[月別過去ログ] 2008年11月
« 2008年10月 | 最新のページに戻る | 2008年12月 »2008年11月28日
■ 子どもと「BigDogごっこ」をした。
パパがyoutubeで見ていたBigDogのことを、一番下の子がえらく気に入って、「へんなドボット見たい」とせがむようになった。ついでにBigDog Betaも見せてやったら大喜び。
毎日見せるのもよくないかと思って、今日は代わりに「BigDogごっこ」をやってみた。つまり、ふたりで頭と頭をくっつけて、「ブーブー」とうなりながら行進する。そこを横から長女が蹴りを入れて、おっとっととなりながらもなんとか耐える。そんなあそび。ウケてた。
そんな日のこと。
2008年11月26日
■ 「補正」が必要なのは、モデル化が不充分である証拠
超背伸びして書きました。怪しいところをwebで確認したりせずに書いた。もうしらない。厳しくせずに、褒めて伸ばしてほしい。
で、情報理論ってなんか嫌いなんですよね。っていうかニューロンの発火の解析関連での情報理論の応用ってのが嫌いってのが正しいのか。
なにがいやって、扱っているのがp(x,y),p(x)とか確率で、その計算をするのに使ったnが出てこない。基本的に無限試行行った後の理想的状態とかしか考えてないわけですよ。それは統計物理のようにものすごい多い数を扱っているときは良い。ていうかエントロピーって発想自体が元々そこからですからね。だけど、ニューロンの記録のように試行数10回とかでやっている事象にそのまま当てはめるわけにはいかない。
とは言いましたがもちろんわたしもITニューロンの発火パターンの解析に活用していようとか思っていろいろ勉強していたことがあったし、いくつか解析もしてました。そのころはちょうど
"Spikes: Exploring the Neural Code"が出た頃だったし、Panzeriの一連の仕事、たとえば
Golomb D, Hertz J, Panzeri S, Treves A, Richmond B. "How well can we estimate the information carried in neuronal responses from limited samples?" Neural Comput. 1997 Apr 1;9(3):649-65.
(ちなみにPanzeriはさいきんもStefano Panzeri, Riccardo Senatore, Marcelo A. Montemurro and Rasmus S. Petersen "Correcting for the Sampling Bias Problem in Spike Train Information Measures" J Neurophysiol 98: 1064-1072, 2007ってレビューを出してるのを知った。)
とかを読んだり、菅生さんのNature
Sugase Y, Yamane S, Ueno S, Kawano K. "Global and fine information coded by single neurons in the temporal visual cortex." Nature. 1999 Aug 26;400(6747):869-73.
が出た時代で、これのインパクトは大きかった。
んで、けっきょくのところ、少ない試行数だと、試行間のvariationの分だけ情報をoverestimateしてしまいます。(音A刺激の試行と音B刺激の試行とで視覚野ニューロンのスパイク数を数えれば、試行間のばらつきがあるから、相互情報量>0となってしまう。) だからその分の補正をしようってのが上述のGolomb et.alとか含めていくつか仕事があるわけです。
そのころは相互情報量ってけっこう扱いにくいなあとかそのくらいに思ってたんだけだけど、その後の業界的にも「選択性の指標」みたいなかんじで相互情報量を使うってのはあまり見なくなってきたし、私もすっかり忘れてました。(いまだにノンパラでの分離度としてROCのd'とかAUCとか使うのは多いんだけど、あれはなんなんだろ。自分でも使ってるけどね。)
んでずっと放置してたんだけど、この「補正」という発想がポイント(ガン、って書こうと思ったけど、これっていまどきpolitically incorrectですかね)なんではないかと思ったんです。つまり、「補正」が必要なのは、モデル化が不充分である証拠。
