[月別過去ログ] 2012年12月
« 2012年11月 | 最新のページに戻る | 2013年01月 »2012年12月30日
■ さうして、このごろ 20120629
20120601
「また逢う日まで」を歌いたい。こればっかりはギター弾いて歌うより、マイクを通して絶唱したい。変態っぽく、粘っこく。
よく替え歌にして、「そのとき心は何かを放つだろう」(エクトプラズムかなんかが口からスポーンと飛び出してくるイメージ)、とか車の中で独りで歌っては噴いてたのですが、それもこれからは出来ないかと思うと、寂しいものです。
20120603
車ってのは、殺人兵器であり、人間の能力を超えていて、それでも乗るのをやめることが出来ない、悪魔の文明の利器だと思うのだけれど、それでも、ハチクロでデザイン事務所のイケメンが、山田にそば食いに行こうってまるで近所のそば屋にでも行くような言い方で誘いつつ、東京から蓼科まで中央道を飛ばして行った、という逸話を読んだときは、なんかその爽快感にやられちまった。
スピードに惹かれないのだな。MTB寄りの安物クロスバイクに乗っているけど、ロードとか乗りかえたいとは思わない。岡崎で時速30キロ出したら死ぬ。それよか、なんらかの方法で負荷かけて、時速15キロでいいから心拍140bpmで2hrキープできるほうが望ましい。山道登れってか。
20120604
なんか名残惜しかったので、ちと外へ歩きに行ったら、空気が気持ちよかった。コンビニでカルピスサワーとおにぎり買って、奈良井公園のあずまやで暗闇の中BB2Cでも見ながらゆっくりとするつもりだったのだけれども、2,3人のインド系の方々があずまやには先客としていた。
ブラジル系の人はけっこういるのだけれども、インド系の人は珍しい。研究所の人がここまで来るだろうか? ともあれ、異国の地で、でもものすごく安全で(車運転して珍走団にでも巻き込まれない限り)、誰もいない真っ暗な公園で楽しそうに話をしているのをわたしはちょっと離れたベンチで聞きながら、カルピスサワー飲み干して、ちと気分を高揚させて、空気が気持ちよくて、見上げたら月がほとんど満月で、そういえば明日は部分月食だったのを思い出して、そうしてうちに帰ってきた。俺のことをダメ人間だと思うやつがどれだけいたとしても、俺は生き延びてやるんだ。
先日子どもたちの運動会を見にきたお婆ちゃんズが回転寿司屋に連れて行ってくれたんだけど、そこで店員が注文を受けるたび「ガッテン承知」と返事するのを次男が気に入ったらしく、日常生活で活用している。ママが風呂に来なさい、と言えば「ガッテン承知」。ネタに走る精神がたいへん素晴らしい。
「ネコまっしぐら」の代わりに「ベコまっしぐら」っていう、東北弁に基づいた切れ味の鋭いフレーズを考えたんだけど、前にも同じこと書いた気がする。そしてそれを検索して確認する元気がない。元気重要。眠りたい。
20120607
ブラッドベリは「たんぽぽのお酒」と短編いくつかしか読んだことがなかった。西尾図書館で廃棄処分になっていた洋書をゲットして「華氏451度」「火星年代記」といくつかの短編集を積んでいたのだが、ちょうど先週捨ててしまったところだ。なんというタイミングだろう!
いまは、オーウェルの「カタロニア賛歌」を読んでいる。いまはバルセロナでバリケード作っているところ。飢えと退屈と寒さとがきつい、みたいにでさらっと書いてあるが、なにげに周りで人は死にまくっていて、死線を乗り越えてきているというのがじんわりと読み取れる。
20120615
Magic Trip (DVD) これのレビューで、"what really surprised me is how crew-cut all the Merry Pranksters were!" ここ笑った。1964年だもんな。
20120618
次男に「お父さんスイッチ」で操られた。なるたけキモめな反応を出すことに心を砕く。「あ」「開いた口がふさがらない。え?え”----------?」「い」「イカれてる、イっちゃってる、異ノーマル」「う」「歌いつづける、24時間。もぐってもぐってもぐってランランラン…」「え」「えずく。う、う、うぼぇーーーーー」「お」「おがくずだらけの海に飛び込む。どっぽぉーーーーーん」たいへん体力が必要。しかも6周する必要があるとか言われる。わけがわからないよ。
20120622
「高等遊民」になりたい。親から膨大な遺産を受け取って(<-そんなものはありません)、都会から1時間くらい離れた静かだがそれなりに便利な街で(昔の鎌倉のイメージ)、早朝に起きて書き物をして、昼から外に出て人間観察して、5時にはそば屋で日本酒を飲んで、そんなかんじで朽ち果てたい。
昨日はけっきょく3D and VR展に行くには時間がもう少なかったので、渋谷の街をちょっと歩いて、まっすぐ帰ってきた。ひさびさに歩く渋谷の街は何も変わっちゃあいないといえば変わっていない。いや、レコード屋が無くなった。べつに俺の体は渋谷を欲していないなと確認できたので収穫だった。
豊橋ではいつも通り精文館書店をぶらついて新刊チェック。いつも通りその手前に通りにはアフリカ系っぽい人が待機している。いつもいる。ファミマでは「まどかマギカ Half Age Characters」というのが新発売されていて、出来はよさそうだがラボの机以外置き場がない。あきらめた。
知らない街まで車で行ってみたい。昼はジャスコの駐車場で車中で寝て、夜になったらツタヤとかで涼んで、だらだらとサブカル本とか無駄に読んで時間を浪費して、夜はひたすら次の県へと車を飛ばして、10時にジャスコが開店したらまた駐車場で仮眠するんだ。ヴィレヴァンとか人工的なものに囲まれて。
美しい田んぼの風景とか美しい山並みとか河川敷とかには目もくれず、ファスト風土のまっただ中だけを縫いながら、日本を縦断してみたい。風景とか歴史名勝とかそういうものをまったく目的に入れずに、機械的に日本を縦断することだけを目的として行動してみたいんだ。
20120624
今日のフットベースの練習はキック練習担当だったので、投げまくって疲れた。お子さんらは5回に一回だが、こっちは全部投げる。腰に来た。
奥さんがソフトボールの練習の後にコクワガタを見つけて家に持ってきて、メープルシロップをしみこませたのに乗せておいたのだが元気がなかった。しょうがないのでどっかに掴まらせておいたようなのだが、消灯すると俄然元気になって天井をブーンブーンと飛び回っている。それを私は横になって見てる。
100円ショップでレエン・コオト(芥川風)を見つけたので買ってきた。イギリス人傘差さないっていうし、イギリスの雨は土砂降りじゃなくて小雨らしいし、ASSC16はこれで対処してみようと思う。折りたたみ傘は持って行くが持ち歩かない方針で。
0629
今日も次男を連れて空手教室へ。7時だけどまだ日が落ちていなくて、ちょうどいいかんじの夕焼けがビルの谷間に落ちていくところで、車を降りると次男がそっと手を繋ぎに来た。
長男や長女のときを思い出して、こんなことももうすぐになくなるのかと思うと感慨深い。この気持ちを忘れまいと思うのだけれども、じつのところいろんな記憶で上書きされてしまって誰のエピソードだったかごちゃごちゃになっているのだ。「納豆ご飯」のことを「なっとぐわん」と言ったのは誰だったか?
