[月別過去ログ] 2013年04月
« 2013年03月 | 最新のページに戻る | 2013年05月 »2013年04月25日
■ さうして、このごろ 20121129
長女は夜更かしして手芸かなんかやってる。わたしは布団に入ってスライド作ってる。次男は寝相が悪いのでそっと押しやる。こうやって夜が更けてゆく。
ラップトップのACアダプタを家に忘れてきてしまった。なんど同じ過ちを繰り返すというのだろう。そう、これは習慣を変えるためにはどうすればよいかという、太古の昔よりわれわれ人間につきまとう構造的問題なのだ。(<-さっさと家とラボ両方にACアダプタを買って設置しておけ)
「止(と)める」「止(や)める」問題シリーズ、「床(ゆか)」か「床(とこ)」かってのもあったか。「川床料理」とか。こういうのが気になる。(<-人生もっと大事なことがあるからそういうことを気にするべき)
「涙は心の汗というがの、あれは逆での、体を動かすと体が悲しみ、涙をたくさん流すというわけでの」 (<-いんちき聞き取り調査風)
総武線の車内広告で「gabaだけは許す つかかなり認めてる」みたいな文面があって、みんないろいろたいへんなんだな、と世界に向けてなんだか優しい気持ちになった。
次男と城西高校まで「ふれ愛ときめきフェスティバル」に行ってきた。グレート家康公「葵」武将隊が来ていて、チャンバラが好きな次男は見入っていた。曲調が仮面ライダー/スーパー戦隊だったんで、なんか通底するものがあるんだなと面白かった。じっさい次男は「侍戦隊シンケンジャー」が好きだった。
木工ミニカー競争で惨敗したのが悔しかったらしく、帰りの車でずっと敗因を探していた。タイヤとシャフトの大きさが合わなくて直進性が悪かったんだよと教えてやった。案外細かく理知的であった。
図書館に本を返しに行く車で、iTunesの選曲からエレカシの「歴史」が流れてきた。これまでに幾度となく聴きながら歌ってきた曲だが、今日初めて歌詞の意味を実感して、涙が止まらなくなった。
「栄達がのぞめなくなる」「肩の荷が降りたのだろうか」「死に様こそが生き様だ」こういった歌詞をまあ、意味は分かったつもりいたし、「死に場所を見つけるんだ」という言葉には共感していた。それでも、ちょっと他人事みたいに聴いていて、森鴎外を出してくるところにはファニーささえ感じていた。
でも、まあそういうんじゃないんだなってことは分かったし、なんというか浄化された思いだった。
「世界を火の海にする」なんて言われても、心に響かない。ならば「世界を火の海にする」とはどんなことか、時間をかけて想像してみることにしよう。そこは「悲しみで涙が涸れた」り「三十年来の復讐を果た」したりということのない世界だ。
「どこ吹く風」ってあらためていいフレーズだな。自然に日常会話の中に忍び込ませてみたい。
「この窓から見える矢作川へと沈む夕日を見ることが出来さえすれば、僕は楽園にいるのとおなじなんだ。」
「ちくしょー、おれはまだやれるぞー」(<-不可視境界線のむこうに引きずり込まれる様子)
次男とだらだら遊んでる。「うらにわにはにわ、にわにはにわ、にわとりがいる」を理解させることに成功した。つぎは「わたしたわしわたしたわ」をどのくらい速い喋られるか特訓。
次男がごっこ遊びで「これがにくであるとします」と仮定を持ちだしたのでなんだか面白くなった。
次男がずっと「わたしたわしわたしたわ」で替え歌作ってる。プーさんみたいで楽しいので静観している。
次男を連れて、刈谷市中央児童館「はばたき」まで車で40分かけて行ってきた。卓球とバスケットボール。
次男はフォアはなかなか鋭いのを返すのでけっこう試合が成り立つ(10対5とか)のだが、バックハンドを攻略したいらしく、今日はなるたけバックに玉を集めるようにと要求。向上心のあるやつだ。
いつの日か、いろいろ回り道はしたがやってきたことは間違ってはいなかったと言えるような日が来たらよいと思うのだが、どちらにしろそれを認めるのも認めさせるのも俺自身だ。いまはまだ人生を語らず、ただ生き延びていることを根拠に、まだすべてが終わったわけではないと言いきかせるしかない。
2013年04月21日
■ NBNI2012で発表してきました
2012/11/22
トーク終了した。FAQの「じゃあ色はどうやってV1なしで処理できるの?」はスライドを用意していたが、そこへの質問は来なかった。「じゃあサリエンシーマップはSCにあるの?」という質問に関してはvalue-basedとかがLIPにあったりとかいろんなのが分散してるのでは、って答えた。
山口陽子さんからは「なんでV1が意識に重要なの?」という質問をいただいたので、V1じたいにneural correlate of consciousnessがあるわけではないらしい、ということを答えておいたのだけど、もっと端的にventral pathwayへの経路をブロックするから、と明示的にGoodale and Milnerに従ったラインで言った方が伝わったなとあとで思った。
Perceptual learningやっているひとから、free-viewing taskで見ているのはrecovery trainingによるperceptual learningの結果ではないか?と質問されて、でもmotion stimuliとかは出したことがないのでそれでは説明できないよ、って答えた。
でもそのときにひらめいたけど、たしかにrecovery trainingではmotionは使ってないけど、luminanceでdefineされた刺激をずっと使ってきた。Lesioned monkeyではluminance saliencyの寄与がnormal monkeyよりも高い。
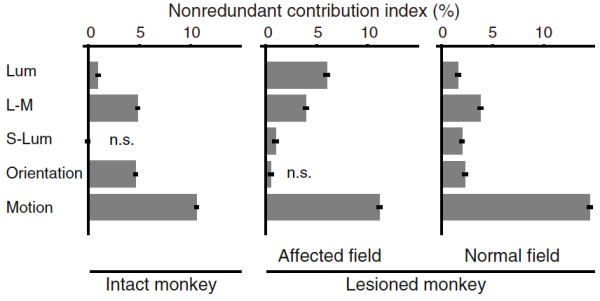
[Current Biology論文のFigure 4より]
