 5/25
"Dynamic mapping of human cortical development during childhood through early adulthood."
- / ツイートする
- / 投稿日: 2004年07月20日
- / カテゴリー: [Paper archive]
- / Edit(管理者用)
2004年07月19日
■ Spatial hemineglectの原因部位
Spatial hemineglectは主に右半球のparietal cortexの損傷で起こり、左視野を無視するようになります。左半球の損傷の例は少なく、障害の程度も少ないことがわかっています(右脳左脳なんて言いますが、これこそが人間の脳の左右差を示す一番はっきりしたものであると言えます)。一番印象的なのは、時計の絵を患者さんに模写してもらうと右半分だけを書いた絵になる、というものでしょう。
んでもってその原因部位は右半球のparietal cortexのうちのIFG (inferior frontal gyrus)からTPJ(temporoparietal junction)のあたりということでだいたい考えられていたのですが(前述のJon DriverのNature Neuroscienceの図1を参照)、2001年になってUniversity of TübingenのHans-Otto KarnathがNatureに
"Spatial awareness is a function of the temporal not the posterior parietal lobe."
を出しました。Karnathはこれまでの研究ではneglectのある患者さんにはhemianopia (optic radiationの損傷による視野欠損:blindsightの原因になるやつです)も併発している例が多い(ある例では80%)ことに注目し、hemianopiaのない、純粋なneglectの患者さんのデータだけにしぼってその損傷部位を調べると、じつは一番頻繁に損傷が見られたのは前述のIPL、TPJではなくて、そのすぐventral側のSTG(superior temporal gyrus)であることがわかったというものです。しかも確認のため、neglectとhemianopiaの両方の症状がある患者さんの損傷部位を調べると、これまでの報告と同様、IPLとTPJでの損傷が多く見られたのです。損傷していた右のSTGの反対測、左のSTGはいわゆる言語野であって、そのためKarnathはnon-human primateとhumanとに分かれる進化の過程で左STGが言語野としての専門化することによって右STGが注意をつかさどる領域として専門化した、というspeculationをするのです。
一方、これはかなりcontrovercyを惹き起こしたようで、これに対する反論などがCortex誌上で行われたらしいのですが私のところからは読めません。その代わり、私が知るかぎりでは、NeuroReport '03において、
"The anatomy of neglect without hemianopia: a key role for parietal-frontal disconnection?"
という論文が出版されてます。ここで著者らは、KarnathのNature論文での損傷部位は急性期(出血などが起こった直後)のものを見ている点に注目して、慢性期に入った患者さんのうちでKarnath論文と同様、neglectのみがあってhemianopiaがない患者さんの損傷部位を調べました。すると一番頻繁に見られた損傷部位はKarnath論文のようなSTGではなくて、SMG(supramarginal gyrus、IPLの一部)だったのです。というわけで著者らはKarnath論文の結論は一部言い過ぎであり、気をつけて扱うべきである、としています。
一方で、KarnathはCerebral CortexのAOPにおいて、
"The Anatomy of Spatial Neglect based on Voxelwise Statistical Analysis: A Study of 140 Patients." Hans-Otto Karnath
というのを出版しており、140人の患者さんの損傷部位の解析から元のNature論文と同様な結論を導いています。なお、このCerebral Cortex論文はNeuroreport論文をreferしておらず、acute、chronicの問題には触れていなさそうです。
- / ツイートする
- / 投稿日: 2004年07月19日
- / カテゴリー: [半側空間無視(Spatial hemineglect)]
- / Edit(管理者用)
2004年07月17日
■ 青山ブックセンターが倒産
なんてこった。私の青春の一ページですよあそこは。渋谷でのバイトでの帰りに自転車で寄ってあそこでサブカル系の漫画やらニューエイジ系の本などを読み漁ったものです。背伸びして洋書を買い込んだり。建築系とかのでかい写真集立ち読みしたり。夜想とかバロウズとかそういうものに出会える唯一のところでした。ベイトソンの「天使の恐れ」を買ったのもあそこではなかっただろうか。今にして思えば、ヴィレッジヴァンガードほど旧宝島文化くさくはなかったし、丸善ほどはおっさん臭くなかったし、リブロとも違う、今でもほかにない存在でありつづけたのですよね。そして明け方までの営業時間。渋谷で飲んだ帰り、自転車こいで西麻布とか六本木とかの町に目もくれずABCに寄って、ふらふらしながら立ち読みして、なんも買わず、もしくはやっすい雑誌をなんか買って、そんでそこから赤坂、銀座を経由して永代橋を越えて帰っていってたんです。若いなあ。それはまだBOOKOFFもTSUTAYAもないころの話でしかないのだけれど。
■ Annual Review of Neuroscience
"NEURONAL CIRCUITS OF THE NEOCORTEX." Rodney Douglas and Kevan Martin。
Douglas and Martinと言えば、1991年のJournal of Physiology四連報ですな。
- "An intracellular analysis of the visual responses of neurones in cat visual cortex."
- "Mechanisms of inhibition in cat visual cortex."
- "Excitation by geniculocortical synapses is not 'vetoed' at the level of dendritic spines in cat visual cortex.."
- "A functional microcircuit for cat visual cortex."
そののちで私の興味は多電極記録へと移行しましたが、スパイクを見るだけではなくてsynapticなイベントを見なければ、ニューロンの入力(PSP)から出力(スパイク)を変換する過程が見れていない、という意味での重要性がまったく薄れていないのは確かです。
In vivoでのintraというアプローチはのちにFregnacのNatureでの刺激応答時のshunting inhibitionの発見、さらにDavid Fersterによる膜電位のup- down-stateの発見などへとつながってゆきます。また、1991年以降にはoptical imagingによるV1のfuncitonal organizationの解明が劇的に進みました。このannual reviewでどのくらいそのへんが盛り込まれて進歩しているか、というのに興味があります。
# ガヤ
関係ないですがV1について。KonigがCurrOpinNeurobiolで何やら書いているようですよ。読んでませんがAOPで出てますね。
# pooneilおひさしぶり。königのは8月号のSensory systemsの号の原稿のようですね。リンク付けときました。ありがとうございます。
■ Annual Review of Neuroscience
- "THE MIRROR-NEURON SYSTEM." Giacomo Rizzolatti and Laila Craighero
- "THE MEDIAL TEMPORAL LOBE." Larry R. Squire, Craig E.L. Stark, and Robert E. Clark
- "CORTICAL NEURAL PROSTHETICS." Andrew B. Schwartz。今まで彼が使っていたdecodingのアルゴリズムはたんなるpopulation vectorだったんだけど、ちらっと見たらカルマンフィルター、なんて書いてある。少しは改良しようという意志はあるらしい。
- "ATTENTIONAL MODULATION OF VISUAL PROCESSING." John H. Reynolds and Leonardo Chelazzi。Desimone傘下がV4ニューロンのattentional modulationについて書いている模様。
- / ツイートする
- / 投稿日: 2004年07月17日
- / カテゴリー: [Paper archive]
- / Edit(管理者用)
■ Current Opinion in Neurobiology Article in Press
ガヤのコメントにより追記。
"Processing of complex stimuli and natural scenes in the visual cortex." Peter König。
Current Opinion in Neurobiologyは
- 2月号 Development
- 4月号 Cognitive neuroscience
- 6月号 Signalling mechanisms
- 8月号 Sensory systems
- 10月号 Neuronal and glial cell biology
- 12月号 Motor systems
Natural sceneとV1という関連では
あたりが重要論文でしょう。
- / ツイートする
- / 投稿日: 2004年07月17日
- / カテゴリー: [Paper archive]
- / Edit(管理者用)
2004年07月16日
■ コメントしてくだった皆様へ
いつもコメントありがとうございます。うまく繋がったスレッドに関してはniftyの方のホームページに編集したものを掲載していますが、これまでのものについてはコメントを掲載する許可をいただいていなかったのでコメントは掲載していませんでした。もしよければ今回のNewsome論文およびMilner論文以降はみなさまのコメントも掲載させていただきたいと思いますがよろしいでしょうか。それ以外に転載したくなった場合には改めてお願いをしますので。なお、この件については0000年00月01日の注意書きのところにも記載を入れました。
■ Science Newsome論文
放置してましたがまだ続けるつもりはあります。とくにmmrlさんが送ってくださったエントリ(6/31)およびJCについての報告に関しては忘れずやります。
- / ツイートする
- / 投稿日: 2004年07月16日
- / カテゴリー: [Paper archive]
- / Edit(管理者用)
- / ツイートする
- / 投稿日: 2004年07月16日
- / カテゴリー: [Paper archive]
- / Edit(管理者用)
■ Nature Neuroscience 7月号
"Automatic avoidance of obstacles is a dorsal stream function: evidence from optic ataxia."
つづき。
Optic ataxiaに関する重要な論文にGoodale and Milnerの
Nature '91 "A neurological dissociation between perceiving objects and grasping them."
があります。以前にも言及しましたが、visual agnosia (視覚認知ができない)の患者さんであるDFさんは目の前のスリットの角度がわからないにもかかわらず、このスリットにカードを差し込むことはできます。一方、optic ataxiaの患者さんでは話は逆で、スリットの角度はわかるのに、そのスリットにカードを差し込むことができない、というわけです。
つまり、視覚意識がないのに視覚運動変換ができる場合と視覚意識があるのに視覚運動変換ができない場合というdouble dissociationを示したわけです。さらに彼らは"Visual brain in action"(MIT press)においてその二つがdorsal pathwayとventral pathwayとに分かれている、といった説を展開することになります。
今回の論文の著者であるRossetti and Vighettoは最近EBRにoptic ataxiaに関するreviewを書いていて、上記のGoodale and Milnerの説を批判しています。
"Optic ataxia revisited: Visually guided action versus immediate visuomotor control." Yves Rossetti, Laure Pisella and Alain Vighetto
つまり、前述のとおり、optic ataxiaでは注視点へ手を伸ばすのには問題がないわけで、optic ataxiaの場合のテストは周辺視野で行っており、中心視野で行っていない。一方でvisual agnosiaの場合のテストは中心視野で行っており、周辺視野で行っていない。よって、二つの症状を比較するためにはどの視野でのことなのかをそろえて検証すべきだ、というわけです。んで、このレビューが出た後の今回のNature NeuroscienceにはMilnerが入っている、というわけなんですな。
なお、Rossetti and Vighettoの代表作は
"Optic ataxia: a specific disruption in visuomotor mechanisms. I. Different aspects of the deficit in reaching for objects." Perenin MT, Vighetto A
こいつです。Brainなんですが、Iとか書いてあって連報のように見えて、続報が出てないのです。かっこ悪い。内容は別として。
- / ツイートする
- / 投稿日: 2004年07月16日
- / カテゴリー: [腹側視覚路と背側視覚路]
- / Edit(管理者用)
2004年07月15日
■ JNS 7/14
- "Target Selection in Area V4 during a Multidimensional Visual Search Task." 小松研の小川さんのbottom-up / top-down attention taskが掲載されてる。おめでとうございます。
- "Temporal Dynamics of Neural Adaptation Effect in the Human Visual Ventral Stream." こちらもあるので今回のJNSのBEHAVIORAL/SYSTEMS/COGNITIVEセクションは生理研祭りですな。
- "Independent Coding of Reward Magnitude and Valence in the Human Brain." rewardのmagnitude (small or large) and valence (win or loss)がERPのP300とfeedback negativityとでdouble dissociationする。追記:mmmmさんによるコメントがあります。それからCamille et al. (2004) Science 304:1167-1170は5/22にリンクした"The Involvement of the Orbitofrontal Cortex in the Experience of Regret."です。
- / ツイートする
- / 投稿日: 2004年07月15日
- / カテゴリー: [Paper archive]
- / Edit(管理者用)
■ Nature Neuroscience 7月号
"Automatic avoidance of obstacles is a dorsal stream function: evidence from optic ataxia."