一つの例はあれですね、fMRIでのvoxelごとに検定をすることによるmultiple comparisonの問題を、Bonferroniの「補正」を使うのではぜんぜんpracticalでない(補正が効き過ぎる)のを、Worsley-Fristonがrandom field theoryという、voxelのデータにsmoothingがかかったものを明示的にモデル化した理論を使うことによって解決したわけです。これによってfMRIの解析のスタンダードが確立したと言えると思います。
もうひとつ例を挙げると、時系列データの解析(たとえばLTPの時間経過の検定とか。それともいまだに時点ごとのt検やってんのかな)では、repeated measures ANOVAがよく使われますが、球面性の仮定が成り立たない場合、自由度に「補正」をかけます(Greenhouse-GeisserのεまたはHuynh-Feldtのε)。つまりじっさいのデータの数から計算した自由度では自由度をoverestimateしてしまうから、自由度を下げてやっているわけです。
こちらのほうの問題はけっきょく、球面性の仮定を行っていないMANOVAを使うことで解消されました。つまり、モデル内で要因間の差の分散が違っていることをを想定していないrepeated measures ANOVAで、要因間の差の分散が違っている分の補正をかける、なんてトリッキーなことをせずに、残差の共分散構造をあらかじめモデルに取り込んだMANOVAを使うほうがstraightforwardなわけですから。(じっさいにはMANOVAはデータ数が少ないと使えないとかいった弱点がありますが:反復測定 ANOVA か、(G)MANOVA かの選択の問題 ) General linear modelからgeneralized linear modelへの拡張によって問題が解決した、というのが正しい言い方でしょうか。
んで、翻って、おなじような解決法が「ニューロンの発火の解析関連での情報理論の応用」でも見られないかなと思うんです。ここはたぶんベイジアンですよね。というのも、「少ない試行数だと、試行間のvariationの分だけ情報をoverestimateしてしまう」というのは、fittingにおけるoverfittingの問題とたぶん等価もしくは相似ですよね。(PRML本の第一章読んだのでかぶれてる…) Overfittingの場合も、データのnが少ないときに推定値の分散を考慮していないためにバイアスが出る、というのが元凶でした。
ペナルティの項を与えるっていう発想はちょっと「補正」の発想に似ていていやな気はするけど、試行数を明示的に入れた上で相互情報量やKL-divergenceのことを考える、というとベイジアン的な取り扱いをするということになりますよね。そういうのってあるんだろうか。たぶんあるんでしょう。よく知らないけど。なかったら作るべきだ。相互情報量の「補正」なんかしてないで、このレベルから捉え直すべきだ。たぶんbinの問題(binの存在を前提としていること、binの中に必ずデータが入っているようにbinを切らないといけない)もここで解消すべき問題なんではないかと思います。
そもそもわたしの分野で、真の意味で情報理論的取り扱いをする必要がある部分はどこにあるのか(反応選択性の指標代わりとかぢゃなくて)、というあたりが問題だったりもします。ただ、これからBMIとか大規模データ集積とかそういうところが発展していくことによって、たぶん必要なデータ解析自体は変わってくることになります。もともとこのfieldは、subjectのn=2で、記録ニューロンの数がm=100とかそういった歪んだ状況でANOVAとかGLMとかやってきたわけでそれはおかしな話でした。次へ進むためには、そのへんは整備されないといけない。これがわたしの仕事だとは思わないのだけど、上記のFristonがfMRIで行った業績のように、だれかが手がける必要がある問題なんじゃないかと思ってます。
落ち穂拾い:nをものすごく大きくするとなんでも有意になってしまうという問題があります。けっきょくこれはp-value至上主義の弊害であって、ホントはtype I errorとtype II errorとでのpayoffを考えるべきなんですよね。
# しか
僕は,情報量や情報量規準を使って神経科学について何か言うにしても,いまのところは間接的か限定的だと思います.
ところで,情報量規準で,一致性(標本数が無限の時に真のモデルと一致する性質)の「無い」AIC(Akaike Infometion Criteria),それを考えた赤池さんはデータが有限であるってことを理論体系の前提にして考えたとか(←受け売りなんで,その実をわかっていませんが).