いままではこれを説明することが出来なかったけど、彼の質問を敷延すれば、recovery trainingで輝度で定義された刺激でovertrainしたことによって輝度でdefineされたsaliencyが強化されている、もしくはsaliency mapとしていろんなfeatureを足し合わせる際にtop-down的にluminance saliencyの重みを高くしている(もしくはrewardによるvalue-based saliencyが高くなっている)というふうに説明できるんではないかと考えた。
つまり、この条件以外(normal monkeyおよびnormal field of lesioned monkey)では輝度サリエンシーのvalueは低いと言える。完全にspeculationだけど、featureごとの寄与の大きさにつじつまを合わせる説明が出来たのでちょっとこの解析に自信を持った。
(もともと、orientationサリエンシーが0になった上でカラーサリエンシーが0でなくて、色弁別課題ができる、というふうにconsistencyが得られたのでそれでやっと自信を持つことができたし、レビューワを説得できたのだが、それが自分の中で少し補強された感じ。)
ちなみに輝度の寄与がなぜ高い買って聞かれたことはほとんど無かったと思う。じつのところ、この寄与インデックスは絶対値にはあまり意味がない。(full modelでのAUC - leave-one-out modelのAUCだから、%で書いているけど合計しても100%にはならない)
2012/12/6
高等研カンファレンス終了。下條先生はいろんな話を入れながら、social interactionとcross modalを中心にという感じで楽しかった。Changiziの「三色性は顔色を見るために発達した説」の話もしてた。
そこで思ったのだけれども、わたしのfree-viewing実験では、normal monkeyではB-Y (konio)がsaliencyに使われない。(寄与が0%。) かえってblindsight monkeyのほうがB-Y使う。これはやっぱ、R-Gはsaliencyとして使っているのだけれども、B-Yは機能違うんじゃないかとか思った。
- / ツイートする
- / 投稿日: 2013年04月21日
- / カテゴリー: [Saliencyと眼球運動]
- / Edit(管理者用)
2013年04月15日
■ ひとりツール・ド・三河湖・チャレンジコース行ってきた
そんなこんなでまた自転車再開してるんだけど、1時間から1時間半くらいかけて20-30kmくらい走ってる。次の日が期限の論文も無事提出できたので 今日はまるまる一日空いた。そんなわけでひさびさに遠出することにした。
一つの可能性は、浜名湖一周コースで80kmくらい。これは以前にやったことがある。あともうひとつは、くらがり渓谷経由でつくで手作り村までの往復のコース。これはツールド三河湖のチャレンジコースと同じ。けっきょく後者にした。
行ってきた。トータルの走行距離 68.25km、経過時間202分。平均速度 20.23km/h。(runkeeperへのリンク)
行きは緩やかな上り坂で、26kmで250mの高さを進み、くらがり渓谷まで辿りつく。

くらがり渓谷から、これから登る道を写したところ。峠を攻め終わったオートバイが通りかかっている。
くらがり渓谷からつくで手作り村までは6kmで300m登るきっついコース。こういうのは三河湖スカイライン以来。ギアをいちばん軽いのにして、なんとか一度も足を付かずに登り切った。いちばん遅いときは時速7キロ切ってた。ロードバイク及びオートバイの人がたくさんいて、さんざん追い抜かされた。

目的地である、つくで手作り村に到着。 五平餅食った。鹿カレーは食ってない。クリーム大福も買ったが、凍っていたので背中に入れて溶かして食べた。 いまにして思えば、開封せずにポケットに入れておいて、帰りに補給用に食べればよかった。
帰り道は下り。これが最高だった。そんなにスピード好きではないのだけれども、クネクネ道を時速30キロオーバーで飛ばすのは最高だった。位置エネルギーってすごい!これのためにまたこのルート行こうと心に誓った。ブレーキワイヤがゆるみぎみだったので出発前に調整しておいたのだが、大正解。
帰りは楽に漕いでくるつもりだったのだけど、くらがり渓谷まで降りてきた時点で全行程での平均時速が18km/hであることに気づいた。これはつまり、帰りを24km/hくらいでキープできれば平均時速が20km/hを越えることを意味している。新しい目標が出来たのでちゃんと漕ぐことにした。
根詰めていたら、いつの間にか心拍数が165bpmくらいになっていた。これはまずい。行きの登り道でさえも155bpmくらいだったのに。信号で停まった2カ所で10分ずつくらい休んで、心拍が<120bpmになるのを待ってから再スタート。
ゆったりとした下りが続くのでスピードが出るはずだが、逆風で思ったほどスピードが出ない。思えば行きのゆっくりとした登りのときは追い風だったのだろう。まったく気づかなかった。追い風のときは気づかないけど向かい風のときは気づくものだ。まるで人生のように。(<-なんかうまいこと言った風)
けっきょく田舎道を降りきって、国1まで出たあたりで平均時速20キロ越えを達成。だいたい満足した。朝10時に出発して、午後4時前にはすべてが終了。妥当な線。もっと先の三河湖あたりまで遠出するとしたら、二時間早く出発することが必要そうだ。
2013年04月11日
■ レスコーラシンポのために予習
2012/11/8
レスコーラシンポのために予習としてRWモデルの理解(<-そこから?)。読んでて楽しいのは、salienceとかsurpriseとか、わたしがこれまで聞いてきたようなキーワードが出てくること。
すると、ここで言ってるsurpriseをIttiのベイジアン・サプライズで置き換えらんないか?なんてことを考える。連合学習理論の本質がなんなのかまだ私には分かっていないのだけれども、もし究極には「連合強度の上げ下げ」を扱うかどうかであるなら、ベイジアンな連合学習理論もありうる。
つまり、複数のCS1, CS2の出現頻度とそれぞれでのcontingency P(US|CS1), P(US|~CS1), P(US|CS2), P(US|~CS2)を逐一「推定」して行動判断をする過程があれば、そのときの「連合強度」を計算することが出来るだろう。
