[月別過去ログ] 2012年03月
« 2012年02月 | 最新のページに戻る | 2012年04月 »2012年03月29日
■ 超解像とマッピング
複数フレームからの超解像画像みたいな話題の場合はわざと解像度を下げた画像を作ってそこから元画像を再現するみたいな例がよくある。そういうばあい、samplingがせいぜい2倍とか4倍とかそういうのしか見ない。もっとセンサーの数が少ない例はないのだろうか。
たとえば、4*4のセンサでちょっとずつずらしながら128*128の画像を再構成するような。っていうかそれだったら一個のセンサーをスキャンして128*128の画像を作ればよいのか。 要は電極でニューロンの記録するという状況でなんか超解像的なことが応用できないのかって話なんだけど。
テトロードってのは三角測量みたいなもんで、一個のセンサだとシグナルの大きさとシグナルからの距離が分離できないから、複数のセンサを使ってそれを分離するということに原理的にはなるのだけれども、テトロードの場合、センサの配列の仕方に制約があるから完全にそれが出来ているわけではない。そういうわけで、これはべつもん。
佐藤先生のforward modelを作ってVBでセンサの数よりももっと詳しく推定をするってのと同じ問題。でも電極は動かせないので、超解像みたいなことをそのまま応用することは出来ない。
一個のセンサーをスキャンして128*128の画像を作ればよい<< これかならずしも正しくなかった。そんな狭い部分を記録できるセンサがあればよいが、もし、受光する部分が広いとしたら、それを使ってどのくらい細かいものを再構成できるか。
具体的な問いとしては、128*128の元画像から4*4のモザイク画像を様々なshift(とりあえず平行移動だけ)で作ってやって、そいつらからどのくらい128*128の元画像が再現できるかをやってみればよいのだな。
layer 2-3はoptical imagingのほうがほとんどすべての意味で勝っているからそちらのことを考えるよりかは、layer 5のprojection neuronの活動の空間分布みたいなことを電極アレイで解決できないか考える。てかもう超解像関係ないし。
一般的なシリコンプローブみたいな直線的にコンタクトが配列しているものでもなく、剣山電極みたいな間隔が開きすぎているものではなくて、ある立方体の中で3D方向すべてで50um間隔でコンタクトが配置されているようなものから記録することが出来れば、本気でポアソン方程式とか使えるのだな。
なんかlayer 5あたりに電位感受性細胞をinjectionしておくと、そいつらが他のニューロンとはシナプスを作らずに脳表までprocessを延ばしてくれて、その部分をoptical imagingする。うーむ、SFすぎるか。
SFついでに考えると、そもそも細胞をinjectionする必要ないな。いまあるニューロンを電位感受性細胞として使えばよい。なんか感染させてやって、そのニューロンの出力とシナプス入力を停めてやって、そいつらの軸索側枝が光学測定できるくらいの浅さまで伸びてくれればよい。
なんか自分で言っててわけわからなくなってきたぞ。つまり、ニューロンはそれじたい前シナプスニューロン活動のセンサなのであって、べつに出力停める必要もない。シナプスの入力は全くのランダムというわけではないので空間的な統計があまり使えない。
だから電位感受性センサとしてだけ使うとしたら、dendrite全体にチャネルが分類してしまっていると再構成しなくてはいけなくなるので、細胞体のチャネルだけで、しかも超局所的ではない細胞外電位が記録できるようなことが可能であればよい? そうするとチャネルのコンダクタンス的には既存のものでは向かない?