つづき。
じつは著者らはほぼ同じタスクを使ってspatial hemineglectの患者さんで同じテストをしています。spatial hemineglect(半側空間無視)は主に右のPPCの損傷によって左視野半分への注意が向かなくなる、というもので、基本的に視覚注意の障害です。
"Preserved obstacle avoidance during reaching in patients with left visual neglect." Neuropsychologia '04
こちらでは結果はまったく逆で、円柱のあいだを手を伸ばすときには、円柱の位置によって手の軌道に変化を受けます。つまり健常者とまったく同じで、automatic avoidanceは可能です。しかし、円柱の中間を示すタスクではちょうど真ん中を指すことが出来ません。これは左側の円柱が無視される影響で中心より右側を指すということが起こるためです(じつは結果はしょぼいのだけれど)。
つまり、この二つの論文の結果を合わせると、double dissociationが起こっていることがわかります。つまり、optic ataxiaもhemineglectもおなじPPCの障害によって起こる症状であるにもかかわらず、optic ataxiaの方ではautomatic avoidanceの機能のみに、hemineglectの方では視覚注意の機能のみに障害が起こり、別の機能には障害がありません。つまり、optic ataxiaとhemineglectとで傷害される部分は別の独立した機能を持っていることがわかるのです。
それではじっさいの障害部位はどこか、といいますと、
"Two different streams form the dorsal visual system: anatomy and functions." Giacomo Rizzolatti and Massimo Matelli
によりますと、上のほう(SPL)が傷つくとoptic ataxiaになって、下のほう(IPL)が傷つくとhemineglectになる、ということのようです。じっさいには単純にSPLとIPLとに分けられるかどうか疑問でして、optic ataxiaの方がparietooccipitalである、というように書いている場合もあるようですが。なんにしろ、Rizzolattiはそれまでのanatomyなどの結果から、PPCには二つのpathway、SPLを通る経路とIPLを通る経路とがある、ということを提唱しています。(じつは、ventral pathwayでも単一の経路があるのではなくて、STSの中を通るような別の経路がある、という話があるんですが、それはまたいつか。)しかし、Rizzolattiはいろんなことやってますな。
なお、PPCのもう一つ重要な機能としてbody schemaの形成というものがありますが、これも二つの症状で現れ方が違っておりまして、optic ataxiaの患者さんの多くは、自分の体を指差すときには不思議と問題がなかったりします。一方、hemineglectの患者さんはしばしば空間を無視するだけでなく、自分の体の半分も無視するという現象が起こります。つまり、IPLのほうがおそらくはbody schemaの形成に関わっているということでしょう。(>>Correggioさん、合ってますか?)
明日つづきを貼ります。これで最後。Goodale and Milnerに関連して。
追記:Rizzolatti and Matelliでoptic ataxiaとneglectについてどう書いてあるか貼っときます。
Lesions centered on SPL determine optical ataxia (Ratcliff and Davies-Jones 1972; Perenin and Vighetto 1988). Lesions of the right IPL, especially its lower part, produce neglect (Vallar and Perani 1987; Perenin and Vighetto 1988). (中略) These data suggest a clear dichotomy between the functional roles of SPL and IPL. SPL is involved in action organization. In contrast, IPL is deeply involved in perception, but it is also involved in the organization of motor activities.
というわけでSPLとIPLという言葉はhumanのものですので、ご隠居の誤解ではありません。Correggioさんも書いているようにRozzolattiは相同に関しては回避している模様です。
- / ツイートする
- / 投稿日: 2004年07月15日
- / カテゴリー: [腹側視覚路と背側視覚路]
- / Edit(管理者用)
# Correggio
じつは、そういう風にきれいに行けばいいのですが。。。。実は、Rizzolatti等の主張とGalletti等の主張の根拠は基本的にはそんなに違っていません。解剖学的に、確かに二つの経路(正確には3つか)に分ける。つまり、V6-V6A-MIP-PMd のV6-VIP-F4の背側背側経路とCIP-AIP-F5(実はもう一つPF-F5こっちはミラーニューロン)の背側外側経路です。大まかにいえば、到達運動に関わるのが背側内側経路、背側内側経路は把握運動というふうに考えられますが、Gallettiたちは背側内側経路も把握運動に関わっているといっています。ところで、ヒトSPLとIPLはサルのどこにあたるのかははっきりしませんが(Rizzolatti等もはっきり言ってない感じ)とりあえず背側外側のMIP経由はSPLをとおり、背側内側経路はIPLを通るとして、でも、こうするとVIPの経路はどっちにはいるのかなあ(一応RizzzolattiたちはIPLにいれてるみたい)。で、V6-V6Aの経路が、optic ataxiaに、そして、VIPの経路が空間無視に、AIPの経路は、失行症に関わると考えられます。身体感覚は、どちらともいえないようです。Shiriguたちの1999年の論文(Brain)では、自己と他者の手の視覚イメージが、どちらに属するのか区別できない症例の中に39野と40野の傷害の例を挙げています。一方、Wolpertの1998年のNature neuroscienceでは、SPLで、身体感覚の障害を示す患者の例を紹介しているのです。確かに、ニューロンの記録では、入來さんやGrazianoはSPLですし、Hyvaerinenの記録は、PFですね。
# pooneil解説ありがとうございます。うーむ、そんなに簡単ではないようですね。GallettiのEBRのイントロを読んでみました。SPLからPMdへ行くのがreachingで、IPLからPMvへいくのがgrasingというパラレルなstreamがある、という考え自体は以前から提唱されていて、しかもそれがすでに批判にさらされている、ということからすると、GallettiのもRizzolattiのもそれのmodificationみたいな立場にあるようですね。身体感覚がIPLってのはかなり納得がいってたのですが、なるほどうーむ、もちろんSPLとIPLとのあいだでの情報伝達もあるんでしょうけど、不思議な話ですね。SPLの方がおそらくsomatosensoryとの関連が強いだろうし、IPLの方がミラーニューロンのシステムとの関連は強いでしょう。よって身体図式といっても、違った要素からできているということなのかもしれません。
# mmmmYeung&Sanfey(reward magnitudeとvalenceとの分離の論文)を一瞥しただけの茶々です(並立スレッドを立てると混乱しそうですが)。課題も面白いし、Fig. 2はかなり美しいですね。本当はregret(あるいはfactual vs counterfactual)も含めてtriple dissociationを狙ったんでしょうね。ところがこの三番目のfactorでmedial frontalを狙ったのは失敗だったのかもしれないですね。この論文ではbehavioral adjustmentと関連づけるという間接的な役回りを果たしているようです。Camille et al. (2004) Science 304:1167-1170がfMRIを使ってOrbitofrontalで出し抜いた形になってしまいました。
# pooneilmmmmさん、ありがとうございます。なるほど、そのように読めるのですね。とっかりがなくてどう捉えたらよいかわからなかったのですが。Camille et al. Science ’04は5/22にリンクしておいたものですね。もういちどリンクし付けときます。
# pooneilCorreggioさんから上のコメントの清書バージョンをいただきましたので、ここに貼っておきます。あとで編集するときに忘れないように。Correggioさん> じつは、そういう風にきれいに行けばいいのですが。。。。Rizzolatti等の主張とGalletti等の主張の根拠は基本的にはそんなに違っていません。解剖学的に、確かに二つの経路(正確には3つか)に分けられる。つまり、V6-V6A-MIP-PMd のV6-VIP-F4の背側内側経路とCIP-AIP-F5(実はもう一つPF-F5こっちはミラーニューロン)の背側外側経路です。大まかにいえば、到達運動に関わるのが背側内側経路、把握運動は背側外側経路というふうに考えられますが、Gallettiたちは背側内側経路も把握運動に関わっているといっています。ところで、ヒトSPLとIPLはサルのどこにあたるのかははっきりしませんが(Rizzolatti等もはっきり言ってない感じ)とりあえず背側内側経路のMIP経由はSPLをとおり、背側外側経路はIPLを通るとして、こうするとVIPの経路はどっちにはいるのか。(一応RizzzolattiたちはIPLにいれている)。で、V6-V6Aの経路が、optic ataxiaに、そして、VIPの経路が空間無視に、AIPの経路は、失行症に関わると考えられます。身体感覚は、どちらともいえないようです。Shiriguたちの1999年の論文(Brain)では、自己と他者の手の視覚イメージが、どちらに属するのか区別できない症例を報告し、39野と40野の傷害の例を挙げています。一方、Wolpertの1998年のNature neuroscienceでは、SPLで、身体感覚の障害を示す患者の例を紹介しているのです。確かに、ニューロンの記録では、入來さんやGrazianoはSPLですし、Hyvaerinenの記録は、PFですね。
2004年07月14日
■ Nature Neuroscience 7月号
"Automatic avoidance of obstacles is a dorsal stream function: evidence from optic ataxia." "The visual brain in action"をGoodaleと書いたDavid Milnerです。それからoptic ataxia関連の論文を書いてきたRossetti and Vighetto。Posterior parietal cortex (PPC)の障害でoptic ataxiaの症状がある患者さん二人へのテストの結果です。
Optic ataxiaとはなにかというと、視覚は正常で、物がどこにあり、何なのかもわかっているのにもかかわらず、それを手を伸ばして掴むことができない、という症状のことです。後述のRizzolattiのEBRによれば、"a disorder of visually guided movements of the arms toward a goal"です。この障害は、その物体が視野のはしっこにあるときにより厳しく、物体を視野の真ん中で見ている(foviate)ときには問題にならなくなります。
んでもって実験パラダイムはFig.1の通りですが、テーブルの向こう側に注視点があってそこを注視しながら手を伸ばして注視点に触れることが要求されています。手を伸ばす途中には円柱が二本、手が通るあたりから10cm横の左右に置いてあります。この円柱は手を伸ばすときに当たることは決してありません。しかしこの円柱の位置を左右に4cmほどずらしてやると、このタスクをコントロールの健常者がやるときには手の軌道がこの円柱を避けるようにわずかに逸れます。被験者はこのような微妙な調節を意識してやっているわけではありませんので、これは"automatic avoidance"です(円柱が手の軌道に近いところに置かれているときには意識してそれを避けて手を伸ばすので、automaticではありません)。しかし、このタスクをoptic ataxiaの患者さんにやってもらうと、そのような手の位置の調節がなく、円柱の位置によらずいつも同じ手の軌道を描くことになります。*1 つまり、optic ataxiaでは、視覚誘導性のreachingの障害だけではなくて、このようなautomatic avoidanceにも障害があるということがこの論文でわかったのです。なお、視覚には障害がないということを示すためのコントロールとして、別のタスクでこの二つの円柱の中間を指差すように、というタスクをすると、これはまったく問題なくできます。よって、円柱の位置の変化を認識していないということではないのです。