そのときの更新則がサプライズ(連合強度の次元での引き算)になっているのがRWモデルで、この更新則にベイジアン・サプライズを使えないかってこと。(ベイジアン・サプライズ = posterior beliefとprior beliefとのKL距離)
"associative learning" bayesianでググってみたら、John K. KruschKeの論文がみつかった。("Doing Bayesian Data Analysis"の人)
あと、Fristonはどうかというと、本人筆頭の論文ではRW1972をSutton & Bartoと並べて言及するだけでそんな深いことを言ってない。共著ではCerebral Cortex 2009、 あとスライドでこのあたりに出てくる。
ともあれ、ここまで極端に振ってみると、ベイズ的になんでも全部取り込んで確率分布として表象しちゃう方策と、必要最小限のことだけ表象する方策(そこで「連合強度」を使うのが連合学習理論)とでそれぞれに学習をして行動を最適化をすることが可能だと言える。これで毎度のベイズ脳批判に到達する。
あとはどこまでそういった表象を持っているかを調べるというempiricalな問題になる。
ここまで考えると、データドリヴンでの連合学習モデルの形成とかも思いつく。つまり、CS1, CS2, US1, US2くらいのパラメータがあって、連合学習で知られているあらゆるスケジュールは上記パラメータ間の時間的関係として記述される。
それで、これらのパラメータとスケジュールを振った上で、CR1, CR2のデータ(それぞれUS1, US2にたいするもの)を収集する。これらのパラメータとスケジュールとCRの関係をうまく説明するモデルを機械学習で作ってやる。(<-出たっ機械学習信仰)
そのとき「連合強度」とよべるような潜在変数が出てくるかを問う。もしくは既存のモデルでの連合強度をモデルに入れてやるとモデルの自由度が効率的に下げられるかどうか検証するとか。
CSもUSもなんらかの刺激だから本質的には差がないので(じっさいSOPではそうなっている)、もう少しパラメータ数を減らしてやることは出来るかもしれない。
一般化すれば、P(R1(t),R2(t),...) = f(P(S1(t),S1(t-1),...,S2(t),S2(t-1),...))となるわけだから。
こういうのもどっかにあるか。ググってみたが、data-driven "associative learning"では見つからなかった。
いろいろ外している気はするが、それでもとにかく書いてみた。
明日の予習用にと"Pavlovian Conditioning: It's Not What You Think It"を読んだ。よく構造化されているし、かなりかみ砕いて書いてあるので読みやすい。
まとめておくと、1988年の段階でも、教科書的なPavlovian Conditioningの説明というのは連合学習理論の進歩をちゃんと反映していない。
1) temporal contiguityはconditioningの必要条件でもなければ、充分条件でもない。(P(US|CS) vs. P(US|~CS)の関係やblockingの例を出して、動物含むわれわれ有機体はinformation seekerなのだと強調する)
2) 単純な刺激CSとUSの間を連合するものだけではない。RHというcompoundとR,H単体とを別々にconditioningする例を出して、この連合によって単純なものから複雑な概念を作ることが出来ることを強調している。(これがたぶんHumeに繋がる)
3) CSはUSが引き起こした応答だけを誘導するわけではない。たとえばCSがlocalizedであるかdiffuseであるかによって引き起こす応答は違いうる。ゆえに、あるCSによってUSで見える応答が起こさなかくても、べつの応答に反映する可能性がある。
ここまで。1)に関しては「学習心理学における古典的条件づけの理論」で読んでだいたい分かっていたが、それ以外にももっといろんな論点があって面白かった。たとえばorganismのexisting structureの重要性なんてのは、すごくエコロジカルだ。
なぜ急にこの分野に勉強しているかというと、連合学習理論はじつはすごく表象主義的なので、反表象主義的なsensorimotor理論とぶつけて考える意義があると思うから。一方で、徹底的に擬人化を避けるという意味では両者は共通していて、認知神経科学とはちょっと違ったところにいるようだ。
明日は"Contemporary Study of Pavlovian Conditioning"を読んでみることにしよう。
自分の興味の向き方を考えてみると、現象学的経験自体には興味があるのだけれども、心の理論とかミラーニューロンとかはどうも擬人的すぎてなじめない。身体性には興味があるが、社会性はよく分からない。あと、いつまで経っても「言語」に興味が向かない。良くも悪くもいまのところそんなかんじ。
つまり、意識経験は重要なんだけど、人間的であること(概念、言語、…)にはなぜかあまり惹かれない。非人間的な意識科学者。ってまさに自分がやってきたことか。
2012/11/9
.@kosukesa おつかれさまでした。新幹線の時間があったのでレスコーラの若手へのメッセージを聞いたところまでで失礼させていただきました。
レスコーラのトークは「compound stimuliを使うとあれも出来るこれも出来る」みたいな話で、神経科学者としては難易度高かった。昨日読んだAmerican Psychologistをもし読んでいなかったら、「そもそもABはAB->USなのかA->US, B->USなのか」ってところからつまずくところだった。
レスコーラのメッセージでいいなと思ったのは"Identify critical difference among accounts"(そのためにきっちり統制実験しとけよ)ってのと"Find the improbable prediction"(インパクトのある仕事になる)の二つ。