アノラックからパステルズバッヂを外して、引き出しにしまって、そして僕らは誓うんだ、この気持ちを忘れないと、だからさよなら」というわけだけど、この気持ちは忘れてしまう。いや、気持ちは覚えているけど、それと結びついていたエピソードが抜け落ちてしまう。
これってオートノエティックかつコンテントのないエピソード記憶?
次男が「2000年前の0年のときって太陽にものすごく近かったんでしょ?」って聞くんで違うって言ったら、「じゃあ無量大数くらい?」(無量大数は最近の次男のマイブーム)と聞くので、46億年前だよ、と答えたのだけれども、無量大数と比べたら46億年前なんてすぐ近くだな、と思った。
ネットで探して、原始的生命の始まりが39億年前、魚がいるのが5億年前、と説明しだしたら異様に興味を持った。年表にはさらに続きがあって、10億年後には太陽が膨大して地球が滅びるとある。パパたちはせいぜい100年しか生きられないから関係ないよと教えたのだけれども、また100年後になったらなにか変わっているかもしれないので教えてほしいと言われた。逆にこっちが教えてほしい。
2012年12月27日
■ 脳科学辞典「マイクロサッケード」の項目書いた
脳科学辞典の「マイクロサッケード」の項目を書いた。査読されるまえの原稿をブログ用の記事として活用してみる。
マイクロサッケード
英:microsaccade
同義語:マイクロサッカード、フリック(flick)
類語:固視微動、fixational eye movement
マイクロサッケードとは
われわれが視野の物体を見るために視点を固定する(固視)とき、われわれの眼球は完全に動きを止めているわけではない。固視微動またはfixational eye movementと呼ばれる三種類の眼球運動がある。一番目は高頻度(90Hz程度)で小振幅(<1 min of arc)のトレモア(tremor)、二番目は低速度(~6 min of arc / sec)のドリフト(drift)、三番目が高速度(~10 deg / sec)の跳躍的運動であるマイクロサッケード(microsaccade)(フリック(flick)とも呼ばれる)である[1]。本項目では三番目のマイクロサッケードについて記述する。
マイクロサッケードの性質
マイクロサッケードは振幅は1度以下、運動にかかる時間(duration)は25 ms程度、平均速度は10 deg / sec程度、頻度は1-3Hz程度であるが、個体差、種差などによって報告はばらついている[1]。
マイクロサッケードは運動としてはその名の通りサッケード(急速眼球運動)の振幅を小さくしたものであると言える。たとえば、サッケードでは運動にかかる時間(duration)と最高速度(peak velocity)との間に正の相関が見られ、主系列曲線(main sequence curve)として記述することが出来るが、マイクロサッカードもこの主系列曲線の上に乗る[2]。
マイクロサッケードと視覚
マイクロサッケードは単なる脳内ノイズの結果ではなくて、機能を持つと考えられている。マイクロサッカードは他の固視微動とともに網膜像を絶えず変化させることによって視覚入力を絶えず生成していると考えられている。つまり、静止網膜像の実験では、固視微動に同期させて視覚像を動かすことによって網膜像の変化をなくすと視知覚の消去(fading)が起こる(たとえば古典的な実験としてはYarbus[3]など)。
また、マイクロサッカードは意志によって止めることは出来ない非随意的運動ではあるが、ランダムな運動というわけではない。注意などによってその方向や頻度が影響を受ける[3]。たとえば周辺視野に視覚刺激を提示すると、その方向へのマイクロサッカードの頻度は提示直後(0.2秒程度)には上昇し、さらにそのあと(0.5秒程度)では頻度は平均よりも低下する[4]。
マイクロサッケードの脳内メカニズム
マイクロサッケードの指令は上丘吻側の固視ニューロン領域で生成されていることを示唆する報告としてHafed et. al. がある[5]。この論文で著者らはマカクザルが固視課題をしている間の上丘の活動を記録してその反応野をマップすることによって、上丘の眼球運動マップの吻側部にある低振幅部分のニューロンが活動することを明らかにした。
また、Martinez-Conde et.al[6]では、マカクザルが固視課題をしている間のV1の活動を記録して、視覚応答への影響を見たところ、マイクロサッケードの直後(50-100 ms)のV1の応答は増強される。これはマイクロサッケードが視知覚を増強している可能性を示唆している。
マイクロサッケードの計算論的モデル
マイクロサッケードの生成過程は上丘及びその下流の脳幹の神経ネットワークのモデルによって説明することが出来る。これまでに報告されている計算論的モデルとしては、Hafed[7]、Engbert[8]、Inagaki et.al.[9]などが挙げられる。
参考文献
-
Susana Martinez-Conde, Stephen L Macknik, David H Hubel
The role of fixational eye movements in visual perception.
Nat. Rev. Neurosci.: 2004, 5(3);229-40 [PubMed:14976522] [WorldCat.org] [DOI]
-
B L Zuber, L Stark, G Cook
Microsaccades and the velocity-amplitude relationship for saccadic eye movements.
Science: 1965, 150(3702);1459-60 [PubMed:5855207] [WorldCat.org]
-
Yarbus, A. L.
Eye Movements and Vision (Plenum, New York, 1967) - ↑
Ziad M Hafed, James J Clark
Microsaccades as an overt measure of covert attention shifts.
Vision Res.: 2002, 42(22);2533-45 [PubMed:12445847] [WorldCat.org]
-
Ziad M Hafed, Laurent Goffart, Richard J Krauzlis
A neural mechanism for microsaccade generation in the primate superior colliculus.