明日つづきを貼ります。
追記:Correggioさん指摘のGallettiらのEBRのreview。
Experimental Brain Research '03 "Role of the medial parieto-occipital cortex in the control of reaching and grasping movements." Claudio Galletti , Dieter F. Kutz, Michela Gamberini, Rossella Breveglieri and Patrizia Fattori
追記:ご隠居のコメント関連。RizzolattiのはBrain '83 "Deficits in attention and movement following the removal of postarcuate (area 6) and prearcuate (area 8) cortex in macaque monkeys."、HeilmanのはArch Neurol '94 "Posterior neocortical systems subserving awareness and neglect. Neglect associated with superior temporal sulcus but not area 7 lesions."ということで合ってますか? ほかにメジャーな論文としてはGaffanのBrain '97くらいではないでしょうか。この2、3年でneglectの動物モデルでいいjournalに載ったのはないように思います。マーモセットの論文(Behavioural Brain Research '02)をひとつ見つけましたが。んでもって、neglectの種差の問題の参考文献はNature '01 "Spatial awareness is a function of the temporal not the posterior parietal lobe."ですね。あとご存知でしょうが、Nancy KanwisherのCurr Opin Neurobiol '01 "Neuroimaging of cognitive functions in human parietal cortex."はPPCの種間の相同の議論をするのに役立ちそうですね。
*1:なお、前述したとおり、optic ataxiaがあっても、注視しているところへ手を伸ばすことは難しくないそうです。
- / ツイートする
- / 投稿日: 2004年07月14日
- / カテゴリー: [腹側視覚路と背側視覚路]
- / Edit(管理者用)
# Correggio
この論文まだちゃんと読んでいませんが、まだちゃんと読んでないですが、GallettiらのEBRのreviewでは、optic ataxiaがサルでいうPOに領域の傷害で起こるのではないかと主張しています。彼らはサルのPOで眼球位置に関係なくターゲットの位置をコードすると思われるニューロンを記録しているわけですが、つまり周辺視野における物体の位置情報をとらえているということでもあります。automatic avoidance の傷害の背景にはこうしたことがあるんだろうか。
# pooneilCorreggioさん、ありがとうございます。コメント、お待ちしておりました。7/15にも書きましたように私はRizzolatti and MatelliのEBRの方に準拠して書いていたので、SPLがoptic ataxiaでIPLがhemineglectと書いておいた一方で、optic ataxiaはparietooccipitalであるという書き方をしているものもあることを示しておきました。たぶん、これがGallettiのEBR ’03でのV6というやつに対応しているのでしょう。周辺視野での影響に限られている、という意味でも符牒が合うわけですね。なるほど。Rizzolatti and Matelliの方は「二つのdorsal pathway」というアイデアのために強引に解釈している節はありますが、それでもVIP-F4とAIP/PF-F5という二つのstreamの分け方は面白いと思ってます。
# ご隠居いつも大変ためになる議論を楽しく拝見させていただいております。相変わらず冬眠ぼけのまま(もう真夏だろうにっ(^-^;)ですが。古典的なはなしでは、「物体が視野のはしっこにあるときにより厳しく、物体を視野の真ん中で見ている(foviate)ときには問題にならなくな」るような障害は、ヒトでは頭頂後頭接合部の病変で起きるといわれており、(POに相当する部位も含まれているのかもしれません)一方、注視対象もつかめなくなるという症状(Balint症候群の一症状ですね)は両側頭頂後頭接合部から深部白質の病変の広範な病変で起こります。ひとによっては、前者をataxie optique、後者をoptische Ataxieなんて言い分けたりして、よけい訳がわからなくなりがちですが(まあBalintが1909年に提唱しているわけで、いろいろな歴史的経緯があるから仕方がないのでしょう)同じ用語でもどういった概念(症候)をさしているのかはちょっとだけ注意する価値があるかもしれません。病変側と程度の差なのかもしれませんが。。。ちなみに、 Balint症候群のほかの症状として、精神性注視麻痺(いったん注視した対象から他の対象へのsaccadeが起こりにくくなる)と視覚性注意障害(視野内の一つの対象を注視するとその周囲の対象が認知できなくなる)といった症状が知られています。ここまで書いて、論文をはじめてちらっと眺めてみたのですが、じっさいには頭頂葉から後頭葉の一部にかけての広範な病変がありますね。おっと、Balint症候群との記載もありました。そう、要はBalint症候群と彼らがいっているからには、その患者さんは注視対象へのreachingも障害されている(いた)はずです。また、discussionで simultagnosia(同時失認:Balint症候群でいうところの視覚性注意障害です)の影響がないといっているひとくだりは、うーん、あまり説得力がないですね。冷や汗をかいている姿が見えてきます。また新しい患者さんの例をだしていますが、そのひとの症状はさきの言葉ではataxie optiqueに相当する方で、あまり補強にはなっていません。なんか、ついつい、あたりまえの結果のようにも思ってしまうのですが、それじゃいけないのでしょう。ただ、Balintの論文、きっちりと引用してほしいですね。ついでに、個人的な古い僅かな経験では、optic ataxiaとhemispatial neglectについてはdouble dissociationを持ち出す必要性にかられたことはあまりありません。頻度的にもneglectが圧倒的に多いですし(医者くさい考え方ですみません)病変部位・症候いずれもあまり混同する機会がないように思います。もっとも、病変によってはoptic ataxiaにhemispatial neglectを合併することはあると思いますが(neglectっぽい症状は本当にいろいろな部位の病変で起こります)。そう、不勉強ですみませんが、本当にSPLの病変でoptic ataxiaが起こるのでしょうか?SPLに脳梗塞が起こることはよくあるのですが。。。高次体性感覚障害による運動障害(触覚性失認とか、また紛らわしい用語を使うひとがいますが)が起こることは時々あると思いますけど。ただ、いろいろとあたらしい観点からみると違うのかもしれません。そのまたついでですが、neglectの患者さんで、指差したところがずれていたことの原因については長年の論争があり、この論文も、それよりは障害物をよける機能は保たれていたということがいいたかったのでしょう。neglectは注意障害だと捕らえるのはちょっと単純化されてすぎていて、通常はさまざまな機能が多々障害された結果を症候群として観察していると考えたほうがよいでしょう(とくに clinical neuropsychologyの多患者研究では)最近はそれらのなかでdouble dissociationを出してpurifyする努力がされているようですが。素人のなにもよまないでのコメントですので、はずしているでしょう、すみません。さきにあやまっておきますね。ただ、ちょっとなつかしかったものでついつい。 駄文長文失礼しました。これからも楽しみにしています。
# pooneilご隠居、どうもありがとうございます。すばらしい。さすが本職。んでもって、ご隠居の書いていただいたことに特に付け加えることはありませんがなんとかもう少し書いてみます。Optic ataxiaがBalint症候群のひとつであって、simultagnosiaやocular apraxiaがこれらの患者にどのくらいあるかどうか、というのは気をつけて見ておくべき問題ですね。ご隠居のメインのメッセージは「neurologyの症状の多様さへの感度」であると捉えました。「optic ataxiaとhemispatial neglectについてはdouble dissociationを持ち出す必要性」ですが、これは私の疑問として、なぜ著者らはoptic ataxiaとhemineglectとを別の論文にしたのか、なぜいっしょにしてdouble dissociationを示す論文にしなかったのはなのだろうか、というのがあったからなのです。そっちの方が論文として強いと思うし、PPCの機能局在についての重要なデータだと思うわけでして。SPLがoptic ataxiaの原因部位であるかどうかについては、上のコメントでも書きましたが、Rizzolattiの過剰な図式化に乗っただけのことでして、頭頂後頭接合部というのが本当だと思います。「neglectは注意障害だと捕らえるのはちょっと単純化されてすぎていて」 うーむ、そうなのですか。これも症状の多様性の問題ですね。一時期Jon Driverの論文がたくさん出たことがあって、あの頃から気にはなっていたのですが、hemineglectに関してはどうしてもきっちり勉強しなければならないようです。まずは御礼まで、というかおかげでまたこの日記の内容が充実するのでありました。
# pooneilご隠居、SFNには参加しますか?
# ご隠居好フォローありがとうございます。基本的におっしゃるとおりですね。。どうしてもメカニズムを考えるにあたって現象を抜き出して単純化した上で図式化することになるので、その過程として当然というか、やむを得ないことではありますが。でも、Rizzolattiの仕事(のスタイル)はとても好きです。って、おっと、たしか Milner論文へちょっと批判をしただけつもりだったのですが。ところで、てっきり humanの話だったのでSPLも humanのことをさしているのかと思ってしまったのですが、 saruの話だったのですね(当然か)。失礼いたしました。ついつい用語にひっぱられて、外延(内挿か?)してしまいました。 saruの lesion、electrophysiology、(やfMRI??)のきれいなデータと human neuropsychologyの実験の結果をつきあわせるのは大変なのかもしれないですが興味深いですよね。とくにparietalでは。ところで、 saruの neglectの論文っていろいろあるのでしょうか? Rizzolattiや Heilmanらの古い論文以降のフォローをしていないので。たしかしばらく前に Olsonらが SFNでなんか出していたような気もするのですが、その後どうなったのでしょう。
# ご隠居そうそう、今の本職は隠居老人なので。。。そのときまで生きていられたらSFNに遊びに行きたいですね。
# NHKご隠居さん、お元気ですか?NHKです。教えていただきたいのですが、neglectとextinctionは区別したほうがよろしいのでしょうか?
# ご隠居どうもです.neglectとextinctionは一見似ておりまた同時に起こることも多いですが(特に右中大脳動脈の大きな梗塞の後など),double dissociationが示されており機序も別々に考えうるので,区別したほうがよいのではないかと思われますが,いかがでしょう.