こんかいのしゅっちょうでは、アイパッドミニをじっさいにてにとってかくにんすることができたのがよかったことです(<-小学生の感想文)
2013年04月07日
■ 脳科学辞典「盲視」の項目書いた
脳科学辞典の「盲視」の項目を書いた。査読されるまえの原稿をブログ用の記事として活用してみる。
盲視
英:blindsight
類語・同義語:
盲視とは、第一次視覚野(primary visual cortex: V1)が損傷した患者において、現象的な視覚意識がない(phenomenal blindness)にもかかわらず見られる、視覚誘導性の自発的な反応のことを指す。盲視という現象は視覚情報の処理(光点の位置を当てる)と現象的な視覚意識(光点が眼前に見えたという経験をする)とが乖離しうること、そしてそれらがべつの脳部位で処理されているということを示している。ヒトだけではなく、マカクザルにおいても盲視と同様な行動が見られる。盲視に関わる脳部位としては上丘を経由するとする説と外側膝状体を経由するとする説とがある。盲視の能力の発現には機能回復トレーニングと可塑性が関与していることを示唆する報告が複数ある。
盲視とは
盲視とは、第一次視覚野(primary visual cortex: V1)が損傷した患者において、現象的な視覚意識がない(phenomenal blindness)にもかかわらず見られる、視覚誘導性の自発的な反応のことを指す[1]。
V1は大脳皮質での視覚情報が最初に入ってくる領域であり、左右の半球でそれぞれ右左半分ずつの視野の情報を処理している。たとえば左側のV1全体が損傷すると、左右の眼ともに右半分の視野が見えなくなる。このような症状は同名半盲と呼ばれる。盲視はそのような患者の一部でのみ見られる。
盲視という現象は視覚情報の処理(光点の位置を当てる)と現象的な視覚意識(光点が眼前に見えたという経験をする)とが乖離しうること、そしてそれらがべつの脳部位で処理されているということを示している。気づきの項目も参照。
ヒトでのV1損傷後の残存視覚については1973年のPoppelらの仕事[2]によって最初に報告された。ひきつづきWeiskrantzらがBrain誌に詳細な報告を行い[3]、"blindsight" (盲視)と呼ばれるようになった[4]。
歴史的にいえば、V1損傷後の残存視覚についてはじつはヒトでの知見の前にすでにサルでの知見がWeiskrantzらの研究グループから報告されていた[5]。しかし、厳密な意味で盲視の存在を証明するためには、「残存視覚があること」を証明するだけでなく、「現象的な視覚意識がない」ことを証明しなければならない。これは言語報告を使えないサルなどの動物の場合には原理的な問題となる。ヒト盲視で見られる現象と同様な行動の乖離を示した実験は、Weiskrantzの同僚であるAlan Coweyらによって1995年に報告された[6]。
包括的なレビューとしてはPetra StoerigとAlan CoweyによるBrain 1997[7]がある。また、日本語で読むことができる総説としては[8][9]などがある。
盲視で出来ること、出来ないこと(1) ヒト
ヒト盲視患者では動き刺激の方向弁別の成績は非常に高い[10]。一方で、線分の方位弁別の成績は偶然のレベルに留まっている[11]。
グレーティング刺激の検出課題において、輝度コントラストに対する閾値は通常と比べて上昇している[12]。また、空間周波数の影響を調べた論文[13]によると、盲視では空間周波数の高い成分(> 4 cycles / deg)への感度が落ちている。
色情報の検出、弁別は可能であるとする報告[14][15]がある。しかし一方で、V1損傷または半球皮質切除によるヒト盲視患者では青-黄の色拮抗チャネル(koniocellular経路)の刺激を検出することが出来ないという報告もある[16][17][18]。
ポズナー課題による注意課題において、盲視患者は手がかり刺激の情報を使って見えない視覚刺激に対する応答潜時が短くなるなどの注意の効果が見られることが報告されている[19]。
顔の表情の弁別においては、二択で偶然以上の成績で正解した[20]。このような表情の認知は"affective blindsight"と呼ばれる。一方で、おなじ患者は誰の顔であるか(identity)の弁別では偶然のレベルの成績であった。
盲視で出来ること、出来ないこと(2) 動物モデル
この項では、マカクザルの片側のV1を損傷させた盲視動物モデルでの知見をまとめる。
視覚誘導性サッカード課題において、損傷部位に対応した視野に提示した視覚標的に向けてサッカードすることが可能であることが示された[21]。また、レバープレス課題によって提示された刺激を検出することも可能であった。
視覚誘導性のリーチング課題によって、ディプレーに提示された視覚刺激の位置を二択で正しく選択することが可能だった[6]。一方で、視覚刺激があるか無いかを報告させる課題においては、視覚刺激が提示されていても、視覚刺激が提示されていないことを示す選択肢を選んだ。
視覚誘導性サッカード課題において、損傷視野に提示した視覚標的の輝度コントラストに対する閾値は正常視野と比べて上昇していた[22]。また、サッカードの終止点は不正確であり、軌道も正常視野へのサッカードと比べてより直線的になっていた。このことはV1損傷が視覚だけでなく運動コントロールにも影響を与えていることが示唆している。また、応答潜時は分布が狭くなっており、計算論的解析から、V1損傷が意志決定の過程にも影響を与えていることが示唆している。
記憶誘導性サッカード課題を用いて、盲視モデル動物が見えていない部分に提示された視覚刺激の位置を2秒間記憶することが出来るかどうかを検証したところ、盲視モデル動物はこの課題を90%以上の成績で行うことができた[23]。また、注意誘因課題において、キュー刺激を事前に提示することによって視覚誘導性サッカードの応答潜時は短くなった[24]。これらのことは盲視の動物モデルでは反射的な視覚情報処理だけではなく、高次認知機能も遂行可能であることを示唆している。