Science: 2009, 323(5916);940-3 [PubMed:19213919] [WorldCat.org] [DOI]
-
S Martinez-Conde, S L Macknik, D H Hubel
Microsaccadic eye movements and firing of single cells in the striate cortex of macaque monkeys.
Nat. Neurosci.: 2000, 3(3);251-8 [PubMed:10700257] [WorldCat.org] [DOI]
-
Ziad M Hafed
Mechanisms for generating and compensating for the smallest possible saccades.
Eur. J. Neurosci.: 2011, 33(11);2101-13 [PubMed:21645104] [WorldCat.org] [DOI]
-
Ralf Engbert
Computational modeling of collicular integration of perceptual responses and attention in microsaccades.
J. Neurosci.: 2012, 32(23);8035-9 [PubMed:22674278] [WorldCat.org] [DOI]
-
Keiichiro Inagaki, Yutaka Hirata, Shiro Usui
A model-based theory on the signal transformation for microsaccade generation.
Neural Netw: 2011, 24(9);990-7 [PubMed:21741208] [WorldCat.org] [DOI]
- / ツイートする
- / 投稿日: 2012年12月27日
- / カテゴリー: [Saliencyと眼球運動]
- / Edit(管理者用)
2012年12月24日
■ 脳科学辞典「サリエンシー」の項目書いた
脳科学辞典の「サリエンシー」の項目を書いた。著作権に関しては「各用語解説の著作権はそれぞれの執筆者に帰属します。執筆者は編集委員会へ無償で非独占的に使用する権利を与えています」となっている。そういうわけで、査読されるまえの原稿をブログ用の記事として活用してみる。
サリエンシー
英:saliency
類語・同義語:顕著性。サリエンス(salience)。
サリエンシーとは
もし夜空に月が光っていれば月にすぐに目が向くだろう。これは月が目立つ(salient)からだ。このように感覚刺激がボトムアップ性注意を誘引する特性を「サリエンシー」と呼ぶ。 夜の月がsalientであるのは周りの空と比べて明るいからであって、昼の月はsalientではない。つまり、サリエンシーは刺激の時間的または空間的配置によって決定づけられるものであって、その刺激自体の特性ではない。明るいスクリーンに暗い部分があればそこはsalientになる。つまり刺激強度が高いこと(たとえば輝度が高いこと)とサリエンシーが高いことは等価ではない。
しかし、一般名詞としてsalience / saliencyという言葉を(物理的な強度と対比させて)心理的な強度自体を表していることもあり、かならずしも上記の用法で統一されているとは言えない。
視覚探索
サリエンシーは、心理学的研究においては、視覚探索におけるpop-outという概念と関連している。
図1 視覚探索
図1の三つの刺激でそれぞれ仲間はずれの要素を見つけなさい、と問われたとしたら、左と真ん中の二つの図ではひとめで見つかるのに対して、右の図ではなかなか難しい。これは左の図では色特徴でpop-outするから、真ん中の図では傾きの特徴でpop-outするから、と表現される。
このような知見を元にしてAnn Triesmanは注意の「特徴統合理論」(feature integration theory)を作り上げた[1]。特徴統合理論では、視野像は各特徴(輝度、色、傾きなど)ごとに平行して処理され、それらの特徴が最終的に統合される。よって、図1左の色のpop-out刺激では、色特徴の処理の段階で仲間はずれを検出することが出来るので処理が速いのだが、図1右のconjunction searchでは統合された情報を探索しなければならないために処理が遅くなる、と説明される。
同様な現象は他のモダリティー、たとえば音刺激でも見られる。たとえば聴覚でのoddball taskでは、ピ、ピ、ピと連続する純音刺激に違う周波数の純音が混ざったり、または音が鳴らなかった場合にはその時に注意が誘引される。この場合は空間ではなくて、時間的配列がボトムアップ性注意を誘引する例と言える。
サリエンシー・マップ(saliency map)
特徴統合理論はあくまで心理学的な理論であったが、計算論的立場からどのようにボトムアップ性注意が計算されているかを説明するモデルとして「サリエンシー・マップ」が提唱された[2]。
サリエンシー・マップとは、特徴に依存しない視覚刺激のサリエンシーをスカラー量として計算して、二次元マップとして表現したもののことを指す。
サリエンシー・マップの機能的な特徴としては以下の二つがあげられる。
- 平行処理:特徴統合理論からの影響を受けているため、サリエンシーはまず各特徴ごとに計算されて、特徴マップを作る。
- Winner-take-allルール:これら複数の特徴マップが足しあわされて計算されたサリエンシー・マップの中からいちばんサリエンシーの高い部分が選択される。
Koch and Ulman 1985[2]においてはあくまで計算の原理のモデルであったのだが、それを実際の画像から計算できるようなモデルとして実現したのがItti, Koch and Neiburによるサリエンシー計算論モデルだった[3]。
このモデルのアルゴリズムレベルでの特色としては、
- 各特徴ごとのサリエンシーを計算するため、center-surround抑制を用いる。
- さまざまな解像度(pyramids)でこの作業を平行して行う(画像処理の分野でのmulti-scale representationに対応) 。
- 以上の操作を繰り返して正規化する(iterative normalization)。
がある 。
この計算論モデルはC++ソフトウェアとして、南カリフォルニア大学Ittiラボより、ソースがGNU General Public Licenseに基づいて入手できるようになっている。
このソフトウェアを使って図1の画像のサリエンシーを計算したのが図2となる。

図2 サリエンシー・マップ
ほかにもMatlabでのサリエンシー・マップを計算するプログラムとして以下のものがwebから入手可能である。
- SaliencyToolbox
- Graph-Based Visual SaliencyおよびItti, Koch, Nieburのサリエンシー・マップ
- Matlab codes for measuring image saliency
Itti, Koch and Neibur[3]以降、サリエンシー・マップはさまざまな実装が報告されており、たとえば三次元への拡張、トップダウン注意への拡張などcomputational visionにおいて重要な分野の一つとなっている。
サリエンシーの脳内表象
サリエンシー・マップはあくまで計算論的概念であるので、脳にサリエンシー・マップが表現されている保証はない。オリジナルの定義からすればサリエンシー・マップは単一のものであるはずだが、複数の処理レベルのサリエンシー・マップが脳内で分散して表現されていると主張しているものもある (たとえば[4])。
サリエンシーが表象されている部分としてこれまでに、V1[5]、上丘[6]、視床枕[7]、LIP[8]、FEF[9]、V4[10]などがその候補として挙げられている。
サリエンシー・マップの応用
画像や映像を見ているときの視覚探索をサリエンシー・マップによって予測するという一連の研究がある。そのなかではたとえば視覚探索時の眼球運動のデータからADHD患者やパーキンソン病患者を分類することに成功したもの[11]やマカクザルの視覚探索時の眼球運動のデータから第一次視覚野損傷の影響を解明したもの[12]などがある。
参考文献
-
A M Treisman, G Gelade
A feature-integration theory of attention.