# pooneil>>今の本職は隠居老人なのでなにを言ってるんですか、ご隠居。私より若いんだから、またアグレッシブにいってくださいよ。もし体を悪くしてるんでさえなければ。Parietalの相同の問題およびneglectの動物モデルに関してはもう少し調べてみますが、少し本文で関連論文にリンクを貼っときました。Neglectよりはextinctionの方が動物モデルとしてはやりやすそうな気がするのだけれど、別ものなのか。これは重要なことを聞きました。
# NHKご隠居さん、ありがとうございました。他の方のレスをみるにつけても、言わずもがなですが、古典的なdorsal/ventral streamの見なおし、そして両stream間のinteractionのあたりに大きなbreak throughのタネがありそうな気がします。それにしても皆さんのコメントを拝見するとおぼろげながら、どなたなのかみえてくるところが面白いですね。
# NHK私は、申し遅れましたが、かつてご隠居さんと席をならべていた者です。
# ご隠居pooneilさん>> 私より若いんだから 確かに数ヶ月若いかもしれませんが、なにぶんこのところ苦労しすぎですっかり老けきってしまいまして。。。まだ論文の束の整理がついていないのですが、いろいろ教えていただいた論文を探して読んでみます。そう、Gaffanもあった、いろいろとぶっとんだこといっていましたね。たしかレビューもあった。楽しいヒトです。Perenin and Vighetto 1988、すっかり忘れてしまっていましたが、pubmedでabstractを見る限りではPPCに病変の主座がありそうな記載ですね。もうちょっとあたってみます。NHKさん、申し遅れても遅れなくてもわかりますよ。これからもますますお忙しくなり大変でしょうが、素晴らしいお仕事を楽しみにしております。
# pooneilNHKさん、確信度95%でしたが、ということは私とも席を並べてましたよね。なお、私自身はもう完全に匿名でなくなったのでそのへんに関してはご注意ください。>>両stream間のinteraction 昨日慌ててJon DriverのNature Neuroscienceのレビュー”Parietal neglect and visual awareness”を読み直してみましたが、Jon DriverはまさにIPLをdorsal streamのSPLとventral streamとをつなぐinterfaceと捉えているわけでして、そう考えるとますますhumanとnonhuman primateとのあいだでの相同が気になります。HumanのIPL/TPJに対応するのは7a/7bだろうか、それとももっとventralだろうか。ご隠居と意識について話をしていたときに、ventral stream偏重の私にparietalはどうなのかとご隠居がご指摘されたのを懐かしく思い出します。Parietalが意識に関与しているであろうことはJon Driverの言う通りであるけれども、それでもparietalが意識のcontentそのものにはなりえないだろうと思っています。このへん、もう少しまとめて書く必要が出てくることでしょう。
# CorreggioヒトのSMGが、7bに相当するという考えがあります。私もそう思っています。Zillesは、サルにはSMGはないといっています。7aは、角回にあたるんでしょうか。これは今だはっきりしません。TPJは、どこでしょう。TPJが、Size constancyに関わるという話しもありましたっけ。
2004年07月13日
■ Current Biology
- Magazine "Qualia." Christof Kochによる解説。というか自著"The Quest for Consciousness: A Neurobiological Approach"の宣伝ですな。この本専用のサイトへのリンク "The Quest for Consciousness"で1章と20章が読めます。
- Dispatch "Visual Cortex: The Continuing Puzzle of Area V2." Boynton and Hegdé。伊藤南さんのJNS '04 "Representation of Angles Embedded within Contour Stimuli in Area V2 of Macaque Monkeys."(V2のニューロンがangleをrepresentしている、というもの)に対するコメンタリ。
- / ツイートする
- / 投稿日: 2004年07月13日
- / カテゴリー: [Paper archive]
- / Edit(管理者用)
■ 臨床医のネクタイは病原菌のすみか?米学会で報告
以前書いて忘れてたのを貼ってみます。ってもうソースが消えてるし。
臨床実習先のニューヨーク市内の病院に勤務する医師ら医療従事者が着用したネクタイ42本で、ほぼ半数の20本から黄色ブドウ球菌などが検出された。患者と接することがほとんどない警備員のネクタイでは、検出されたのは10本のうち1本だけだった。
Yomiuri Online 6/2より。ていうか医師のネクタイと警備員のネクタイではなくて、医師のネクタイと医師のシャツとで比較してほしい。もしくはネクタイと手。それと医師/警備員とネクタイ/手との2x2でinteractionを出してくれ、と思うのです。そうしないと、単に医師がネクタイをしなければよいというだけの話なのか、そもそも医師の着るものが充分にきれいになっていなくて、院内感染を引き起こす元となっているのか、ぜんぜんわからない(ニュースの口調からすると前者として捉えられているようだが)、ってニュース記事につける文句ではないですな、これは。
# ご隠居
個人的な経験によると、ぼくみたいな貧乏人はネクタイをしていてもタイピンをもっていなくて、ベッドサイドで診察するときにネクタイがでれーって白衣から出てきてしまって、触れてしまうことがありました(過去形)。反省します。
# pooneilへー。へんてこな研究かと思ったけど、意外に的を射ているんだろうか。
■ Cerebrel Cortex
やっとオンラインのフルテクストが使えるようになったのでpdf取りまくり。
- "Oscillatory Synchrony in the Monkey Temporal Lobe Correlates with Performance in a Visual Short-term Memory Task."
- "Single Cell Integration of Animate Form, Motion and Location in the Superior Temporal Cortex of the Macaque Monkey." David I. Perrett
- "Color Discrimination Involves Ventral and Dorsal Stream Visual Areas." Guy A. Orban
- "Medial Prefrontal Activity Predicts Memory for Self."
- "Experience-dependent Changes in Basal Dendritic Branching of Layer 2/3 Pyramidal Neurons During a Critical Period for Developmental Plasticity in Rat Barrel Cortex." Karel Svoboda
- "Neural Correlates of Change Detection and Change Blindness in a Working Memory Task." Leslie G. Ungerleider
- "Enhanced Temporal Non-linearities in Human Object-related Occipito-temporal Cortex." Rafael Malach
- "Search for Color ‘Center(s)’ in Macaque Visual Cortex." Guy A. Orban
- "Task-independent and Task-specific Age Effects on Brain Activity during Working Memory, Visual Attention and Episodic Retrieval." Lars Nyberg
- "Perception-related Modulations of Local Field Potential Power and Coherence in Primary Visual Cortex of Awake Monkey during Binocular Rivalry." Reinhard Eckhorn
- / ツイートする
- / 投稿日: 2004年07月13日
- / カテゴリー: [Paper archive]
- / Edit(管理者用)
2004年07月12日
■ Trends in Cognitive Sciences 6月号
"Neuronal representations of cognitive state: reward or attention?" John H. R. Maunsell。世で研究されているattentionやreward expectancyのneural correlateはしばしば分離できていないことを指摘している。その理由としてabstではreward contingencyのパラメータのみがそれらの研究では振られていることを挙げています。このへんの分野の概念の複雑化と曖昧さとを考えるととても重要です。将来この辺の概念はもっと整理され、それぞれの概念間の関係がより確立していくことでまだしばらく先はやることはあるでしょうが*1、次は何か、それが問題なのです。
*1:実際には今まで40年の歴史を見て考えればしばしば複雑化の道を進むだけだったりするのだけれど。
- / ツイートする
- / 投稿日: 2004年07月12日
- / カテゴリー: [Papers_unclassified]
- / Edit(管理者用)
# mmmm
全般的にMaunsellの述べていることはもっともですが、reward or attention?という問いに意味があるのかどうかは疑問です。概念としてはattentionの方がreward expectationよりも上位だと思います。例えばMounsellはtask-difficultyとreward expectationとは逆相関するからそれらを分離することを提案してますが(それ自体は面白い実験であるし、Shidara & Richmond 2002やWalton et al. 2002はそれに関連したデータを示していると思いますが)、両者ともある種のattentionと見るのが妥当という気がします。passive attentionはともかく、active attentionの一般性を脳がどのように実装しているかというのは難しい問題で、reward expectationを含め、多くの詳細な研究の先にある問題だと思います。もちろんreward expectation意外のattentionという言い方はありで、そのうちのひとつがtask-difficultyとかeffortであったりするのではないでしょうか。
# pooneilありがとうございます。「Active attentionの一般性」、「Shidara & Richmond 2002やWalton et al. 2002」、このへんキーワードですね。もう少し読んどきます。それで、うーむ、Maunsellも挙げているように、Platt and Glimcherのreward probabilityにselectiveなLIPニューロンがじつはattentionによるものではないか、という問題はじっさい、このあいだのNewsome論文でも当てはまる話でして、ここはよくよく考えるべき問題ではないかと考えます。じつはこの間のjournal clubでもそういう指摘が出たのですが、Platt and Glimcherでとりあえず片はついていると思う、と言ってお茶を濁してしまいました。ふつうreward or attention?といった問題はより簡単な説明で済むほうで説明されるべきなのですが、attentionとrewardではどちらがより簡単な概念かとは言えないわけで、そこが難しい点なのだと思います。それが私が書いた、それらの概念が整理されていくことが必要である、と書いたことの動機でもあります。あと、mmmmさんが書いたことを展開すると、attentionというのは必ず何かほかのものに付随していて、reward expectationやtask difficultyやeffortというものがあるときにそれらに付随してくる、というふうに捉えられるかもしれません。この辺でattentionの心理学的研究についてもう少し知っておきたいところです。つまり、attentionがそれ以外のcognitiveなfactorとどのように関連付けられているか、といった点について。もしくはWilliam Jamesでも読んだ方がよいのかも。
- / ツイートする
- / 投稿日: 2004年07月12日
- / カテゴリー: [Paper archive]
- / Edit(管理者用)
■ JNS 6/16
- "Sequences of Abstract Nonbiological Stimuli Share Ventral Premotor Cortex with Action Observation and Imagery."
- "Brain Mechanisms for Inferring Deceit in the Actions of Others." Chris Frith and Passingham。こういうオモロげなのにはかならずChris Frithが入っているな。
- "The Role of the Central Nucleus of the Amygdala in Mediating Fear and Anxiety in the Primate."