盲視モデル動物がムービークリップを受動的に見ている間の眼の動きをサリエンシー計算論モデルによって解析することによって、盲視で利用可能な情報処理のチャンネルを網羅的に調べた報告がある[25]。盲視モデル動物では「輝度」「赤-緑」「青-黄」「動き」の情報は利用できるが、「傾き」の情報は利用できなかった。同じ動物に等輝度色刺激を提示して刺激を検出できるかどうか検証したところ、赤-緑、青-黄どちらの刺激ともに偶然より高い成績で検出できることが判明した。この結果は盲視モデル動物で色情報の処理が出来るとするこれまでの報告[15]と整合的だった。
盲視の脳内メカニズム(1) 解剖学
盲視で網膜からV1を経由せずに視覚情報を伝えるルートとしては、解剖学的には以下の可能性がある。
- 網膜から上丘への入力が直接眼球運動などの行動を引き起こす[1]。
- 網膜から上丘への入力が視床枕を経由してextrastriate cortexに到達する[26][27]。
- 網膜から外側膝状体(lateral geniculate nucleus: (LGN))への入力が直接extrastriate cortexに到達する[28][29]。
LGNへの入力には網膜からの直接的な投射だけではなく間接的なものもある。LGNへの間接的な視覚入力としては、上丘のうち網膜から直接入力を受ける上丘浅層(stratum griseum superficiale: SGS)からLGNへの投射があることが知られている。しかもこのような投射はヒト、マカクザルを含むほ乳類のさまざまな種で保存されている[30]。よって、LGNには、網膜からSGSを経由した情報が入力している可能性がある。
マカクザルでMT野にrabies virusを注入すると逆行性および多シナプス性に上丘が標識される[31]。このことは上丘からMT野までシナプス一つを介して入力していることを示している。上丘からMT野への間接的投射には、SGS 2b層 -> 視床枕 -> MT野という経路と、SGS 2b層 -> LGN -> MT野という経路の二つの可能性がある。はたして、ウイルスによって標識されたのはSGS 2b層であった。このことは上丘からMT野への投射は視床枕を経由していることを示唆している。よって、上丘からLGNを経由してMT野へ情報が入るという可能性は解剖学の知見からは否定的である。
盲視の脳内メカニズム(2) 生理学
V1損傷後のマカクザルのMT野ニューロンの視覚応答は、発火頻度は小さくなっているものの方向選択性はほぼ正常時と同じ程度に保持されていた[32]。一方で、V1と上丘とを両方損傷させた場合にはMT野の応答は完全に消失した[27]。このことはV1損傷後のMT野ニューロンの応答が上丘を経由するものであることを示唆しており、LGNから直接MT野へ入力する経路によっては説明できない。
マカクザルを盲視の動物モデルとして用いた研究では、上丘を薬理学的に抑制すると、視覚弁別能力が消失する[21][33]。この実験結果は盲視が上丘を経由する回路で処理されている可能性を示唆している。一方で、V1の部分的除去のあとでも、視覚刺激への応答がV2やMT野で見られることがサルでのfMRIを用いた研究から報告されている[34]。この状況では、LGNを薬理学的に抑制すると盲視が消失する。この実験結果は盲視がLGNを経由する回路で処理されている可能性を示唆している。この二つの説のどちらが正しいかを実証するためには、同じ動物で上丘、LGNそれぞれを抑制する実験をする必要がある。
マカクザルを盲視の動物モデルとして用いた研究では、V1の切除後の上丘からニューロン活動を記録したものがある。損傷したV1と同側の上丘には、視覚刺激に応答するもの、サッカードの実行時に活動するものが見いだされた[23]。つまり、V1損傷の後も上丘は機能している。さらに、記憶誘導性サッカード課題遂行中の上丘のニューロン活動を記録したところ、上丘は短期空間記憶を保持している期間のあいだ持続的に活動していた。このことはV1の損傷からの機能回復によって、上丘が普段は行っていない、空間的短期記憶の機能を担うようになったということを示唆している[23]。
機能回復トレーニングと可塑性の寄与
盲視の能力の発現には機能回復トレーニングと可塑性が関与していることを示唆する報告が複数ある。
Sahraieらの研究[12]では、視覚皮質損傷患者を被験者として視覚弁別のトレーニングを行った。視覚刺激が試行の期間1または期間2のどちらに提示されたかを被験者は答える。このようなトレーニングを被験者は自宅で継続して行うことでその成績は数ヶ月をかけて向上した。また、Huxlinらによる報告[35]では、ランダムドットモーション刺激の方向弁別のトレーニングを行ったところ、9-18ヶ月後には正常レベルに近いところまで感度が向上していた。これらふたつの研究での被験者は成人であり、脳損傷を受けてから年月が経っている。よって、これらの研究は、成人の脳でも大規模な構造的な変化によって機能回復が起こっている可能性を示唆している。
マカクザルを動物モデルとして用いた研究では、機能回復トレーニングとして視覚誘導性サッカード課題を用いて成績の時間経過を調べたところ、術後1週間では、損傷の反対側の視野へのサッカードは上下の2カ所を弁別できなくなっていた。継続的にトレーニングを行ったところ、およそ8週間程度で損傷視野の成績はほぼ正常視野と同等のレベルまで回復した。つまり、動物モデルにおいても数ヶ月の機能回復トレーニングによって、盲視の能力が回復することが明らかになった[22]。前述した、上丘の神経活動が損傷と同側では正常側と変わっているという報告[23]も、V1損傷後に大規模な構造的変化が起きた結果である可能性がある。