Cogn Psychol: 1980, 12(1);97-136 [PubMed:7351125]/a> [WorldCat.org]
-
C Koch, S Ullman
Shifts in selective visual attention: towards the underlying neural circuitry.
Hum Neurobiol: 1985, 4(4);219-27 [PubMed:3836989] [WorldCat.org]
-
L. Itti, C. Koch, & E. Niebur
A Model of Saliency-Based Visual Attention for Rapid Scene Analysis.
IEEE Transactions on Pattern Analysis and Machine Intelligence: 1998, 20(11):1254-1259. - ↑
Vidhya Navalpakkam, Laurent Itti
Modeling the influence of task on attention.
Vision Res.: 2005, 45(2);205-31 [PubMed:15581921] [WorldCat.org] [DOI]
-
Zhaoping Li
A saliency map in primary visual cortex.
Trends Cogn. Sci. (Regul. Ed.): 2002, 6(1);9-16 [PubMed:11849610] [WorldCat.org]
-
Wolf Kienzle, Matthias O Franz, Bernhard Schölkopf, Felix A Wichmann
Center-surround patterns emerge as optimal predictors for human saccade targets.
J Vis: 2009, 9(5);7.1-15 [PubMed:19757885] [WorldCat.org] [DOI]
-
D L Robinson, S E Petersen
The pulvinar and visual salience.
Trends Neurosci.: 1992, 15(4);127-32 [PubMed:1374970] [WorldCat.org]
-
J P Gottlieb, M Kusunoki, M E Goldberg
The representation of visual salience in monkey parietal cortex.
Nature: 1998, 391(6666);481-4 [PubMed:9461214] [WorldCat.org] [DOI]
-
Kirk G Thompson, Narcisse P Bichot
A visual salience map in the primate frontal eye field.
Prog. Brain Res.: 2005, 147();251-62 [PubMed:15581711] [WorldCat.org] [DOI]
-
John H Reynolds, Robert Desimone
Interacting roles of attention and visual salience in V4.
Neuron: 2003, 37(5);853-63 [PubMed:12628175] [WorldCat.org]
-
Po-He Tseng, Ian G M Cameron, Giovanna Pari, James N Reynolds, Douglas P Munoz, Laurent Itti
High-throughput classification of clinical populations from natural viewing eye movements.
J. Neurol.: 2012; [PubMed:22926163] [WorldCat.org] [DOI]
-
Masatoshi Yoshida, Laurent Itti, David J Berg, Takuro Ikeda, Rikako Kato, Kana Takaura, Brian J White, Douglas P Munoz, Tadashi Isa
Residual attention guidance in blindsight monkeys watching complex natural scenes.
Curr. Biol.: 2012, 22(15);1429-34 [PubMed:22748317] [WorldCat.org] [DOI]
- / ツイートする
- / 投稿日: 2012年12月24日
- / カテゴリー: [Saliencyと眼球運動]
- / Edit(管理者用)
2012年12月20日
■ 計算論的モデルを使うということについて実感に基づきつつ。
2012/9/11
いままで自分が関わってきた計算論モデルは、サッカードの応答潜時をモデル化するdiffusion modelと、saliency mapという計算論的、心理学的な仮構物を中間に置いてボトムアップ注意をモデル化するsaliency model。
どちらとも、biologicalにplausibleであることに重点を置いて、たくさんのパラメータを入れ込んで行動データを予言するのに成功していたものを後から利用する立場だったんだけど、post-hocにパラメータどんどん突っ込んでその予言性能を上げるというのに違和感を持った。
当時はまだベイズ的なものを十分理解していたわけではなかったけれど。それで、どちらのときも不完全ながらモデル選択的なアプローチをした。
Diffusion modelの場合には解釈可能なパラメータ以外をなるたけ減らした上で、二つの条件であるパラメータthetaが等しいモデルと異なっているモデルとでAICを計算して、パラメータ増えたことを考慮してもthetaをべつべつにしたほうが良いモデルであるということを示した。
Saliency mapのほうはモデルを簡略化することは状況的に不可能だったから、チャネルの寄与をleave-one-outを使って評価した。こっちはcross validationはしていないし、いろいろ足りなかったが、やりたかったのはそういう方向性。どちらも、なんでもパラメータ突っ込めばいいというわけではないというのが基本的なアイデア。
そんなわけで、この方向性を突き詰めて、モデルベースで行こうとしたら、 たぶんベイズ的になるんだろうと思ってた。いっぽうで、この種のモデルはモデルの中でしか話ができない。たとえばsaliency mapの話で行けば、あれはもともとV1を経由した皮質での視覚情報処理のモデルなんだから、V1 lesionの話をこれで説明しようってのが端っからズレてる。
つまり、もっとぜんぜん違うモデルを持ってくれば予測性能が上がる可能性はまったく否定できない。Diffusion modelのほうも同じで、パラメータをthetaとmuの二つに絞っちゃったので、二つのグループの違いはこのパラメータの違いでしか説明できない。
もちろんモデルがまったく間違っているのならパラメーターの変動でデータを説明することがそもそもできないけど、もっといいモデルがあった可能性はある。以前も書いたけど、diffusion modelを含むaccumulator modelというのはevidenceを蓄積する。
でも、蓄積しないである時点でのevidenceの大きさが閾値越えるかどうかでサッカードするってモデルでも良い。ふつうはこういうモデルは有効でないけど、V1 lesionしたときにそういうものが有効である可能性はある。
まとめると、モデルベースだとモデルの空間の中で一番ましなものを探索しているだけということになる。まあ、そういってしまえば当たり前な気がするが、パラメータ空間だけ見てると、今言ったような蓄積モードかその時々モードかとかそういうものが見えなくなる。
そういうわけで、モデルベースでやることの限界と帰結みたいなものを自分なりの経験に基づいてまとめてみた。
こういうことに目が向く前は、統計でANOVAかけたとか、ここは対応があるデータだとか、多重比較がどうのとか、多重共線性がどうのとかそういうことに多大な興味があったが、モデルベースで考えるようになったらなんかどうにも間接的すぎる気がしたものだった。
神経生理の方と話していて、基本はt検ですよ、と主張するのを聞いてて、気持ちは分かる、たしかにそういう美しい実験デザインを汲んでみたいものだと思うけど、でも自分はずいぶんと隔たったところへ来てしまったなと実感した。
そんな自分としては、いまは信号検出理論とか見てるとあまりに原始的でもちょっとなんとかならんのかと思う。すくなくともガウス分布は勘弁してほしい。
そんなこんなで、あらためてこう書いてみると、自分がこれほどまでモデルベース思想だったかと驚いた。私はどっち向いて進んだらいいんだろう、と途方に暮れた。
ぜんぜん違うモデル、なんてあり得るかって議論はあり得る。たとえばサリエンシーモデルはもともとTriesmanのfeature integration theoryから来ているので、featureのチャネルはそれぞれ独立して計算されて、後で足しあわされる。当然、このようなパラレルな処理がいつも厳密に行われているとは限らない。でもそういうのは、featureののinteractionのチャネルを付加してやればよい(たとえばmotion * color)。けっきょくモデルを拡大して、パラメータを増やしてやることになるだろう。
同様にして、diffusion modelも蓄積モードと瞬時モードの両方を計算しておいて、必要ならどっちか荷重見かけてやればよい。こうして、モデルの柔軟性を上げることは、モデルの肥大化とそれに伴うパラメータ数の増大によって実現される。そうなると問題は「はたしてすべてのあり得る可能性を取り込んでいるのだろうか」というなんかフレーム問題みたいなものに帰着する気はする。そこまで行ってしまう前に、そもそもモデルの膨大かでは対処できないような致命的なモデルの改変というのはあるだろうか?