- "Improvements in the Signal-to-Noise Ratio of Motor Cortex Cells Distinguish Early versus Late Phases of Motor Skill Learning."
- / ツイートする
- / 投稿日: 2004年07月12日
- / カテゴリー: [Paper archive]
- / Edit(管理者用)
2004年07月10日
■ Science Andersen論文つづき
"Brain Waves: Cognitive-based Neural Prosthetics"にて、Andersen論文が採り上げられてます。
- / ツイートする
- / 投稿日: 2004年07月10日
- / カテゴリー: [BMI, BCI and neural prosthesis]
- / Edit(管理者用)
■ Neuron 7/8
FulltextがScienceDirectのページになってる。なんかダサいな、これ。追記:わかった、そのデカい"neuron"っていうbannerがダサいんだ。
- "Posterior Parietal Cortex Neurons Encode Target Motion in World-Centered Coordinates." またPeter Thierか。"Posterior Parietal Cortex"と書いてMSTの研究なのはなんか馴染まないのだけれど。
- "Coincident but Distinct Messages of Midbrain Dopamine and Striatal Tonically Active Neurons." Hagai Bergman @ The Hebrew University。この間の岡崎のCOEシンポジウムに来てました。いい歳だと思うんだけど、引き締まったジーンズ姿でかっこよかった。
- / ツイートする
- / 投稿日: 2004年07月10日
- / カテゴリー: [Paper archive]
- / Edit(管理者用)
■ Nature 5/20
"Time waits for no man." KEVAN MARTINによるBenjamin Libetの新著"Mind Time: The Temporal Factor in Consciousness"の書評なのだけれど、"ネイチャ-月刊ダイジェスト 私たちは時間には追いつけない."を読むかぎりそうはわからなかった。著者は著者はって書いてるんだけれど、Libetはって書けばよいではないか。原文はそうなってるんだし。翻訳としてそれでは硬いというのだろうか。でも流し読みしたときは著者ってKEVAN MARTINのことかと誤解しましたけどね。そう思うのが普通ではないでしょうか。もっとも、一番悪いのは訳文にはLibetの著書のことがまったく書かれていないことですけどね。と思って紙に印刷されてる方見たらちゃんと書かれてました。それだけ。
- / ツイートする
- / 投稿日: 2004年07月10日
- / カテゴリー: [Paper archive]
- / Edit(管理者用)
■ The Journal of Comparative Neurology 5/3
いちおう貼っとこ。- / ツイートする
- / 投稿日: 2004年07月10日
- / カテゴリー: [Paper archive]
- / Edit(管理者用)
2004年07月09日
■ Science 7/9
"Cognitive Control Signals for Neural Prosthetics." R. A. Andersen @ caltechでNeural Prosthetics来たっすよ! 以前からannual reviewとかでもそっちを意識しているのはわかっていたけれど。
タイトルを読んだ印象:いままでのNeural Prostheticsってのは脳の信号を使って物を操作する、つまり運動機能の代替をしてやろうとするものだったわけです。たとえば、NicorelisやらSchawartzやら。んでもって、今回の論文ではもっと高次の指令("cognitive control")を脳から取り出してやろうというわけですな。選択のレパートリーが少なければ(Andersenですから、たとえば二つの選択肢のどっちへのmotor intentionを持っているか、とか)、それ自体は簡単なことのようにも思えるのですが。
アブスト読んでみました。うーむ、たんにM1からの信号ではなくてparietal reach regionからの記録でコンピューターカーソルを動かしただけのようにも思えるのだけれど(そんなもん、M1だろうがPMだろうがPPCだろうが可能な気がするし)。
また、この記録から、Expected value signals (= expected magnitude of rewardやらexpected probability of reward)に関わる信号が記録された、というのもLIPでのsaccadeタスクで見つかってくるのと同じ話だし(この前のNewsome論文、それからPlatt and Glimcher論文とか)。よって話としてはM1やPMからの記録でneural prosthesisするのと比べてメリットがなんであるかが焦点となることでしょう。アブストを見るかぎり、goalに関する情報とvalue signalに関する情報との両方がモニターできる、という点のようなんだけれど、それだけではM1からの記録のadvantageを越えられないように思います。
それは別として面白い問題であるのはたしかです。今回はカーソルを動かすだけのようですが、もしgoalの情報だけが得られて、それでロボットアームを動かしたらどうなるでしょうか。以前盛んに書いたように、goalのpositionという情報は関節の角速度のようなキネマティックな量に変換され、それが筋肉の張力のようなダイナミックな量に変換されます。つまり、goalの情報だけ与えられてもそれを実現するためのキネマティックまたはダイナミックなパラメータの組み合わせは無限にあるのであって、それの中のなにが選ばれるのか、というかいったいなにが起こるのか、ということに興味があります。
以上、まだ本文も図も読んでません。続きます。
追記:SI読んだところ、Plexon使ってるらしい。
- / ツイートする
- / 投稿日: 2004年07月09日
- / カテゴリー: [BMI, BCI and neural prosthesis]
- / Edit(管理者用)
2004年07月08日
■ Nature Neuroscience 7月号
"Deficits in saccade target selection after inactivation of superior colliculus." Edward L Keller @ The Smith-Kettlewell Eye Research Institute。
上丘はサッケードのinitiationだけではなくていろんな認知機能(Horwitz and Newsomeによる"target selection"やPeter Thierによる"covert shift of attention"など)に関わる情報を持っていることが主にsingle-unit recordingからわかっています。しかしそれらの研究に必ず付きまとう問題はそのような機能は上丘が担っているのではなくて、単に大脳皮質で表象されている情報が伝わってきているだけではないのか、といわれてしまう難点があることです。そのためには上丘のサッケードの機能を阻害せずに上丘での"target selection"などの機能だけを選択的に阻害できることを示すことが必要です。これは主にmuscimolを局所的に注入することで行われています。今回の論文もサッケード自体には影響がなくてtarget selectionだけが阻害されるような結果を提示しています。そういえばPeter Thierも玉川のシンポジウムで話をしたときには上丘のlesionやmuscimolによるreversible inactivationが重要である、というような言い方をしてました。あれの場合はサッケードのターゲットとattentionのcueの位置が離れていることから、両者を分離してinactivationすることが可能です。そのうち論文になって出てくるかもしんない。
- / ツイートする
- / 投稿日: 2004年07月08日
- / カテゴリー: [上丘、FEFと眼球運動]
- / Edit(管理者用)
- / ツイートする
- / 投稿日: 2004年07月08日
- / カテゴリー: [Paper archive]
- / Edit(管理者用)
■ PNAS 6/29
- "Beta oscillations in a large-scale sensorimotor cortical network: Directional influences revealed by Granger causality."
- "The loss function of sensorimotor learning." Daniel M. Wolpert @ University College London。
- / ツイートする
- / 投稿日: 2004年07月08日
- / カテゴリー: [Paper archive]
- / Edit(管理者用)
■ Science Express 7/1
"That's My Hand! Activity in Premotor Cortex Reflects Feeling of Ownership of a Limb." Richard E. Passingham。
うーむ、self-consciousnessに思いっきり関わる論文ですね。読まなくては。あと、Grazianoのfake handの論文はこれですね。
Science '00 "Coding the Location of the Arm by Sight."
- / ツイートする
- / 投稿日: 2004年07月08日
- / カテゴリー: [Paper archive]
- / Edit(管理者用)
# Correggio
7/2のScienceexpressには”That’s My Hand! Activity in premotor cortex reflects feeling of Ownership of a limb” Passinghamのグループからでてますね。Grazianoのfake handを使った実験のヒトversionですけど。
# pooneilCorreggioさん、ありがとうございます。リンク付けておきました。
2004年07月07日
■ Nature Neuroscience 7月号
- "Recalibration of audiovisual simultaneity." NTTの西田先生とCaltechの下條先生の名前が入っている。 私たちが目の前でボールがバウンドしているのを見るとき、バウンドする動きと地面に叩き付けられる音とが同じ物体(ボール)から来ていることを間違えることなく結び付けます。しかし、これはじつは不思議な話で、視覚情報が処理される脳の領域と聴覚情報が処理される脳の領域が違うだけではなくて、それらの処理されるのにかかる時間も違います。つまりそのような光と音との同時性(simultaneity)がどのようにして確立するか、という問題群があります。これはもちろん光と音だけの話ではなくて、視覚情報内でも動きと色との同時性がどう確立するか、という問題があり、そのような同時性が崩れるようなillusionがいろいろ考え出されています。Flash-lag illusionに関しても動きと光の点滅とのあいだでの同時性の問題であるといえます。ひとことで言ってしまえば、binding problem関連ですな。 そしてこの論文では、そのような視覚と聴覚との同時性の確立がその別々の時間に処理されたものを同時である、というふうに再構成している、という視点に立ちます。つまり、視覚の処理が行われた時間t1と聴覚の処理が行われた時間時間t2が経験上同時であったとき、この処理のずれt1-t2がゼロであるようにrecalibrationされている、という仮説を立てます。重要な点はこのrecalibrationがそれまでの経験によってcalibrateされているということです。よってわざとこの同時性をずらしたような刺激を使ってやってそれにadaptationさせるとこのrecalibrationが起こって同時性の基準がずれる、というのがこの論文でわかったことです。うーむ、面白い。そんなに重要なことが日常の経験から決められているとは。 この話は物体が近くにあるときの話なのだけど、もちろん光速と音速の違いによるcalibraionの問題というのもあることでしょう。
- "Automatic avoidance of obstacles is a dorsal stream function: evidence from optic ataxia." "The visual brain in action"をGoodaleと書いたDavid Milnerです。Parietal cortexにlesionのある患者さん二人へのテストの結果です。これについてはまた。
- "Deficits in saccade target selection after inactivation of superior colliculus." Edward L Keller @ The Smith-Kettlewell Eye Research Institute。これについてはまた。
- / ツイートする
- / 投稿日: 2004年07月07日
- / カテゴリー: [Papers_unclassified]
- / Edit(管理者用)
■ Amon Düül
Amon Düülは全て最高なわけだが。よく言われる1stおよび2nd と3rdとのあまりの違い(ドラムたたきまくり叫びまくりギターリフ繰り返しまくり、からアコギ主体の弾き語り系プラス催眠系ギターへ)について、日和ったと言ってもいいだろうし、政治的メンバーが抜けて音楽的になったというのも本当だと思う。けれど、あれって要するに"chill-out"した、と捉えればよいのではないでしょうか。ひとつのセッションぶっつづけ(20時間とも48時間とも言われる)から編集した1st, 2ndでの狂騒に荒れ果て、そこでレイドバックというかチルアウトしてしまったのが3rdであって、そのような「祭りのあと」感が3rdのに流れている空気だと思うのです。なんかドパミンだかアドレナリンだかが枯渇してしまった状態。それはサイケデリックロックからカントリーロックへと移ったByrdsなどのアメリカのバンドが辿った道ともほぼ同時進行だったのでしょう。そういうわけで、3rdのアウトテイク集が出たら"Chilling-out Düül"なんてタイトルはどうでしょ。