拡散テンソルイメージング(DTI)を用いることによって、盲視の患者では脳損傷後に投射経路の可塑的変化が起こっていることが示唆されている。たとえばLGNからMT野への結合がより強くなっている[36]。また、上丘から視床枕を経由して扁桃体へと入力する結合がより強くなっている[37]。また、半球皮質切除を受けた患者のうち盲視の能力を持つ患者では、通常では見られないような、切除側の上丘から反対側の大脳皮質へと投射する経路が同定されている[38]。
盲視での意識経験
盲視の被験者は視覚刺激に対してまったく意識経験がないわけではないらしい。たとえば有名な盲視被験者のGY氏は、視覚刺激の強度が高いときにはしばしば「何かある感じ」がすると報告する[39]。しかしそれはいわゆる視覚経験とは違うらしい。たとえばGY氏はその感覚について「黒い影が黒い背景上を動いている感じ」と表現する(ただし、この表現はあくまで比喩であることを強調している)。Weiskrantzはこのような盲視をtypeII盲視と呼んで、このような意識経験を全く持たないtype I 盲視と区別している[1]。
いっぽうでZekiはこのような感覚は視覚経験の一種であり、Riddoch症候群として捉えるべきであると主張している[39]。Riddoch現象とは、V1を損傷した患者で、静止した物体はまったく見えないのに対して、動いているものに関しては感知できる現象のことを指す[40]。
ほかの感覚でも盲視に対応したものはあるか?
視覚で盲視があるのと同じように、ほかの感覚でも盲視に対応したものがあるのだろうか? 以下の論文ではそのような症例があることが報告されている。
- 触覚での症例 ("Tactile ananogue of blind sight") [41]
- 聴覚での症例 ("Deaf hearing") [42]
- 嗅覚での症例 ("Blind smell") [43]
関連項目
参考文献
- ↑ 1.0 1.1 1.2 L. Weiskrantz
Blindsight: a case study spanning 35 years and new developments
Oxford University Press.: 2009 - ↑
E Poppel, R Held, D Frost
Leter: Residual visual function after brain wounds involving the central visual pathways in man.
Nature: 1973, 243(5405);295-6 [PubMed:4774871] [WorldCat.org]
- ↑
L Weiskrantz, E K Warrington, M D Sanders, J Marshall
Visual capacity in the hemianopic field following a restricted occipital ablation.
Brain: 1974, 97(4);709-28 [PubMed:4434190] [WorldCat.org]
- ↑
M D Sanders, E K Warrington, J Marshall, L Wieskrantz
"Blindsight": Vision in a field defect.
Lancet: 1974, 1(7860);707-8 [PubMed:4132425] [WorldCat.org]
- ↑
N K Humphrey, L Weiskrantz
Vision in monkeys after removal of the striate cortex.
Nature: 1967, 215(5101);595-7 [PubMed:4963569] [WorldCat.org]
- ↑ 6.0 6.1
A Cowey, P Stoerig
Blindsight in monkeys.
Nature: 1995, 373(6511);247-9 [PubMed:7816139] [WorldCat.org] [DOI]
- ↑
P Stoerig, A Cowey
Blindsight in man and monkey.
Brain: 1997, 120 ( Pt 3)();535-59 [PubMed:9126063] [WorldCat.org]
- ↑ 吉田 正俊
盲視の神経科学
Clinical Neuroscience: 30(8): 955-957 - ↑ 吉田 正俊
盲視の神経機構
BRAIN and NERVE: (In press) - ↑
L Weiskrantz, J L Barbur, A Sahraie
Parameters affecting conscious versus unconscious visual discrimination with damage to the visual cortex (V1).
Proc. Natl. Acad. Sci. U.S.A.: 1995, 92(13);6122-6 [PubMed:7597090] [WorldCat.org]
- ↑
A B Morland, J A Ogilvie, K H Ruddock, J R Wright
Orientation discrimination is impaired in the absence of the striate cortical contribution to human vision.
Proc. Biol. Sci.: 1996, 263(1370);633-40 [PubMed:8677261] [WorldCat.org] [DOI]
- ↑ 12.0 12.1
Arash Sahraie, Ceri T Trevethan, Mary Joan MacLeod, Alison D Murray, John A Olson, Lawrence Weiskrantz
Increased sensitivity after repeated stimulation of residual spatial channels in blindsight.