たとえばIttiのサリエンシーモデルは片道通行なのが特色で、Tsutosとかのものと比べると違っている。それすら、あらたに逆向きの流れを付加してやれば対処可能な気はする。とか書いているうちにだんだん分かってきて、これを推し進めるとけっきょくパーセプトロンからバックプロパゲーションに至ったニューラルネットワークと同じ道なのだな。なんという車輪の再発明w
2012/9/12
そういう意味であれば、数時間前に書いたdiffusion modelとsaliency mapを題材に書いた話とずれてはいないか。モデルを作っているときはそういうこと考えたりするけど、でも、論文を書いて戦っているときに「贋作だよな」という感覚をレフェリーと共有した経験はない。
いや違うか。「これこれこういうモデルの中で最適化したパラメータはこういう挙動を示します」みたいな言い方に留めておいて、もっと違ったモデルがあること自体は否定しないように書いているか。いまのところ、モデルをゼロから作ったわけではないのでそういう場面にはまだ遭遇してないんだろう。
「サリエンシーモデルはV1があること前提なのに、V1損傷のモデルをチャネルのゲイン変えるだけで対応っておかしいだろ」て言われたらどうしようかと事前に考えたけど、いままでに指摘されたことはない。そう言われたとしても「もっと良い(正しい?)モデルがある可能性は否定しません」でよいか。
「原理drivenで微分方程式で動かすようなモデル」このフレーズを見て、それからさっきの「お話としての説明」「物理学的な説明」 http://pooneil.sakura.ne.jp/archives/permalink/001364.php と突き合わせてみると、さっき書いたことは転倒していることが分かる。
つまり、認知科学的なモジュールを置いた説明が「お話としての説明」であって、バイオロジカルモデルが物理的因果で起こす現象が「物理的説明」で、「正しいモデル」というならそれは後者。前者こそが「贋作」であることを分かった上で使うべき仮構物。
Diffusion modelの話でいけば、サッカード開始のために「閾値を越える」というが、そんな閾値は仮構物。実際に起こっているのは、ニューラルネットワークの中でSelf-organized criticalityが出来ていて、どっかから雪崩が始まるということ。
以前diffusion modelをマーの三段階に当てはめようとしたことがあったけど、 http://pooneil.sakura.ne.jp/archives/permalink/001282.php あそこで書いたのと同じ。つまり、閾値はimplementationのレベルではなくて、もう一段抽象度が上がってる。
ニューロン間の相互作用(プロセス)があって、それのSOCとしての状態(なんか温度みたいなもの)をあらわそうとすると閾値(表象)になる。かくして表象とプロセスとのやりとりが抽象度をレベルを上下する。ここが「ベイトソン流に解釈する」って書いたところ。
だから、データドリブンにデコーダ作ってベイズの法則でひっくり返して(<-簡単に言いやがる)生成モデルを作るのも、認知心理学的な知見からモジュールを想定して作るモデルも両方とも贋作。すべてのニューロンの発火とシナプス伝達からなる力学系が「真作」。それは記述の抽象度を反映している。
ネットワークの状態(プロセス)を閾値として表象するとき、それは他者による観察ではあるけれども、客観的なものであるとは言える。
つまり、閾値は温度みたいな統計物理的な量に対応するから、ミクロでは定義できない。1個のニューロンの発火の閾値イコールネットワークのバーストの閾値ではないということ。ともあれ、物理量だから、まだここは物理的因果が働く世界であって、認知モジュール間の文法の世界ではない。
閾値はそういった統計量である一方で、蓄積されるエビデンスの大きさは発火頻度だから1個のニューロンの量のようにも見える。でもそうすると抽象度の違うものの間でのモデルになるからなんかおかしい。エビデンスは一つのニューロンでも集団でも質的には同じように見えるからだまされるのか。
2012年12月17日
■ クール・ビッズ!
クール・ビッズ!
