[月別過去ログ] 2016年06月
« 2016年05月 | 最新のページに戻る | 2016年07月 »2016年06月15日
■ 駒場学部講義2016 「意識の神経科学:「盲視」と「統合失調症」を手がかりに」レジメアップしました
駒場学部講義2016 「意識の神経科学:「盲視」と「統合失調症」を手がかりに」レジメアップしました。
今年は神経現象学の話を入れてみた。うまくいくとよいのだけれど。乞うご期待!
- / ツイートする
- / 投稿日: 2016年06月15日
- / カテゴリー: [脳科学メモ] [駒場学部講義2016]
- / Edit(管理者用)
2016年06月11日
■ 駒場講義2016/6/15の準備メモ
毎年恒例のオムニバス講義が近づいてきたのでそろそろ準備を始めてる。今年は6月15日(水)で、タイトルは「意識の神経科学:「盲視」と「統合失調症」を手がかりに」。2012年から毎年、5年目となった池上高志さん @alltbl の人間情報学VIでの学部講義 105min * 2。
昨年の講義の準備の様子やレジメなどはこちら:[カテゴリー別保管庫] 駒場学部講義2015
今年も例年通り、前日くらいにレジメをブログおよび生理研のサイトにアップロードする予定。
さてそれで内容をどうするかなのだけれども、昨年導入した統合失調症と異常サリエンス仮説の話を今回はさらに広げてゆくつもりで考えてた。神経現象学に関する話題(ブログ記事参照)は前回は見送ったのだけど、金井さん(Araya/東工大) @kanair_jp のConsciousness Clubで先月喋ってみたら、ひととおり話せることが分かったので、そちらを膨らませるという方向もある。
Consciousness Clubのときには、早々にヴァレラの「神経現象学」に飛んでしまったけど、順序立てて説明するためにはデネットの「ヘテロ現象学」を説明して、いま我々が認知神経科学者として行っていることはまさに「ヘテロ現象学」であるということからスタートする必要がある。
このあたりについては、ギャラガー&ザハヴィ『現象学的な心』合評会のときに作った話があるので、そこで端折った部分を入れて丁寧に話をすることができる。
でもって、NCCから力学系へみたいな話をするときには、そもそもDavid MarrのVisionにおいても表象(representation)と過程(process, computation)とが並べて議論されていたことに言及すると良いのではないかと考えて、原本にあたってた。
つか以前このあたりについてはブログに書いたことがある。「MarrのVisionの最初と最後だけを読む」とか「Marrのrepresentationとprocessをベイトソン流に解釈する(2)」とかを自分で読み返す。
そうやって調べものしていたら、"David Marr vs. James Gibson"という記事を見つけた。マーの議論はhomunculus fallacyだって言ってる。
よくよく見たら、PythonでのスパイキングニューロンのシミュレーターBrianの作者ではないの。上丘のスパイキング・ニューロン・ネットワークの仕事をここ数年進めているので(たとえばBMC Neuroscience)、ちょくちょくいじってた。
こういうもの作っている人がKevin O’ReganとかAlva NoëとかMerleau-Pontyに言及しているというのはすごく興味が惹かれる。必然のような、逆説的のような。
- / ツイートする
- / 投稿日: 2016年06月11日
- / カテゴリー: [視覚的意識 (visual awareness)] [駒場学部講義2016]
- / Edit(管理者用)
2016年06月07日
■ 静止膜電位はどうやってできるの?