Proc. Natl. Acad. Sci. U.S.A.: 2006, 103(40);14971-6 [PubMed:17000999] [WorldCat.org] [DOI]
- ↑
Arash Sahraie, Ceri T Trevethan, Lawrence Weiskrantz, John Olson, Mary Joan MacLeod, Alison D Murray, Roelf S Dijkhuizen, Carl Counsell, Richard Coleman
Spatial channels of visual processing in cortical blindness.
Eur. J. Neurosci.: 2003, 18(5);1189-96 [PubMed:12956717] [WorldCat.org]
- ↑
P J Brent, C Kennard, K H Ruddock
Residual colour vision in a human hemianope: spectral responses and colour discrimination.
Proc. Biol. Sci.: 1994, 256(1347);219-25 [PubMed:8058800] [WorldCat.org] [DOI]
- ↑ 15.0 15.1
A Cowey, P Stoerig
Detection and discrimination of chromatic targets in hemianopic macaque monkeys and humans.
Eur. J. Neurosci.: 2001, 14(8);1320-30 [PubMed:11703461] [WorldCat.org]
- ↑
Marco Tamietto, Franco Cauda, Luca Latini Corazzini, Silvia Savazzi, Carlo A Marzi, Rainer Goebel, Lawrence Weiskrantz, Beatrice de Gelder
Collicular vision guides nonconscious behavior.
J Cogn Neurosci: 2010, 22(5);888-902 [PubMed:19320547] [WorldCat.org] [DOI]
- ↑
Petroc Sumner, Thofique Adamjee, J D Mollon
Signals invisible to the collicular and magnocellular pathways can capture visual attention.
Curr. Biol.: 2002, 12(15);1312-6 [PubMed:12176359] [WorldCat.org]
- ↑
Sandra E Leh, Kathy T Mullen, Alain Ptito
Absence of S-cone input in human blindsight following hemispherectomy.
Eur. J. Neurosci.: 2006, 24(10);2954-60 [PubMed:17156217] [WorldCat.org] [DOI]
- ↑
R W Kentridge, C A Heywood, L Weiskrantz
Attention without awareness in blindsight.
Proc. Biol. Sci.: 1999, 266(1430);1805-11 [PubMed:10518327] [WorldCat.org] [DOI]
- ↑
B de Gelder, J Vroomen, G Pourtois, L Weiskrantz
Non-conscious recognition of affect in the absence of striate cortex.
Neuroreport: 1999, 10(18);3759-63 [PubMed:10716205] [WorldCat.org]
- ↑ 21.0 21.1
C W Mohler, R H Wurtz
Role of striate cortex and superior colliculus in visual guidance of saccadic eye movements in monkeys.
J. Neurophysiol.: 1977, 40(1);74-94 [PubMed:401874] [WorldCat.org]
- ↑ 22.0 22.1
Masatoshi Yoshida, Kana Takaura, Rikako Kato, Takuro Ikeda, Tadashi Isa
Striate cortical lesions affect deliberate decision and control of saccade: implication for blindsight.
J. Neurosci.: 2008, 28(42);10517-30 [PubMed:18923028] [WorldCat.org] [DOI]
- ↑ 23.0 23.1 23.2 23.3
Kana Takaura, Masatoshi Yoshida, Tadashi Isa
Neural substrate of spatial memory in the superior colliculus after damage to the primary visual cortex.
J. Neurosci.: 2011, 31(11);4233-41 [PubMed:21411664] [WorldCat.org] [DOI]
- ↑
Takuro Ikeda, Masatoshi Yoshida, Tadashi Isa
Lesion of primary visual cortex in monkey impairs the inhibitory but not the facilitatory cueing effect on saccade.
J Cogn Neurosci: 2011, 23(5);1160-9 [PubMed:20521856] [WorldCat.org] [DOI]
- ↑
Masatoshi Yoshida, Laurent Itti, David J Berg, Takuro Ikeda, Rikako Kato, Kana Takaura, Brian J White, Douglas P Munoz, Tadashi Isa
Residual attention guidance in blindsight monkeys watching complex natural scenes.
Curr. Biol.: 2012, 22(15);1429-34 [PubMed:22748317] [WorldCat.org] [DOI]
- ↑
G S Brindley, P C Gautier-Smith, W Lewin
Cortical blindness and the functions of the non-geniculate fibres of the optic tracts.
J. Neurol. Neurosurg. Psychiatr.: 1969, 32(4);259-64 [PubMed:5807867] [WorldCat.org]
- ↑ 27.0 27.1
H R Rodman, C G Gross, T D Albright
Afferent basis of visual response properties in area MT of the macaque. II. Effects of superior colliculus removal.
J. Neurosci.: 1990, 10(4);1154-64 [PubMed:2329373] [WorldCat.org]
- ↑
S Zeki
The motion vision of the blind.
Neuroimage: 1995, 2(3);231-5 [PubMed:9343607] [WorldCat.org] [DOI]
- ↑
Lawrence C Sincich, Ken F Park, Melville J Wohlgemuth, Jonathan C Horton
Bypassing V1: a direct geniculate input to area MT.
Nat. Neurosci.: 2004, 7(10);1123-8 [PubMed:15378066] [WorldCat.org] [DOI]
- ↑
J K Harting, M F Huerta, T Hashikawa, D P van Lieshout
Projection of the mammalian superior colliculus upon the dorsal lateral geniculate nucleus: organization of tectogeniculate pathways in nineteen species.