うちの高校では柔道の授業があった。体育の先生が柔道の人だったのでみっちり指導してもらえてよかった。それでも怪我をした奴はいたし、私も水泳部の大事な試合の前だったから、乱取りで小林の体落としに無理な体勢になるまで粘らずに倒れたら、サメさんに「本気でやってないだろ」とスゲー怒られた。
浪人時代や大学生時代、なんかいいことあるんじゃないかみたいな根拠のない冒険心で、夜になると自転車でどっか遠くに向かうのだけれども、当然なにもあるわけがない。でもそんな気持ちだけで水戸街道を松戸の方まで延々漕いで、夜中の2時で周りは暗闇で、畑の肥料の香りがして、いったいなんだったんだろう、と思いつつまた家まで帰ったことがあった。そのときのひんやりした空気のことを憶えているような気がする。
井上陽水のセカンドアルバムのA面の最後の曲「夜のバス」、深町純のシンセとストリングスの編曲が印象的な曲だけど、よく聴いてた中学生の頃はこの「バス」をローカルバスだと思ってた。でもこれは長距離夜行バスだ。「どこにも停まらないで」「矢のように走る」わけだし。博多ー東京とかだろうか。
「ブラックボックスの内部では、政党や政治家、省庁、自治体、マスコミなど、あらゆる利害関係が複雑に絡み合い、限られた予算を巡って要求がせめぎ合っていた。しかも、それぞれがそれぞれの立場で正当性を持ち、必死に働きかけている。」(毎日JPリンク切れ) この感じはよく分かるし、これこそがcomplexityだよなあ。それをsimplifyせずにsimplicityを見抜くことなんて本当に可能なのかと途方に暮れる。
クール・ビッズ! (<-作務衣を着て硯に向かい墨をすりながら心を静めていくようなイメージで)
小林がかけた技は正確には、体落としで崩しておいて、そこでいったん私が持ちこたえようとしたのだけど、そこを膝車みたいなかんじで膝に来たので、これは危ないと思って飛んだのだった。なんでこんな26年前のことを覚えているかは分からないが、とにかく思い出したので、お詫びして訂正いたします。
ラノベを図書館から借りてきたので読みたいと思ったが、わざわざスタバに行くのもばからしいし、車でエンジン駆けっぱなしというのもよくないので、けっきょくちょっとは寒くなさそうな、ジャスコの4階駐車場で羅針盤とか聴きながら読んでた。
そうしたらいつのまにか閉店時間が過ぎていて、周りに車が一台もいなくなっていて、それでも読み続けたら、駐車場が完全消灯して、閉じ込められそうな恐怖感を感じて、あわてて逃げ出した。逃げ場所などどこにもなかった。
こんな強風と雨の中をカサぶっ壊しながら自転車でラボに行く意味なんてあるのかね(<-自虐的修辞疑問文)
説明するまでもないが「自虐的修辞疑問文」というのは、「XXXであろうか。いやそうではない。だがやるのだ。やれやれ」という意味の造語。
クール・ビッズ! (<-意味がないフレーズを繰り返していると、なんとなくアートっぽい雰囲気が出るものだという信念を胸に)
2012年12月13日
■ 盲視に関わるMPKチャネルの経路
2012/11/20
「V1から上丘へ行く経路はなにをしているか」これをアップデートした。まず、教科書的にはMPKはパラレルであるように考えられている。
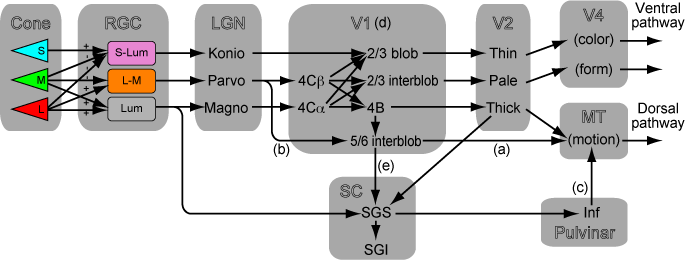
でも実際にはV1の中でかなり混ざっているってのが最近のCallawayの知見。Nat Rev Neurosci. 2009 "Parallel processing strategies of the primate visual system"
さらにV1をバイパスする経路を考えてみると、Koniocellularの系が活躍していることが分かる。
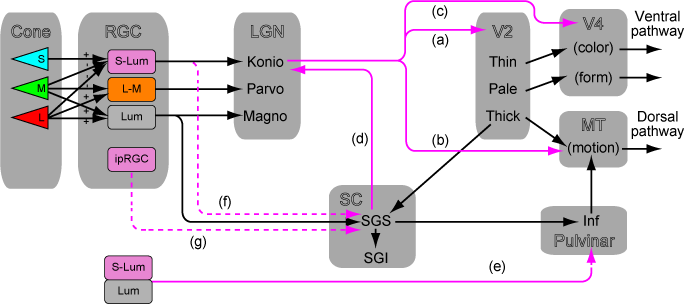
LesionしたマカクではW cell(=konio系)がマカクでSCに行っている。メラノプシン系はマウス。ラットでの知見。
ipRGCsが盲視に関わるかどうかの話をアップデートしておくと、Neuron 2010 "Melanopsin-Expressing Retinal Ganglion-Cell Photoreceptors: Cellular Diversity and Role in Pattern Vision" でipRGCsがpattern visionにも関わる(rot, cone欠失マウスでgrating vs uniformの弁別)ってのがある。
よく知られているSCNやOPNに行くやつはM1 ipRGCで(こっちはサーカディアンリズムとかに関わる)、それとはべつに、dLGNやSCへ投射するnonM1 ipRGCがある。TINS 2011 "Intrinsically photosensitive retinal ganglion cells: many subtypes, diverse functions"
Pattern visionに関わっているのであれば、ipRGCによる光への応答がどのくらい速いのかが重要だけど、PLOS Biology 2010 "Melanopsin Contributions to Irradiance Coding in the Thalamo-Cortical Visual System"を見る限りだと100msとかの潜時で出ているっぽいので充分速い。(斜め読み)
ただし、こちらはmRGCsって言ってるので同じクラスの細胞になるのかよく分からん。著者ごとに名前の付け方が違っているんだろうと思うんだけど。
Pupil reflexとかのproprioceptiveな感覚を盲視ではモニターしていて、ゆえに視覚ではなくて名状しがたい感覚になるのだ説(盲視の能力そのものではなくて「なにかあるかんじ」の説明)からすれば、ipRGC->pretectumがblink経由で情報与えてるかも。
V1から上丘への投射というのは、LGN(M)->4Calpha->4B->5/6interblob->SGSというルートがある。このため、TehovnikのようにV1 L6を微小電気刺激するとSCがドライブされてサッカードが起こる。
いっぽうでNassi et.al. Neuron 2006 "The Parvocellular LGN Provides a Robust Disynaptic Input to the Visual Motion Area MT"ではLGN (P)->V1 layer6 Meynert cells -> MTというルートでMTにもparvo入力が入る。 V1 layer6 Meynert cellsってのはSCにも投射しているかもしれない。
となると、LGN(M, P) -> V1 layer6 Meynert cells -> SC というルートでカラーの情報をV1経由でもらうことが出来るかもしれない。
昔のCoweyの論文 Neuroscience. 1995 "Nasal and temporal retinal ganglion cells projecting to the midbrain: implications for "blindsight"."を読むと、V1 lesion後に生き残ったRGCの細胞はW cell (koniocellular)であると書いてある。つまり、マカクでもretinotectalでS錐体の情報は入る。
ではなんでBrian WhiteのJNS2009 "Color-related signals in the primate superior colliculus"で上丘で速い色応答が出なかったのか。それはSGSから記録してないからじゃん?ってことになる。CatではM cellはSOへ、K cellはSGSへ入力する。Acta Neurobiol Exp 2004 "Motion sensitivity in cat's superior colliculus: contribution of different visual processing channels to response properties of collicular neurons"
なるたけコンタクト小さい電極で浅いところから記録して、DKLで刺激を作って、saturationをmaxにしてやる。つまり、backを灰色ではなくて、blueにしたうえでyellowへの応答を取る。このぐらいやれば応答が出てくるんではないだろうか?