神谷さんのツイートで言及されていた神経細胞の膜電位の話についてだけど、
高校生物の静止膜電位について質問です。静止膜電位は、外側が+で内側が-になる…と習ったのですが、それはなぜでしょうか? プラスイオンが外側に多く、内側に少ないからか?と思ったのですが、専門書を見ると、むしろ内側のほうにプラスイオンが多いです。(Yahoo知恵袋より)
これはかなり根本的な間違いをしている。神経細胞膜の内側も外側も(マクロには)+イオンと-イオンは釣り合っていて、電気的に中性になっている。+イオンと-イオンとが分極しているのは神経細胞膜の近傍だけであって、分極したイオンが膜を挟んで引き寄せ合って分布することで神経細胞膜はコンデンサーとして働く(図1)。
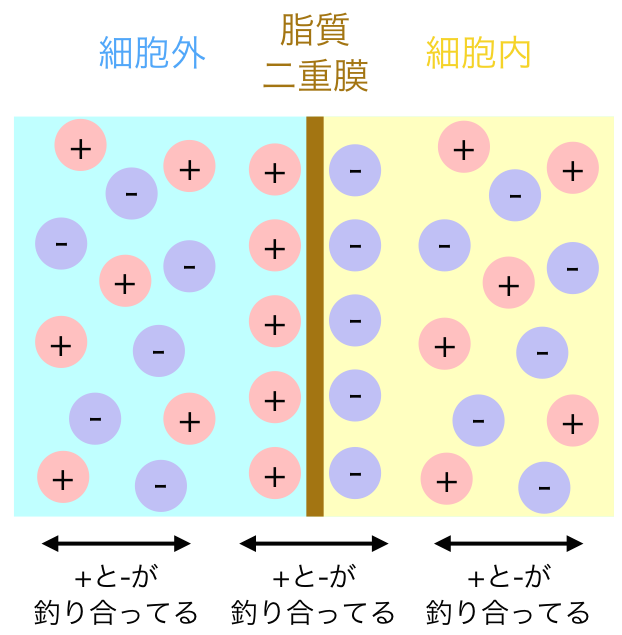
図1 神経細胞膜の内外でのイオンの分布
(カンデル本5ed. Figure6-1を元に作成)
巨視的に見て+イオンと-イオンが釣り合っていないなんて事態はよっぽどのことがないとおこらないということは化学を知っていれば理解しているはずだけれども、でもじゃあなんで電位差ができるのかということはネルンストの式をちゃんと理解しないと説明できない。
「これでわかるニューロンの電気現象」は神経生理学でつまづきやすい部分を対話形式で説明してある名著なのだけれども、この本の第1章はまるまるこの静止膜電位の問題に充てられている。
「キャンベル生物学」を見てみたけれど、たしかに細胞膜の内外のイオン濃度が代表的なものしか書かれていない。これがYahoo知恵袋での混乱のもとだった。じつはカンデル本5ed.(Table6-1)にも代表的なイオン濃度しか書かれていない(図2)。もちろん、Figure 6-1とその説明文章をちゃんと読めば電気的に釣り合っていることは書かれているのだが。
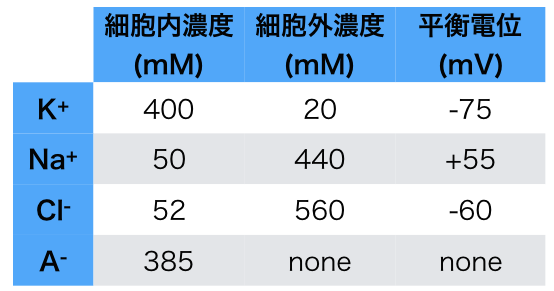
図2 イカ巨大軸索での主要なイオンの濃度
(カンデル本5ed. Table6-1を元に作成)
「これでわかるニューロンの電気現象」のコラム5.1 (p.158)では、陽イオンの濃度の総和と陰イオンの濃度の総和が細胞膜の内外ともに釣り合っている、つまり電気的に中性だということが図で示されている。(ちなみにこの図の引用元はAidley "The Physiology of Excitable Cells"とのことだが、google booksからは該当ページ見つからず。)
じゃあどうやって電位差ができているのかという説明なんだけど、教科書をいくつか読み比べて、どんな説明をしているか調べてみた。
「これでわかるニューロンの電気現象」では「濃度の違うKCl溶液がKイオンだけ透過する半透膜で遮られたとき」という例を使っている。ネルンストの式が濃度勾配による仕事と半透膜間の電位差による仕事が釣り合った平衡状態であることをわかりやすく説明している。
この本の第一章のネタ本はreferenceとして挙げられているとおり、Eckert and RandallのAnimal Physiologyの5章なのだけど、この図を見れば分極は膜の両端でだけ起きているということがよく分かる(図3)。
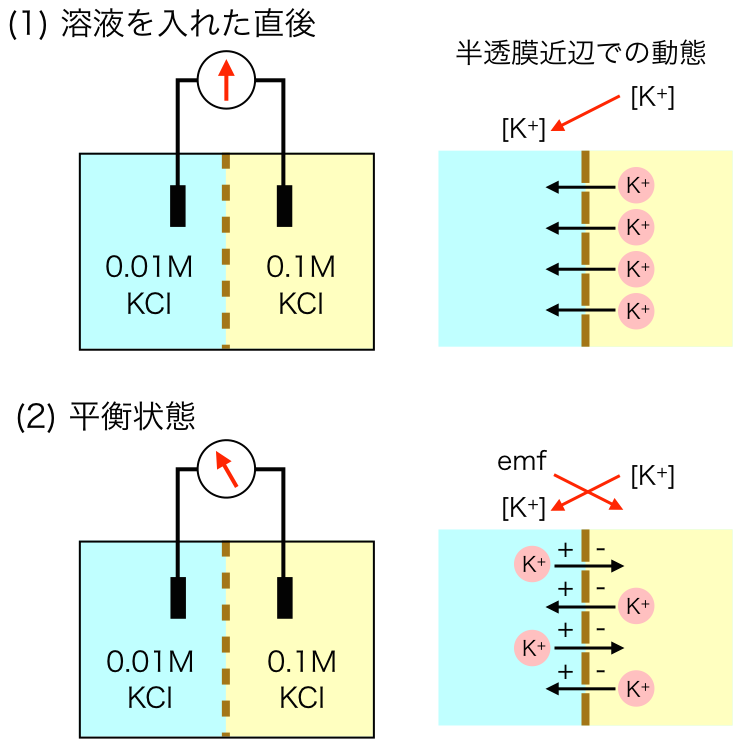