2004年07月06日
■ ω2
1-way ANOVAでのeffectの強さを示すindexとしてω2を計算してたんですが、値のrangeがなぜか0<ω2<1にならない。というわけで検証してみました。
ANOVAのテーブルを
| -- | SS | df | MS | F | P |
| effect | SSeffect | dfeffect | MSeffect | F | P |
| error | SSerror | dferror | MSerror | -- | -- |
| total | SStotal | dftotal | -- | -- | -- |
としたときに
ω2 = ( SSeffect - dfeffect * MSerror ) / ( SStotal + MSerror )
と計算されます。
よってこれの分母は正なので、分子を変形してみます。
SSeffect - dfeffect * MSerrorというわけで、F>1のときだけω2は正であるようです。Web探してもどこにもそんなこと書いてなかったけど。
= SSeffect - dfeffect * MSeffect / MSeffect * MSerror
= SSeffect - SSeffect / F
= SSeffect * ( 1 - 1 / F )
と書いてみてから、ω2==1になるのはSSeffect == SStotalになるときだし、SSeffect == 0のときには絶対ω2 < 0になる、と考えた方が早かったようです。やれやれ。
2004年07月05日
■ 6/29へのコメント再録
mmmmさん、その通りですね>>このサイトの意義。こういう論文を交えた話は事実関係のところであれこれやったほうが面白いわけで、いかに深く、核心までたどり着くか、という方向へ行きたいと思っております。また同時に、このことは各方面の研究の将来性と限界とを検討することになるわけで、今後の脳研究がどういう方向へ行ったらいいかを議論するための重要な材料にもなるであろうことを期待しております。ほんと、ここまで行けたらよいと思ってます。
■ 時系列解析
時系列解析での時系列データ内に相関があるのかあたりまえで、そのような相関をどう取り除いていくかというのが時系列解析には必須です。ARとかARMAとかのモデルのようにある時間のデータY(t)がY(t-1)やY(t-2)などからどのように影響を受けているかを取り込んだものもあります。
■ Science Newsome論文つづき
もうちょっと核心に向かいましょう。月曜の朝がjournal clubなもんで。
このScience論文にもいろいろな問題はあると思いますが(叩いて埃の出ない論文はない)、私が重要なものと思うのは、(1) この論文がglobalなmatchingとlocalなmatchingとを明示的に比較するようになっていない、(2) matchingよりももっと簡便な説明変数があるのではないか、という点です。なお、以下の文章はこの論文がacceptされるべきかどうかにcriticalな論点と今後証拠が出てくれば十分であるものとが混ざっています。ご注意を。
(1) この論文がglobalなmatchingとlocalなmatchingとを明示的に比較するようになっていない。これはかなり意図的に回避している感じがありますが、そういうわけにはいきません。まず、behaviorですが、Fig.2にあるようにいくつかのmeasureを持ってきてlocal matching lawがよく行動を説明することを示しているわけですが、これはロジックが間違ってます。Globalなmatching lawが当てはまることはよくわかっているのですから、local matching lawはglobal matching lawでは説明できないようなfractuationを説明できていることを示さなければならないのです。ちょっとまどろっこしいですね、つまり、Fig.2でlocal matching lawで説明できている、としているところからgobal matching lawでも説明できる部分を差っ引いてやるべきなのです。それでも説明力があるときはじめて、local matching lawがglobal matching lawに加えて役に立つ説明として採用されるべきなのです。GLM的に考えてやりましょう。Fig.1の選択を従属変数Y(t)、incomeを説明変数X(t)とします。X(t)はgobal matching lawによるregressor X1(t) (Fig.1Cでの黒い直線)とlocal matching lawによるregressor X2(t) (Fig.1Cでの黒い曲線から黒い直線を差し引いたもの)とに分解できます。それでY(t) = a1*X1(t) + a2*X2(t) + a3*X1(t)*X2(t) というふうにモデルを組んでやることができます。これでcoefficient a1は有意なのはわかっているので、coefficient a2が本当に有意なのは検証してやればよいのです。Fig.1Cなんてチャンピオンデータなんですから、全てがこうだと信用するわけにはいきません。交互作用の項があるのは、global incomeが1:1と1:8とではlocal incomeの違いの選択への影響が変わりうるからです。長くなりました。なんにしろ、彼らはこういうことはやっていません。つまり、読者は彼らの言うlocal incomeというやつにはglobal incomeの影響が足し合わさっていることを忘れてはなりません。じつのところ、global matchingだけが成り立ち、localなfractuationはそのmatchingからの予測のエラーを最小化するような強化学習のパラダイムで考えた方が尤もらしいと思うのです。だからこそ、global matchingかlocal matchingか、という問題はnontrivialなはずなのです。
また、電気生理学的にも、local incomeのことだけ考えているという点には難点があるのは上記の通りですが、要するに話はとしては、LIPのactivityでglobal matchingを説明することはできなかったということです。だからlocalな方へ話を持っていこう持っていこうという流れにFig.2がなっているのです。これはつまり、(a) 非験者はglobal matchingなどしていない、local matchingだけ、(b) 行動としてはglobal matchingもしているんだけれどLIPはそれには関与していない、のどちらかであるのでしょうけれど、この問題を彼らは回避したのです。それはもちろん、(a)のようなことは言うのは行動分析学者の反感を引き起こすでしょうし、(b)のようなことを言うのはせっかくmatchingのことをやろうとしているのに狙うべき領野を間違えた、ということになるのですから。というわけでどっちでも彼らにとって都合が悪いのです。そして我々はそれを責めるべきです。つまり、ここでもglobalなmatchingとlocalなmatchingとの問題を回避した影響が出ているのです。実のところ彼らはglobal matchingなんていらないという立場でやっているのですが(local matchingさえあればglobal matchingはいらない、もしくはglobal matchingはlocal matchingの結果をglobalに眺めたものに過ぎない)。
また、すでにmmmmさんが6/22のコメント欄で指摘していますが、電気生理でtauを決め打ちしている点には問題があります。個々のまたはpopulationのニューロンのtauと行動のtauとが必ずしも一致している必要はないのですから。じっさい、この問題は以前にMTでのperceptual decision研究のときにも繰り返された問題でして、いろんな感度を持っているニューロンがあって、それらをどう統合して行動に結び付けているのか、という話になるのです。
(2) matchingよりももっと簡便な説明変数があるのではないか。絶対に検討しなければならないことは、今回の「matchingに関連したvalueのneural correlate」という言い方以外のもっとより簡便(parisimonious)な説明はないか、ということです。たとえば、attentionだけで説明できないか、ただのmatchingとは独立なdecisionの確信度みたいなものでは説明できないでしょうか。Control taskとmatching taskとでの違いとは、両者を比べるmatchingをしているかいないか、というよりはFig.4では二つのターゲットからのselectionをしているが、Fig.3のcontrol taskではselectionをしていない、という違いではないでしょうか。とくに、control taskではlocal incomeの違いはサッケードするターゲットを選択するのに役に立ちませんが、Fig.4のmatching taskではlocal incomeの違いがサッケードするターゲットを選択するのにcriticalな情報となっています。つまり私はこの論文で見つかったことの本質は、LIPニューロンはただ単にlocal incomeの比率のような量をコードしているのではなくて、これがdecision makingに関わるときにのみdecision variableとしてその情報がLIPで処理されている、このことだけではないかと考えるのです。そう考えてみるとやっぱりPlatt and Glimcherとの違いがいったいどの程度あるのか、という問題になります。また、Jeff SchallがFEFでさかんに研究したような複数のターゲットからのselection、というパラダイムと結び付けて考える価値があるような気がしてきました。
(2)についてまた違った言い方にしてみましょう。Matching lawは二つの間での選択だけではなくて、ある一つの行動をするかしないかという選択にも応用できます。ということは、Fig.3にあるようなコントロールタスクでもそういうmatching lawが効いているということになります。また、このコントロールタスクはターゲットが一つしかないVI強化スケジュールをやっていることになります。つまり、Fig.4のマッチングタスクとFig.3のコントロールタスクとの間でのcriticalな違いとは、どっちのターゲットを選ぶかのdecisionが要るか要らないか、という点です。この点において、Fig.3とFig.4との違いからPlatt and Glimcher論文とNewsome論文との比較をすると、decisionをするときにreward関連の情報があるとそれを加味したようなrepresentationをLIPがするという意味では両者は同じです。Fig.3のような結果に関してもPlatt and Glimcher論文でも同様なコントロール実験を持ってくればよいだけで、わざわざmatching lawを満たすような実験にする必要性はここまでではありません。よって、もうひとつ「Fig.4のマッチングタスクとFig.3のコントロールタスクとの間でのcriticalな違い」を持ってくる必要がありますが、それは上記の選択をしているか否かを選択行動理論的にパラフレーズすれば、並列強化スケジュールであるか単一強化スケジュールであるか、二つの選択でのmatching lawと行動するかしないかのmatching lawとの違い、ということになるでしょう。
- / ツイートする
- / 投稿日: 2004年07月05日
- / カテゴリー: [価値による行動選択 (expected value)]
- / Edit(管理者用)
2004年07月04日
■ Science Newsome論文つづき
)") 、ついでに"Glimcher Review")。まあ、この間のVernon SmithとDaniel Kahnemanのノーベル経済学賞で心理学と経済学の組み合わせが脚光を浴びたから次は神経科学と経済学というわけですな。しかしこりゃあもう陣地は取られているような感じですな。なんかもっと違うことを考えなくては。
追記:mmmmさん、ありがとうございます。コメントではリンクできないようです。こちらでリンクしておきます。
そういう意味ではラットでたくさんありそうですね。
、ついでに"Glimcher Review")。まあ、この間のVernon SmithとDaniel Kahnemanのノーベル経済学賞で心理学と経済学の組み合わせが脚光を浴びたから次は神経科学と経済学というわけですな。しかしこりゃあもう陣地は取られているような感じですな。なんかもっと違うことを考えなくては。
追記:mmmmさん、ありがとうございます。コメントではリンクできないようです。こちらでリンクしておきます。
そういう意味ではラットでたくさんありそうですね。
- / ツイートする
- / 投稿日: 2004年07月04日
- / カテゴリー: [価値による行動選択 (expected value)]
- / Edit(管理者用)
# mmmm
このあたりの関連としては例えばCardinal et al. (2001): Impulsive choice induced in rats by lesions of the nucleus accumbens core (http://www.sciencemag.org/cgi/content/full/292/5526/2499)でしょうかね。
# mmrl彼らのlocal matchingは、過去の報酬履歴に対する効果が薄れて行くという話しであって、それに対して、上に書いてある図は未来の報酬に対する予測がdiscountされる方の話をしていてちょっと違うのではないでしょうか?さらに、「メイザーの」ではexponential discounting をしてましたか?わたしの理解だとhyperbolic discountingではないかと考えます。というのも、exponential curve を使うと睡眠が勉強を追い抜くことはないと思うのですがどうでしょう?上の図を作るとき実はhyperboric curveを使ったりしてませんか?(もしくは、discount rate を勉強と睡眠で違うものを用いていませんか?)