あとで見たら何言っているか分けわからない。だが残しておく。
2012年03月22日
■ ヒトの脳にはニューロンがどのくらいあるか
ヒトの脳にニューロンがどのくらいあるかちゃんと測った論文がAzevedo et.al. 2009で、860億個だそうな。体積の80%を占める大脳皮質に19%のニューロンがあって、体積の10%を占める小脳に80%のニューロンがある。なにげに小脳すごい。この事実は小脳の重要さを強調するために引用されたりする。たとえばRoger LemonのLife without a cerebellum。
小脳の説明をするのに、大脳はCPUで、小脳はGPUで並列計算するのに向くアーキテクチュアになっていて、わざわざM1から長い線維伸ばして計算させてまた戻すとかコスト払っているので、大脳皮質の構造では出来ないようなことをやってんだろう、とか説明をしたりする。ほんとうのところはわからない。小脳に聞いてください。
Azevedo et.al. 2009でのもう一つのポイントは、ニューロン-グリアの比率は1:101:1くらいで(3/26訂正。Thanks to Gayaさん)、これはほかの霊長類でも同じということ。そういう意味ではヒトの脳はマカクとかと比べてとくに特別なわけではない。
The human brain in numbers: a linearly scaled-up primate brain 同じ著者が書いたレビュー。これの図3がよくて、いろんな齧歯類、霊長類の脳の重さとニューロン数が一覧できる。
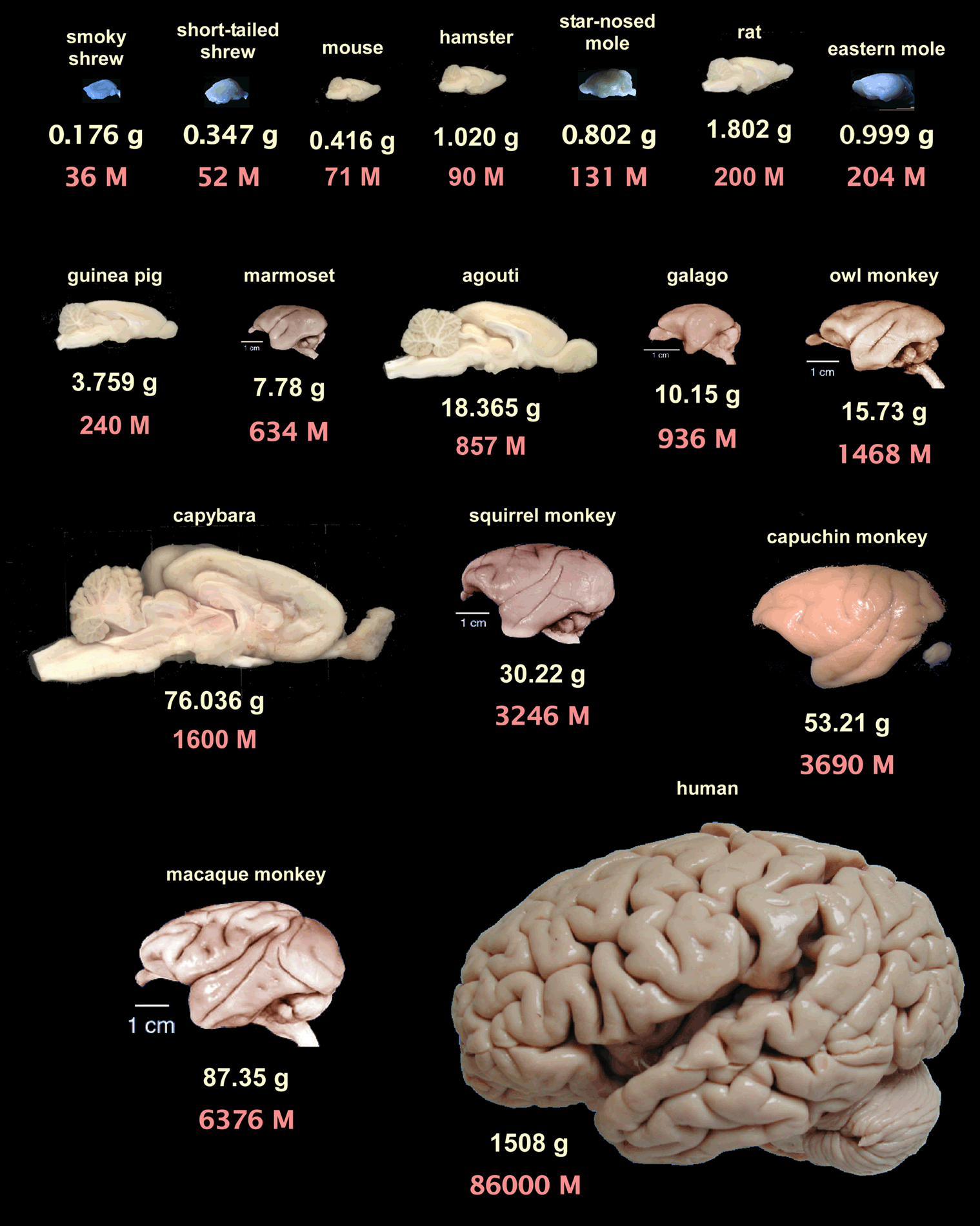{kind=link}
それによると、マウスは脳重が0.4gでニューロン数が7千万、ラットは1.8gで2億、マーモは7.8gで6.3億、マカクは87gで63億、ヒトは1500gで860億。こういうデータを持っておくと講義とかそういうときに捗るんでオススメ。
追記3/26: ニューロン:グリア比が1:1ってのはけっこう重要な話で、上述のFrontier誌を読んだところ、これまで信じられていた1:10というのはほとんど根拠がないうえに、神経神話「人間は脳の10%しか使えていない」の元となっている可能性があると。グリア比が多いことが分かっているのは視床だったり、ventral pallidumだったりというところでの報告だけで、大脳皮質ではグリア比が2を超えることはない。いちばんニューロン数の多い小脳では逆にニューロンの方が多くて、ニューロン:グリアの比は>=25:1 (at leastって書いてある)。 以上の数字はHerculano-Houzel本人の仕事ではなくて、それまでの文献から得られたものなので、カウント方法にはそんなに依存しなさそう。
# Gaya
あれ?この論文では「ニューロンとグリアの比率は、かつては1:10だと言われたこともあったが、実際には1:1だった」と主張していますよ。それがメインの発見だからこそ、論文のタイトルにもなっているのかと。
# pooneilあ、ほんとだ。斜め読みで完全に誤読してました。もうちょっと読んでから直しておきます。ご指摘ありがとうございました。
2012年03月09日
■ 「お話としての説明」と「科学としての説明」という対比
Per BakのPNAS1995 Complexity, contingency, and criticalityの初めのほうを読んでた。History vs Scienceという対比があってなるほどと思った。
History(=進化とかそういった一回性の現象)にはnarrative accountつまり物語的なイベントの連鎖としての説明が行われる。ここでは砂山モデルで戯画的に説明されているが、砂山のシミュレーションで大きな雪崩が起きるとき、それはnarrative accountでは、「いくつかの悪い偶然が重なって予想外に大きい雪崩が起きた」といった説明がされる。