図3 K+イオンの電気化学的平衡
(Eckert and Randall 3rd ed. Figure 5-11を元に作成)
じゃあ、この分極している部分って細胞膜(厚さ5nm)の周りのどのあたりまでか、っていうと案外書いてない。少なくともカンデル本6章にはなかった。私も知らなかったけどWikipediaの膜電位の項目に書いてあった。膜の近傍の2-3nmだそうだ。
このあたりを深掘りするためには膜のチャネルをひとつの抵抗として捉えるような近似では追いつかなくて、膜の中を一定の電場として近似する定電場理論や、さらにチャネル内のエネルギー勾配をモデル化したりする必要がある。このあたりについては標準生理学8edのp.64-65やニューロンの生物物理 第2版のp.22-25に記述があるけどネルンスト・プランクの式(微分方程式)を導入する必要が出てくる。
じつは今回のブログ記事の個人的な伏線は、さいきんラボのOBの方たちと医学部の生理学教育においてネルンストの式をどこまで説明するか、どうやって説明するか、という話をしたことだった。あのあと、自分ならどうするかと考えていた。たぶん、図を多用して定性的な話をするべきで、それでさらに興味を持った者に対してはネルンスト・プランクの式とかを教える、という順番になるのではないかと思った。
(たとえば、ガイトン生理学ではネルンストの式は EK = 61(mV) * log10([Kout]/[Kin]) という表記を使っていて、RT/ZFを見えないようにしてある。しかもTは37度にしてあって、生理学実験でよく使う25度を使ってなかった。このへんはかなり実際的であることを意識していると思った。)
「これでわかるニューロンの電気現象」は著者が岡山大学の理学部生物学科の2,3年生を対象とした講義を元にしたものということなので、医学部の生理学教育を考えるのに良い題材であるといえる。「これでわかるニューロンの電気現象」およびEckert and RandallのAnimal Physiologyでは「どのくらい膜の近傍に限局してイオンが分極しているか」を説明するのに、膜の容量が1uF/cm2が上限であるということを使って、分極しているKイオンの数が細胞内の全体のKイオンと比べると1千万分の1程度であること、つまり膜を透過するKイオンによって細胞内のKイオンの濃度が変わったりはしないという説明をしている。これはわかりやすい。というわけで「これでわかるニューロンの電気現象」は素晴らしいのでおすすめ。(<-回し者)
(ちなみに、この説明を理解してもらうためには等価回路の概念の導入が必要であるので、正確に説明しようするとそれはそれでたいへんだと思う。脂質二重膜の実態に沿った説明と、等価回路を用いた説明をどのようにうまく使い分けて説明するかは、静止膜電位にかぎらず、神経生理学の教育において重要なコツとなるだろう。)
さきほどはカンデル本5edのTable 6-1に文句をつけたけど、どのように説明すると理解しやすいか、という眼でカンデル本を読んでみるとじつにうまく書かれていることがわかる。つか今回これ書くために読んでみて感服した。
カンデル本5edの6章では、Eckert and RandallのAnimal PhysiologyでのKイオンの半透膜の例を持ってくる代わりに、グリア細胞がほぼKイオンのleakチャネルだけである話を持ってきて、ネルンストの式の平衡状態を説明しておく。