# pooneilありがとうございます。あんまりよくわかってないかもしれませんが、私の絵自身はexponentialで書いてます。勉強と睡眠とでdiscount rateは変えてありますが、それはその方がリアリスティックだと考えたからです。なんにしろ、今回の論文とはあんまり関係ないというのはその通りでして、なんか身につまされて面白かったので図にしてみた、というのが正直なところです。
# mmrlなるほど、違う種類の報酬には違うdiscount rateを用いるほうがリアリティがあるかな。どうも強化学習理論にとりつかれていると、最大化する報酬は同じ基準で比較するのがよろしいと頭からおもってしまっていけませんね。人間でもいろんなdiscount rateで価値を計算、比較してるってのは本当なのかもしれません。
# mmmmこの陣地争いに対等に加わっているのは、日本では銅谷さんのところが挙げられるでしょうか。Nature NeuroscienceのAOPに出た論文が、マスメディアで盛んに取り上げられていますね。この分野のサル屋さんはちょっと不甲斐ないというか、時代から取り残されているかも(落胆)。その次の時代を牽引すべくお互い頑張りましょうねー。
# pooneil”Prediction of immediate and future rewards differentially recruits cortico-basal ganglia loops”、これですね。玉川のCOEシンポジウムでもしゃべってましたね。プログラムに書いたメモに[GP SNr: stochastic action selection] -< [striatum: action value (expected reward / candidate action)] なんて書かれてます。なんにしろ、経済学でなくてもいいんだけれど、なんらかの形で役に立つような形を見出せるようにアンテナを張っておきたいと思います。これは基礎科学をする人間にとって忘れるべきでない点ですし。
# pooneil追記:銅谷さんの論文の解説がhttp://www.jst.go.jp/pr/info/info87/index.htmlにあります。
2004年07月02日
■ あーすばらしい。
Newsome論文スレッドはうまいこと伸びましたね。mmmmさん、mmrlさんのおかげです。大感謝。こういうふうにうまくいくスレッドがもっと増えて並行して複数流れるようになって、究極にはべつに私がいなくても参加者のあいだでやり取りが始まっている、というふうにメディア化してゆく、というのがいいなあと思います。1/17に書いた「2ちゃんねる形式で論文ごとにスレを立ててそれぞれにコメントを付けるという風にして、ゆくゆくは私以外もスレを立てられるようにする」と基本的には同じことですが、もう少しイメージが現実味を帯びてきました。あのとき書いた「匿名性」とはいま書いた「メディア化してゆく」ということだったんだと思う。あんまそっちばっかでも疲れるけど。追記:これってようするにスラッシュドットと同じだろうか。あれよくわかんないんですよ。
■ JNS 6/30
"Double Dissociation between the Effects of Peri-Postrhinal Cortex and Hippocampal Lesions on Tests of Object Recognition and Spatial Memory: Heterogeneity of Function within the Temporal Lobe." Timothy J. Bussey以前からrecognition memoryをラットで研究することあたりの話をしましたが、これも関係しそうです。Perirhinalのlesionとhippocampusのlesionとで機能のdouble dissociationがあることについてはMishkinの時代からAmaralやらGaffanやMurrayなどがさかんにやってきているわけですが、それをratの系に持ってきた、ということのようです。これはもちろんラットでやることのメリットがいろいろあるわけでして(ほんとうはマウスの方がもっとよいでしょうけど)、可能であるならより単純な系でanalogousなものを見れるようにしたい、という意味で評価できると思います。内容は読んでないけど、perirhinalがobject recognition、hippocampusがspatial memory、というのはあたりまえのようだが重要です。こんどはBuckley,Gaffan and MurrayのJNP '97 "Functional double dissociation between two inferior temporal cortical areas: perirhinal cortex versus middle temporal gyrus."を再現するようなITとperirhinalの比較に向かったら面白いじゃないでしょうか。Perirhinalはただのobject recognitionじゃないだろうし。
- / ツイートする
- / 投稿日: 2004年07月02日
- / カテゴリー: [内側側頭葉と記憶システム]
- / Edit(管理者用)
■ Nature 7/1
"Local sleep and learning."
NVもあるけど、ちょうどALLAN HOBSONがエッセイを書いている。
"A model for madness?" ALLAN HOBSON
- / ツイートする
- / 投稿日: 2004年07月02日
- / カテゴリー: [Paper archive]
- / Edit(管理者用)
2004年07月01日
■ Science Newsome論文つづき
"Matching Behavior and the Representation of Value in the Parietal Cortex." Leo P. Sugrue, Greg S. Corrado, William T. Newsome
mmrlさんから6/29のコメントにあるbayesian harvestingに関する詳しい解説と図が届きました。どうもありがとうございます。以下に掲載します。今日はmmrlさんによるゲストブログみたいなもんですので、<blockquote>に入れないで地の文に入れます。貼り付けてある図は図2の方です。図1は6/30のところに貼ってあります。はてなは一日一枚しか画像が貼れないもんで。なぜか図が小さくなってしまったので6/29にもっと大きい図2を貼っておきました。そちらをご参照ください。 (追記:はてなからの移行の際に図の配置を直しました。図1、図2ともに下に貼りました。)
mmrlです
先日説明したbayesian harvestingですが、図をはさんでいただけるということですので、ちょっと計算をしてみました。Corrado のポスターの式を参考にしました。すでにpooneilさんがやさしい言葉で説明されているVI-VIの場合に最適行動はmatching にほぼ等しくなることを式で書いただけというだけです。少々長くなりますが、私のような式や実際に動くプログラムやロボットで納得する人間のエゴとお許しください。
離散時間で進行するVI-VI平行スケジュールはある確率で報酬が出現し、取るまでそのままという形で実現できます。毎回red/greenに報酬が出現する確率をp_red/p_green とすると、greenに報酬が存在する確率P(r_green)は前回greenをためした回からの回数をkとしてP(r_green)_k = 1-(1-p_green)^k となります.
なぜなら,
1回では
p_green,
2回では最初の1回で出現する確率+1回目で出現せず2回目で出現する確率=
p_green + p_green(1-p_green),
3回目では
p_green + p_green(1-p_green) + p_green(1-p_green)^2
となり
k回目では
p_green (1 + (1-p_green) + (1-p_green)^2 + ... + (1-p_green)^(k-1))
となり等比級数公式から
P(r_green)_k = p_green(1-(1-p_green)^k)/(1-(1-p_green)) = 1-(1-p_green)^k
red を選び続ければ、p_greenがどんなに小さくてもP(r_green)_kは1に漸近しますから、
ほぼ100%でgreenに報酬があることになります。
ここで、baysian harvestingとは、報酬の存在確率を初回をP(r_red) = 0 , P(r_green) = 0 として上の式にしたがって計算し、その回(t)におけるP(r_red)(t) > P(r_green)(t)ならredを逆ならgreenを選ぶ最適選択者とします。とすると図1にあるようにredが上回ればredを選択し、確率0にリセット、green が上回れば..という周期パターンで行動を選択することになります。
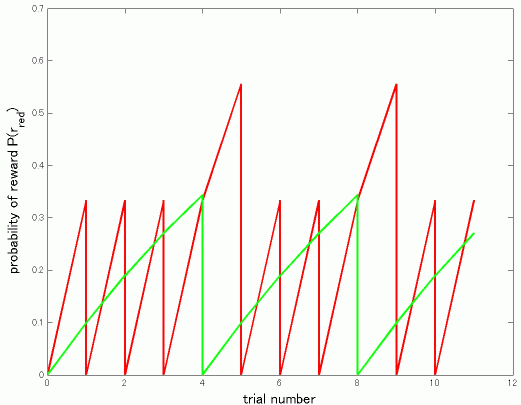
figure 1
ここで、p_red > p_green の場合に限定して考えると、redを選ぶ回数n_red(>=1)回に1回green を選ぶパターンになるはずです。(図1では3回に1回p_red = 1/3, p_green = 1/10)
したがって選択の割合はn_red:1となり、このときn_redは log(1-p_red)/log(1-p_green) の小数点きりすて整数になります。
なぜなら,n_red回はredを選ぶので報酬存在確率は
p_red > 1-(1-p_green)^n_red
次の回でこの関係が逆転するので、
p_red < 1-(1-p_green)^(n_red+1)
これを変形して (1-p_green)^(n_red+1) < (1-p_red)
の両辺をlog取って変形すると
n_red > log(1-p_red)/log(1-p_green) -1
となる最小整数がn_redであるから、
n_red = floor[log(1-p_red)/log(1-p_green) ]
ここでfloor[x]はx以下の最大整数
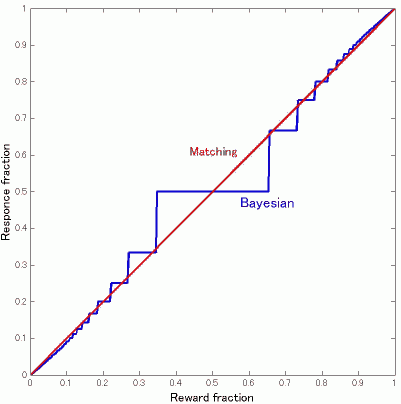
figure 2
これで得られるn_redを使って選択割合、n_red/(1+n_red)を報酬割合p_red/(p_green + p_red)の関数としてプロットすると図2のようになります。(p_red + p_green = 1/3の制約を与えています)。matching は、選択割合と報酬割合が一致するのですから傾き1の直線になります。ごらんのような階段関数となりますが、n_redが増えるにしたがって接近しているのがお分かりいただけるかと思います。ただこれが完全に一致するかというとどうかわからないところです。細かいところまでみると、どうも完全に一致しないように思います。
mmrlさん、どうもありがとうございました。すごい。
ひとつコメントですが、図2ってスムージングをかけてやると、逆S字型のカーブになりますが、それってgeneralized matching lawでいうところの過小な適応(強化の比率ほどには選択の比率が0.5からズレない)の形に近似できる感じがします(スムージングをしてから両座標をlogスケールにすると傾きが<1になる)。Bayesian harvestingによってやや過小ぎみな適応をしている、ということになるとじつはこっちの方がFig.1にある青線と黒線のずれをよく説明できるような気もしてきます。(あ、過小じゃなくて過大だったかも。)