「V1 layer6 Meynert cellsってのはSCにも投射しているかもしれない」というのは当てずっぽうではなくて、Meynert cellsだって言ってる人がいる。
上丘浅層にrabies virus入れて、V1経由で二次で染まるLGNのニューロンを調べてみればいいと思う。Konio, parvo両方が染まるはず。
Lyon 2010 "A disynaptic relay from superior colliculus to dorsal stream visual cortex in macaque monkey"でMTにrabies virus入れてdysynatpicにSCが染まる。これはinferior Pulvを経由している。2011年のIBROのときにCallawayがトークをしてたのでいろいろ質問した。うろ覚えだけど、SC->LGN->MTの可能性を聞いたら、染まっているSCの層はLGNに投射するlayerではないからunlikelyだという話を聞いた。たしかにあらためて Lyon et al 2010を読むと、そういう議論をしている(p.275左上)。SCのlayer2a->LGN(K)とlayer2b->infPVとなってる。
でもって、MTに入れて染まるのはSC layer 2b。 これでSC->PV->MT説にけっこう自信を持った。Leopordの言うようなSC->LGN->MT説は解剖学的にはunlikely。
それでもやっぱりパルボ系をSCに持ってくるのは難しい。しかし、実際にはLGNのレベルでも生理学的にはMPKは混ざっている。Logothetis et al Science. 1990 "Perceptual deficits and the activity of the color-opponent and broad-band pathways at isoluminance"ではこう書かれている:
These results suggest that impairment of visual capacities at isoluminance cannot be uniquely attributed to either of these systems and that isoluminant stimuli are inappropriate for the psychophysical isolation of these pathways.
つまり、isoluminantにしても、LGN parvoのニューロンの応答はゼロにはならない。
2012/11/20
マーモセットではretina->inf Pulv->MTがLGN->MTやV1->MTよりも早く形成される。JNS2012 "The Early Maturation of Visual Cortical Area MT is Dependent on Input from the Retinorecipient Medial Portion of the Inferior Pulvinar" 呼び水説として尤もらしい。この著者は以前にもadultでこの経路を報告している。Front. Neuroanat. 2010 "Retinal afferents synapse with relay cells targeting the middle temporal area in the pulvinar and lateral geniculate nuclei"
Eur J Neurosci. 2007 Selectivity of human retinotopic visual cortex to S-cone-opponent, L/M-cone-opponent and achromatic stimulation Human fMRIでDKL(lum, R/G, B/Y)のそれぞれの刺激で視覚野がどのようにactivation起こるか。MTの活動パターンがとても特異。B/Yでboldが下がってる。
fMRIでのBOLDがベースラインから下がるのはけっして抑制とは言えないわけだが、とにかくほかの領野とはものすごく違っていることが分かるし、やっぱLGN->MTのkonioがfunctionalに効いてんだろう、とすごく印象深い。
2012/12/4
高等研カンファレンス二日目より参加。ポスター発表終了。狙い通りDavid Leopoldと議論をすることが出来たのでミッションコンプリートした。David LeopoldはRGCからLGNとSC両方へ投射しているニューロンの重要性を強調していた。これはJNS 2008 "Y-cell receptive field and collicular projection of parasol ganglion cells in macaque monkey retina"およびJNS 2008 "The smooth monostratified ganglion cell: evidence for spatial diversity in the Y-cell pathway to the lateral geniculate nucleus and superior colliculus in the macaque monkey"のこと。
これはmagnoであって、parvoではない。retina->SCにkonioがあるとする私の持論についてはけっこうふつうに肯定していたので驚いた。ただ、上記のJNS2008二つを見たら、"lacked measurable S-cone input"って書いてあった。うーむ、いまだにS-coneがSCに入っているという証拠は見つからず。
2012/12/5
まさに俺得論文。Cerebral Cortex 2012 "Bypassing Primary Sensory Cortices A Direct Thalamocortical Pathway for Transmitting Salient Sensory information"
- / ツイートする
- / 投稿日: 2012年12月13日
- / カテゴリー: [上丘、FEFと眼球運動]
- / Edit(管理者用)
2012年12月07日
■ フリストンの自由エネルギー原理関連ツイートまとめ
(2012/6/22)昼ご飯でTom Froseとけっこう突っ込んだ話が出来たので(フリストンの自由エネルギー原理とヴァレラのコヒーレンス理論とか)これは有意義でした。
わたしの折衷主義的なアイデア(dorsalはenactiveでventralはpredictive coding)はTom Froseを満足させることは出来なかったけど、「自由エネルギー原理が平衡状態を前提としたものである点が限界」ということで意見が一致しました。
(2012/6/23)Tom Froseにも同じこと言われた。川人先生の双方向理論と同様、感覚と運動両方に適応できるところがインパクトなのはその通り。Predictive codingは腹側に限る、という方が正確です。背側腹側かは不明だけど、理論適応範囲を定めたいという動機です。
サッカードによる視覚探索の場面だと、自由エネルギー最小化はinfomaxと等価になって、これはNature 2005 とかで言ってることと同じになる(たぶん)。
Marrの3段階説で、物理的実装の層, アルゴリズムの層, 計算論の層ってあるときの計算論ってところがなにか最適化しようとするものを定める(ジャーク最小化とか)。自由エネルギー最小化ってのは計算論の層を抽象化してまとめようとしている。どちらも工学的問題解決だから話が収束する。