そこからの差分として神経細胞の場合を説明している。神経細胞はKチャネルとNaチャネルからなるものと単純化しておく。グリア細胞ではKチャネルのみだった脂質二重膜にNaイオンのチャネルを付加すると、Naイオンが流入するようになってやがて膜間の電流の流れは0になるけど、ゆっくりとKイオンの流出とNaイオンの流入が起こる定常状態(平衡ではない)になってしまう。つまりそのまま放っておくとイオンの濃度勾配はなくなってしまう。それをせき止めるためにエネルギー使ってNa-Kポンプを回す話が導入されている。なるほどこれはステップバイステップでうまく説明してある。
今回この辺りの教科書を読みながら自分だったらどう説明するか考えていたんだけど、まったくゼロから説明するためには、「まず濃度勾配のない海水に脂質二重膜で壁を作って、Na-Kポンプを作って濃度勾配を作って、それからKチャネルを付加して」みたいなことを考えてた。でもこれはややこしくなりそう。カンデル方式のほうがすっきりしてる。
カンデル本ではその後でNa-Kポンプが静止膜電位を数mVほどnegative方向に押し下げるということも説明してある。これもKチャネルだけでネルンストの式を説明した後ならばすぐに理解できることなのでわかりやすい。ということでカンデル本おすすめ。と書くのも烏滸(うーろん)がましいが、あらためてその素晴らしさを実感したのだった。
(じつはこれらの教科書では触れられていなかったが、ニューロンの生物物理 第2版では、細胞内には有機イオン(A-)があってこれは膜の外には出られないのでドナン平衡を考慮する必要があって、これによってドナン浸透圧が説明できるということが書いてある。これは心臓の生理学に関わってくる。)
お勧めエントリ
- 細胞外電極はなにを見ているか(1) 20080727 (2) リニューアル版 20081107
- 総説 長期記憶の脳内メカニズム 20100909
- 駒場講義2013 「意識の科学的研究 - 盲視を起点に」20130626
- 駒場講義2012レジメ 意識と注意の脳内メカニズム(1) 注意 20121010 (2) 意識 20121011
- 視覚、注意、言語で3*2の背側、腹側経路説 20140119
- 脳科学辞典の項目書いた 「盲視」 20130407
- 脳科学辞典の項目書いた 「気づき」 20130228
- 脳科学辞典の項目書いた 「サリエンシー」 20121224
- 脳科学辞典の項目書いた 「マイクロサッケード」 20121227
- 盲視でおこる「なにかあるかんじ」 20110126
- DKL色空間についてまとめ 20090113
- 科学基礎論学会 秋の研究例会 ワークショップ「意識の神経科学と神経現象学」レジメ 20131102
- ギャラガー&ザハヴィ『現象学的な心』合評会レジメ 20130628
- Marrのrepresentationとprocessをベイトソン流に解釈する (1) 20100317 (2) 20100317
- 半側空間無視と同名半盲とは区別できるか?(1) 20080220 (2) 半側空間無視の原因部位は? 20080221
- MarrのVisionの最初と最後だけを読む 20071213