しかし物理学的説明からすれば「それはSOCであって、そのような大きな雪崩はpower lawからすればなにか特別なところがあるわけではない」ということになる。(因果的な説明ではなくて、統計的な描写でしかないところに注意)
ここでもhindsight (後付け)という表現が使われているが、けっきょくブラックスワンの特徴として挙げられる「あとから後付けでいくらでも説明できてしまう」という部分はたとえ科学的な言葉を使った説明だったとしてもnarrative accountでしかないのだということが分かる。
おそらく認知神経科学のほとんどの部分はそういったnarrative accountでしかないのだろう。たとえば脳のある部分が損傷したから、XXの処理が出来なくて、その結果YYの課題の成績が低下する。しかし様々な要因(脳内及び行動面での代償とか)があって予想通りとはならない。
だから、論理の連鎖をあんま長く続けることは避けた方がよい。もしくは関係ありそうなパラメータを全部突っ込んでシステムバイオロジー的に行うか。
前者の方策は問題をなるたけ細分化するかまたは禁欲的に操作的に行うという方策。たぶん行動分析というのもなるたけ説明変数を減らそうという方策で前者の範疇に入るのだろう。前者はどこか「禁欲的」な側面を持つ。
意識とかエピソード記憶といった一回性の事象を科学的に説明するというときに、事象の方だけじゃなくて、説明の方にもnarrative accountと物理的説明という、違ったものがあることを考えることは重要だ。narrative accountに頼らないのなら、脳と身体と環境との物理的因果の動力学を描写することこそが一回性の事象の説明となる。
あともうひとつ面白いと思ったのは、恐竜の絶滅も様々なスケールの絶滅がある中で考えれば確率的に起こりうるものだという話で、なるほど、なんだかものすごく特別なイベントのように思えたけど、これもpower lawでいいじゃん(本文中ではこういう書き方はしてないが)とけっこう得心した。
まあこうやって、揺れながら、人混みをうまくすり抜けるつもりで左右に揺れ動きながら、かえって人にぶつかってしまって遅くなるような(じっさいよくあるんだこれが)、無駄な動きをたくさんするのがどうやら私の行動原理らしくて、たぶんなんかメタなパラメータを最適化しているつもりなんだろう。
sandpile modelをmatlabで書いてみたくなったが、禁欲してコードを探すところまでにしておく。
- sandpile (matlabcentral)
- Pattern Formation in Growing Sandpiles (pdf)
- Gradient Based Bak-Tang-Wiesenfeld Sandpile Model (pdf)
2012年03月03日
■ そういうみっともなさを愛する。
空を飛びたいなんて思ったことないな。海水浴をしていて、ずいぶん離れたところまできて、深いところの冷たい水を感じたりすると、ぞっとする。深夜に丸の内のオフィス街あたり自転車で通って、誰もいないってことに気が付いてぞっとしたり、深夜のLAの街を自転車で徘徊しながら、自分を知っている人間はどこにもいないんだということに気づいてぞっとしたり。
いやいや、それはある、ふつうにある。大雨が降って、買った水が全部無駄になって、花火工場が爆発して、いつまでもあれが消えないままに、いやそれはどうでもよい、頭がはっきりしているあいだにたいせつなことに取りかかることにしよう。バカバカしいくらいに積み上げられた蛍光灯の束。