そうすると、いろんな計算論的問題があるんだけど、それの第4層に自由エネルギー最小化ってのが来て、すべてが根本的には同じ問題である、というなんかピラミッドみたいな描画になる。ここまでイメージを広げると、これはE=MC^2のような統一理論ではなくて、雑多な理論に共通性を見いだすメタ理論の提唱と考えた方がよいことが分かる。
(これはFrisotn自身も言っていた。何か新しいことを見つけたというのではなくて、それらを繋ぐものだから、自由エネルギー"principle"と呼んでいるのだと。)
視覚の方は腹側経路をメインにした論理構築で、運動の方はリーチングを元にしているというのがちぐはぐなのであって、vision-saccadeみたいな系で入口から出口までひとまとめで説明するというのが「統一的説明」が取るべき戦略ではないかという気がしてきた。
(2012/6/27)Perceptions as hypotheses: saccades as experiments まさに以前書いたフリストンでサッカードでベイジアン・サプライズだった。これ読んで駒場の冬学期の大学院講義までにこのへんネタにできるようにしておく。
(2012/6/28)(神谷さんによるFree-energy minimization and the dark-room problemの紹介に対して) ご紹介ありがとうございます。さらに論文出てるのですね。もうぜったい追いつけない。「暗い部屋問題」は行動選択の際にsurpriseを下げる=unlikelyな状態を避ける=餓えるまで動かないとかを避ける、で解決すると理解してたのだけどやっぱ問題なんですね。
(2012/7/25)Predictive coding云々と言い過ぎたが、神経生理学者としてどのくらい尤もらしいか言うなら、"explain away"するために「引き算」するって考えは、ぜんぜんニューロンの抑制ってものを分かってないと思う。ニューロンの抑制というのは線形的に足しあわされない。
Shunting inhibitionみたいなかんじで、コンダクタンスに効いてくる。だから、ニューロンの抑制は非線形的に、割り算のように効いてくる。これがHeegerのnormalizationモデルとかで使われている機構。
わたしがよく使う話で「ニューロンの応答はonsetで最大で、その後弱まる。だから、ニューロンの応答はfeatureそのものをrepresentしているのではなくて、そのsurpriseなのだ」みたいな説明をするけれども、じゃあなんでニューロンの応答はゼロにはならないかというと、それはexplain awayされてゼロになるのではなくて、割り算されて小さくなるから、というのが妥当なところだろう。
ちなみにIttiのbayesian surpriseでの説明では、temporal surpriseは時間が経てばゼロになるが、spatial surpriseはゼロにならないので、足し合わせるとゼロにはならない、という説明になる。あと、Fristonの図に出てくるlayer2/3がprediction errorでlayer5がpreditionというのは解剖学からの類推であって、生理学的には根拠がない。これはempiricalな問いなので、二光子の人にはぜひ検証してほしい。
とにかくempiricalな証拠が足りなくて、これまで出てきたようなRao and Balladのend stoppingとか、Hosoya et.al.のRGCとかはmodulationとかadaptationのレベルなので、これがfeature検出器説をrejectするわけではない。そういうわけで、このラインで寄与できることにこそ進むべき。
要は、生物学的なplausibilityだけ考えていると、計算論的原理がさっぱり見えてこないので、アラン・チューリングがやったみたいにsimplicityを見つけ出すべき。だから情報理論的に考えたいって話になる。
(2012/8/7)Attention Reverses the Effect of Prediction in Silencing Sensory Signals ハクワンとFloris de Langeとで、フリストンのラインで注意について議論してる。
Floris de LangeってのはDehaeneのところにいた人だった。
- / ツイートする
- / 投稿日: 2012年12月07日
- / カテゴリー: [ニューロンの情報コーディング]
- / Edit(管理者用)
2012年12月03日
■ 甲状腺刺激ホルモン放出ホルモン(TRH)のスプレー式点鼻薬
甲状腺刺激ホルモン放出ホルモン(TRH)のスプレー式点鼻薬 こんなことしてだいじょうぶなのか?TRH->TSH->T3,T4->TRH,TSHへネガティブフィードバック、となっているんだから、点鼻しないとTRH出ない事態になりそうだけど。
元ネタ( ArmyTimesおよびIndiana University)読んだが、点鼻スプレーを開発するというプロポーザルにお金が下りた、ということか。まあ、オキシトシン点鼻薬とかと同じ発想ということかな。
TRHを脳内投与するとratではarousalが起こる。これは甲状腺への作用ではなくて、オレキシン産生ニューロンを刺激することによるらしい。( JNS2009および JP2009) (ほかの系へもいろいろと作用することが知られている。)
すると、バセドウ病で自己抗体がTSHの代わりに甲状腺を刺激してT3,T4を産生、これが発熱や頻脈などを起こす。一方でT3,T4のネガティブフィードバックによってTRHは下がる、これがオレキシン系経由でarousalを下げてちぐはぐな状態になる、というところなのか。
ああ、もっと基本的な知見として、TRH ->NAc -> DA放出 -> locomotor activityという経路があるのか。 Neuropharmacology. 1979 および Eur J Pharmacol. 1981
お勧めエントリ
- 細胞外電極はなにを見ているか(1) 20080727 (2) リニューアル版 20081107
- 総説 長期記憶の脳内メカニズム 20100909
- 駒場講義2013 「意識の科学的研究 - 盲視を起点に」20130626
- 駒場講義2012レジメ 意識と注意の脳内メカニズム(1) 注意 20121010 (2) 意識 20121011
- 視覚、注意、言語で3*2の背側、腹側経路説 20140119
- 脳科学辞典の項目書いた 「盲視」 20130407
- 脳科学辞典の項目書いた 「気づき」 20130228
- 脳科学辞典の項目書いた 「サリエンシー」 20121224
- 脳科学辞典の項目書いた 「マイクロサッケード」 20121227
- 盲視でおこる「なにかあるかんじ」 20110126
- DKL色空間についてまとめ 20090113
- 科学基礎論学会 秋の研究例会 ワークショップ「意識の神経科学と神経現象学」レジメ 20131102
- ギャラガー&ザハヴィ『現象学的な心』合評会レジメ 20130628
- Marrのrepresentationとprocessをベイトソン流に解釈する (1) 20100317 (2) 20100317
- 半側空間無視と同名半盲とは区別できるか?(1) 20080220 (2) 半側空間無視の原因部位は? 20080221
- MarrのVisionの最初と最後だけを読む 20071213