J. Comp. Neurol.: 1991, 304(2);275-306 [PubMed:1707899] [WorldCat.org] [DOI]
- ↑
David C Lyon, Jonathan J Nassi, Edward M Callaway
A disynaptic relay from superior colliculus to dorsal stream visual cortex in macaque monkey.
Neuron: 2010, 65(2);270-9 [PubMed:20152132] [WorldCat.org] [DOI]
- ↑
H R Rodman, C G Gross, T D Albright
Afferent basis of visual response properties in area MT of the macaque. I. Effects of striate cortex removal.
J. Neurosci.: 1989, 9(6);2033-50 [PubMed:2723765] [WorldCat.org]
- ↑
Rikako Kato, Kana Takaura, Takuro Ikeda, Masatoshi Yoshida, Tadashi Isa
Contribution of the retino-tectal pathway to visually guided saccades after lesion of the primary visual cortex in monkeys.
Eur. J. Neurosci.: 2011, 33(11);1952-60 [PubMed:21645091] [WorldCat.org] [DOI]
- ↑
Michael C Schmid, Sylwia W Mrowka, Janita Turchi, Richard C Saunders, Melanie Wilke, Andrew J Peters, Frank Q Ye, David A Leopold
Blindsight depends on the lateral geniculate nucleus.
Nature: 2010, 466(7304);373-7 [PubMed:20574422] [WorldCat.org] [DOI]
- ↑
Krystel R Huxlin, Tim Martin, Kristin Kelly, Meghan Riley, Deborah I Friedman, W Scott Burgin, Mary Hayhoe
Perceptual relearning of complex visual motion after V1 damage in humans.
J. Neurosci.: 2009, 29(13);3981-91 [PubMed:19339594] [WorldCat.org] [DOI]
- ↑
Holly Bridge, Owen Thomas, Saâd Jbabdi, Alan Cowey
Changes in connectivity after visual cortical brain damage underlie altered visual function.
Brain: 2008, 131(Pt 6);1433-44 [PubMed:18469021] [WorldCat.org] [DOI]
- ↑
Marco Tamietto, Pim Pullens, Beatrice de Gelder, Lawrence Weiskrantz, Rainer Goebel
Subcortical connections to human amygdala and changes following destruction of the visual cortex.
Curr. Biol.: 2012, 22(15);1449-55 [PubMed:22748315] [WorldCat.org] [DOI]
- ↑
Sandra E Leh, Heidi Johansen-Berg, Alain Ptito
Unconscious vision: new insights into the neuronal correlate of blindsight using diffusion tractography.
Brain: 2006, 129(Pt 7);1822-32 [PubMed:16714319] [WorldCat.org] [DOI]
- ↑ 39.0 39.1
S Zeki, D H Ffytche
The Riddoch syndrome: insights into the neurobiology of conscious vision.
Brain: 1998, 121 ( Pt 1)();25-45 [PubMed:9549486] [WorldCat.org]
- ↑ G. Riddoch
Dissociation of visual perceptions due to occipital injuries, with especial reference to appreciation of movement
Brain: 1917; 40: 15–57 - ↑
J Paillard, F Michel, G Stelmach
Localization without content. A tactile analogue of 'blind sight'.
Arch. Neurol.: 1983, 40(9);548-51 [PubMed:6615285] [WorldCat.org]
- ↑
M M Garde, A Cowey
"Deaf hearing": unacknowledged detection of auditory stimuli in a patient with cerebral deafness.
Cortex: 2000, 36(1);71-80 [PubMed:10728898] [WorldCat.org]
- ↑
Gesualdo M Zucco, Massimo Prior, Giuseppe Sartori, Richard J Stevenson
Odour perception following bilateral damage to the olfactory bulbs: A possible case of blind smell.
Cortex: 2012; [PubMed:22578708] [WorldCat.org] [DOI]
- / ツイートする
- / 投稿日: 2013年04月07日
- / カテゴリー: [視覚的意識 (visual awareness)]
- / Edit(管理者用)
お勧めエントリ
- 細胞外電極はなにを見ているか(1) 20080727 (2) リニューアル版 20081107
- 総説 長期記憶の脳内メカニズム 20100909
- 駒場講義2013 「意識の科学的研究 - 盲視を起点に」20130626
- 駒場講義2012レジメ 意識と注意の脳内メカニズム(1) 注意 20121010 (2) 意識 20121011
- 視覚、注意、言語で3*2の背側、腹側経路説 20140119
- 脳科学辞典の項目書いた 「盲視」 20130407
- 脳科学辞典の項目書いた 「気づき」 20130228
- 脳科学辞典の項目書いた 「サリエンシー」 20121224
- 脳科学辞典の項目書いた 「マイクロサッケード」 20121227
- 盲視でおこる「なにかあるかんじ」 20110126
- DKL色空間についてまとめ 20090113
- 科学基礎論学会 秋の研究例会 ワークショップ「意識の神経科学と神経現象学」レジメ 20131102
- ギャラガー&ザハヴィ『現象学的な心』合評会レジメ 20130628
- Marrのrepresentationとprocessをベイトソン流に解釈する (1) 20100317 (2) 20100317
- 半側空間無視と同名半盲とは区別できるか?(1) 20080220 (2) 半側空間無視の原因部位は? 20080221
- MarrのVisionの最初と最後だけを読む 20071213