農道を走る自転車と遠くのたき火の臭い。顕微鏡の下でそっとネジを回しながら、船底で海水が流入してくるのをそっと待つ。腕がしびれて、前を向いた瞬間に、どっちが上だかわからなくなった。外巻きの渦と内巻きの渦がぶつかり合い、無限大を表象する。
いやいや、それはそうではない、と持ち替えた手がそもそも左右が間違っている。牛乳パックが日本刀で手品のように切られ、中の液体がスローモーションで落ちてゆく。眼球の毛細血管がゆっくりと膨張してゆく。バスケットボールが後ろ回転をしながら放物線を描いて、ガラス窓を直撃する。
蜘蛛の巣には誰もいなくて、近くで落ちた実がひからびている。その周りは住宅街で、蛇やら狸やらが追い出されては車に轢かれる。手のひらの皺から三十通りの単語を拾い出す。緊急用の赤いランプが点灯しっぱなしで、もはや意味を失っている。
オーバーロード、オーバードーズ、ウーバーメンシュ。鉄球と魂とペンドラムの歌。練乳をかけた植木鉢に集められた30個の錆びた缶バッチ。独学で至った境地。血に染まった川。顔に修正の加えられた白黒写真とステッキと山高帽。皮ははち切れそうになっているし、枝からは千切れそうになっている。
一斉に押されるスイッチ達。夕日の中を飛ぶ航空機。道ばたに置かれたままになっている白いペンキ。すべては止まっていて、ここから遠く離れていて、私が別れを告げるためだけに存在している。
「黒の二枚刃」「すぐのお出かけ」「ハッシュタグ、ハッシュタグ」
寒い道を康生町から東岡崎の駅まで歩いて行った。東岡崎の駅前は忘年会終了後の客でごった返していて、岡崎の夜とは思えないくらいの様子だった。ちょっと離れると静かさが際だって、なんだかスキー合宿で雪道を買い出しに出かけたような気分がしてきた。
iPhoneの光を頼りにして、足下のあやしい暗い道をたどって向かう。街を縦横に駆け巡るコミュニケーションの束。落ち葉とともに掃き集められた昆虫の死骸。鋭角に割れた窓ガラスが下腹部に突き刺さっているイメージ。路上に敷かれた鉄板から立ち上る冷気。
プルーフロックの"I do not think that they will sing to me" …なんて、なんという自己憐憫の固まりだろうか。ぜんぜんかっこよくないぞ。ダサダサだ。「キリッ」とか付けちゃうぞ。「私のために歌ってくれるとは思わないけど(キリッ)」そういうみっともなさを愛する。
お勧めエントリ
- 細胞外電極はなにを見ているか(1) 20080727 (2) リニューアル版 20081107
- 総説 長期記憶の脳内メカニズム 20100909
- 駒場講義2013 「意識の科学的研究 - 盲視を起点に」20130626
- 駒場講義2012レジメ 意識と注意の脳内メカニズム(1) 注意 20121010 (2) 意識 20121011
- 視覚、注意、言語で3*2の背側、腹側経路説 20140119
- 脳科学辞典の項目書いた 「盲視」 20130407
- 脳科学辞典の項目書いた 「気づき」 20130228
- 脳科学辞典の項目書いた 「サリエンシー」 20121224
- 脳科学辞典の項目書いた 「マイクロサッケード」 20121227
- 盲視でおこる「なにかあるかんじ」 20110126
- DKL色空間についてまとめ 20090113
- 科学基礎論学会 秋の研究例会 ワークショップ「意識の神経科学と神経現象学」レジメ 20131102
- ギャラガー&ザハヴィ『現象学的な心』合評会レジメ 20130628
- Marrのrepresentationとprocessをベイトソン流に解釈する (1) 20100317 (2) 20100317
- 半側空間無視と同名半盲とは区別できるか?(1) 20080220 (2) 半側空間無視の原因部位は? 20080221
- MarrのVisionの最初と最後だけを読む 20071213