そういえばVI-VI並列強化スケジュールとVR-VR並列強化スケジュールに関しては"高橋雅治(1997) 選択行動の研究における最近の展開:比較意思決定研究にむけて"に記載がありました。VI-VR並列ではVIのほうがより多く選択されることや遅延低減仮説に関して書かれています。やっぱりこのへんまだいろいろありそうです。
- / ツイートする
- / 投稿日: 2004年07月01日
- / カテゴリー: [価値による行動選択 (expected value)]
- / Edit(管理者用)
# mmrl
すみません、いきなり間違い発見しました。1つめの囲み記事の最終行は「ほぼ100%でgreenを選択するようになります。」です
# mmrlもとい「ほぼ100%でgreenに報酬があることになります。」
# pooneilありがとうございます。直しておきました。
# mmmmmmrlさん、大変分かりやすい図とご説明、ありがごうございました。非常によく解りました。さて、mmrlさんの図との対応を考えるにあたって、実際に何トライアルくらいに1回、報酬を貰っていたかが気になってきました。議論を単純化するため、overall maximum reward rate (0.15 rwd/sec)を採用しましょう。また常にtargetが表示され続けていたと仮定して考えると、1トライアルに要する時間は平均1950ms + rewarding timeとなるでしょうから、大体2秒と考えてよさそうに思います。0.15 rwd/sec = 1 rwd/6.66 secですから、平均3−4トライアルに1回の報酬があったと考えられます。これはgreenとredの両方を加算した条件ですから、p_red:p_green = 1:1の場合を考えると、red choiceに報酬が与えられたのはそのうちの半分ですから、6−8(平均約7(本当は6.6))トライアルに1回ということになり、red choiceによるreward rate、p(r_red)は単純計算で1/7 = 0.14となります。ここでmmrlさんの図2を見てみると、MatchingとBayesianとの間の一致はかなりよいレベルです。以下同様にp_red:p_green = 3:1の場合p(r_red) = 3/14 = 0.21p_red:p_green = 6:1の場合p(r_red) = 6/24.5 = 0.24p_red:p_green = 8:1の場合p(r_red) = 8/31.5 = 0.25となります(あくまで概算)。p(r_red) = 0.25でもMatchingとBayesianとの間の一致はかなりよいです。ということは必ずしもMatchingである必要はなくて、Bayesianでも説明可能なのではないかということになりそうに思えます。つまり、そろそろ別のターゲットに報酬がつきそうだということをベイズ的な計算に基づいて予想して、選択するターゲットを切り替えているのではないか、というように思えます。このことは、reward rateの小さい方のターゲットを選択する間隔がどれだけ周期的になっているか、また、その選択の切り替えが、mmrlさんの図1で予想されるタイミングにどれだけ一致しているかを検討することで見えてくるのではないでしょうか。ただ、pooneilさんの調査によると、CODを用いているとのことなので、この影響をどう考慮するかも考えなければなりませんね。
# pooneil直しておきました。
# mmmmん?↑の私のコメントでは、p_red + p_green = 1の制約が度外視されているような……。
# mmrlたしかに、bayesian でもよい予測はでき、行動が周期的かどうかを議論することで違いは出せると思いますが、bayesian は細かい短期記憶を必要とする(Matchingに比べて)複雑なモデルなので、オッカムのかみそりを使うと簡単なモデルで説明つくならそっちでいいじゃないかということになる。もし周期性が本当に観察されるなら、bayesianが本当っぽいということになるのかもしれませんが、それはでそうにないのではないかと考えます。なぜなら、bayesian では完全に行動ごとの報酬出現確率を既知として計算されている。本来は確率はサルにはわからないわけで、これを学習する必要があるはずです。しかも報酬出現確率は突然変化するために、常に学習しようとすることが報酬の最大化につながるはずです。とすると各試行におけるサルの行動は完全にはbayesianで予測できるとはおもえない。やるならば、長い時間スケールでは報酬出現確率を学習し、短い時間スケールでは報酬存在確率を考慮しながら行動をするのが最適になるのではないかと考えています。じつはこれはSugrue et al のsupporting online material のfigure S1 choice trigger averaging (CTA) of rewardと関係があって、彼らはこれはうまくexponential curveで近似できているといっていますが、私はこの曲線は異常に3-5trial 前までが大きくなってうまくフィットしない様に感じるのです。これは上に説明したような2重のタイムスケールを持つ学習/決定モデルでフィットできないかと考えています。モデルに関してはまた後日考えて見たいと思います。それと私のミスで、図2の制約条件はp_red + p_green = 1/3です。ごめんなさい。pooneilさん何度も申し訳ないです。overall maximum reward rate (0.15 rwd/sec)で1試行で約2secですから、報酬率は約0.3ということで、大体あっているはずと思って1/3にしました。たしかSugrueのポスターにも約1/3という記述があったように記憶しています。
# mmrlpooneilさんコメントありがとうございますスムージングを掛けてlogとってみるとどうなるか?って話ですが、実は図2の階段関数は、実はめんどくさくて数値解しか出していないので、解析解をみてみないと厳密にはなんともいえないです。ただこのカーブはp_red + p_greenの値に依存して変化します。1/3のときはよくあっているように見えますが、1にするとS字カーブになることを確認しています。
# mmmmああ、そうか。単純に1/3を1:1, 3:1, 6:1, 8:1に分けるだけでよくて、実際の実験状況でのp(r_red)はminimumが1/3 * 1/2 = 0.167、maximumが1/3 * 8/9 = 0.296となると見るだけで良かったのか。先のはダサダサしたね(笑)。
# mmmm報酬出現確率が本当にサルにとって未知であるかどうかは検討する必要があるのではないでしょうか。長期のトレーニングによってサルは、トータルの報酬出現頻度が約1/3であり、しかも報酬のred/green (or green/red)の比率が、1/1, 3/1, 6/1, 8/1しかないことをサルはすでに学習済みである可能性があると思います。するとブロック毎に学習すべきは、どの比率であるかの選択だけでよいことになります。そして各ブロックは100-200 trialsを含んでいますから、そのほとんどを既知の確率に基づいて行動していることはありそうなことではないでしょうか。
もしかしたら、mmrlさんの考えと基本的には同一なのかもしれません。モデルの方も期待しています。
なるほどそうか、長い訓練でいくつかの確率のパターンをいくつかあらかじめ知っていてそれを選択するということも考えられますね。考えていませんでした。わたしが考えていたのは単純にbayesianで予測される報酬の確率と実際の報酬との誤差が確率をupdateさせるモデルです。mmmmさんの言われる方法でも別にモデルができますね。ただ、いくつかのモデルを当てはめることができても、これをどうやって比較したらいいんだろう...。自分でVI-VIの行動データを取ろうかな? もう少しつめる必要がありそうです。
# mmrlbayesian harvesting にchange over delay を入れてみた場合も、一応計算してみました。結果はあんまりかわらなくてがっくししたのですが、まず、CODのもっとも簡単な形はswitchした後は1回は報酬をあげない、だけどもう一回選んだらあげることにします。Sugrue 04のReference and Notesの11に書いてあることをそのまま理解するとこれをしていることになります。ならば簡単switchしたらもう一回は必ず同じ色を選ぶことにする。問題は、これをするとswitchしたほうがよいかstayしたほうが良いかは2試行先まで見て平均報酬の良いほうを選ぶことです。以前と同様にp_red > p_green の場合に限定して話を進めます。redにいるとき、2試行まで見越してやるとp_red < {1-(1-p_green)^n_red+2}/2の関係が成り立つ最小整数n_redを求めればn_redを連続で選択する回数がわかります。結果はn_red = Max{floor[ (1-2p_red)/(1-p_green) ] -1 , 2}となります。同様にn_greenももとめて、p_red/(p_red + p_green)に対してn_red/(n_red+n_green)を求めれば同じようなグラフが得られます。結果はほとんど同じ、ステップは細かくなりましたが、概要は同じでした.これを使ってモデルは作ってやってみましたが、どうもstay dulation(Fig 2 F)がサルの結果にあわないようです。CODの関係でどうしても2回が多くなる。やはりここまで複雑なことは考えていないかも知れません。
# pooneilp_red + p_green = 1/3、直しておきました。
# mmmmmmrlさん、お疲れさまでした&ありがとうございました。Fig. 2Fでは2回が多いですから、モデルの方がそうならなかったという意味ですね?行動の解析には報酬の各比率のトライアルがが必ずしも均等に含まれているとは限らないので、比率毎に分けて、彼らの行動の結果を見てみたいところです。そのデータがあれば、もう少し妥当なモデル化ができるようになるのではないかと思いますが、これは彼らのみぞ知るデータで、続報(がもしあれば)に出てくるのかもしれないので期待したいところです。
お勧めエントリ
- 細胞外電極はなにを見ているか(1) 20080727 (2) リニューアル版 20081107
- 総説 長期記憶の脳内メカニズム 20100909
- 駒場講義2013 「意識の科学的研究 - 盲視を起点に」20130626
- 駒場講義2012レジメ 意識と注意の脳内メカニズム(1) 注意 20121010 (2) 意識 20121011
- 視覚、注意、言語で3*2の背側、腹側経路説 20140119
- 脳科学辞典の項目書いた 「盲視」 20130407
- 脳科学辞典の項目書いた 「気づき」 20130228
- 脳科学辞典の項目書いた 「サリエンシー」 20121224
- 脳科学辞典の項目書いた 「マイクロサッケード」 20121227
- 盲視でおこる「なにかあるかんじ」 20110126
- DKL色空間についてまとめ 20090113
- 科学基礎論学会 秋の研究例会 ワークショップ「意識の神経科学と神経現象学」レジメ 20131102
- ギャラガー&ザハヴィ『現象学的な心』合評会レジメ 20130628
- Marrのrepresentationとprocessをベイトソン流に解釈する (1) 20100317 (2) 20100317
- 半側空間無視と同名半盲とは区別できるか?(1) 20080220 (2) 半側空間無視の原因部位は? 20080221
- MarrのVisionの最初と最後だけを読む 20071213