僕なんかは,そのモデルでデータを説明(or 予測 or きれいにフィッティング)できたら,真実を表現していると思えちゃうんですけど,一致性が無いから厳密に言ったらダメなんですよね??.......この辺,科学者はどう考えてるのだろうと思うところです.
PS.
間違い発見.
「菅瀬さん」→「管生さん」です.
今回の問題提起は、つまるところ「fMRIにおけるFristonの仕事のような『統計の使い方の統一』がnhpにはないが、どうしたら良いか」というお話になるのでしょうか?
似たような統計解析の議論はhuman EEGでもよく耳にしますね。巷で有名な論文でも、100本読めば100通りの統計を使っていてどれを信用したらいいんだかいまいちわかりません。
また、Fristonのrandom field theoryにしてもindividual activation mapが重要になるテーマでない限りは、結局nを増やしてrandom effectsというお定まりの展開に持ち込む研究者が多数派ですので、実は現場的にはあまり意味がない(しかも最近はFDRのようなもっと「ヌルい」補正もあるので)ような印象もあります。
# pooneil>>データが有限であるってことを理論体系の前提にして考えたとか
確かなことはわかりませんが、ベイジアン的発想の方がデータが有限であることに対応しやすそうだ、というheuristicsをもって勉強しているところです。
あるモデルでデータを説明できたときに、より包括性の高いモデルを考えてゆくというのがある道かと。その意味で、モデルの前提部分から作り直す必要があるかどうかの目安の一つが、現状のモデルがpost-hocな「補正」を必要としているかどうかではないか、というのが今回のエントリの論旨です。
あと、訂正ありがとうございます。
>>fMRIにおけるFristonの仕事のような『統計の使い方の統一』がnhpにはないが、どうしたら良いか
これに関しては以前のこれ: http://www.mumumu.org/~viking/blog-wp/?p=175 で書いたことと繋がるかと思いますが、あまり明確なことを考えていたわけではありません。あくまで、「「補正」が必要なのは、モデル化が不充分である証拠 」というのがいくつかの場面で当てはまるのではないか、だからたぶん相互情報量を使った解析でも同じようなブレークスルーが必要なのではないか、ということを提案したわけです。
Random field theoryの話はぶっちゃけ受け売りなのですが、たしかこれもけっきょくのところ自由度の補正というところに行き着くはずで(記憶が不確か)、もっと先の話があるのでしょう。フォローできてませんが。それでも、たぶんFriston以前から考えれば大ブレークスルーだったはずでして。
解析の手法の進歩が漸次的に起きていくときのダイナミクスみたいなものを抽出してやろうという試みとして読んでいただけたらと思います。
>> 結局nを増やしてrandom effects
このへんのことも本当は考えたかったのですが、息切れしてしまいました。教科書的にはbeta (検出力)の評価とか、effect sizeの評価とかを考えることになると思いますが、それだけだと足りないよなあというのが実感です。
そういえば、Friston vs. Kanwisherの時に似たようなお話しましたっけ。
Fristonのrandom field theory(もしくはfamily-wise error: FWE)というのは、確かに根本義から言えば自由度の補正ですね。つまり、賦活voxelの数に応じて自由度を補正する(Bonferroni)のではなく、賦活clusterの数に応じて補正する(random field theory)というアイデアです。
ただ、もちろんFWEにも欠点はありまして、一番わかりやすいのがspatial filteringへの制約です。当然ですが、FWEだとclusterをどう決めるかに検出力が完全に依存しますので、例えばspatial filteringの際のGaussianのFWHMをvoxel sizeに対してどれくらいの倍率にしたかによってp-value thresholdがアホみたいに変動します。そうなると、「FWEで高い検出力を得るためにfilteringする」のか「賦活をcluster単位でわかりやすくまとめるためにfilteringする」のかわからなくなります(本義的には後者であるべき)。
fMRIの方はnhpに比べてnを大きく取れますので、この辺の問題を嫌がって「小さいnでfixed effectsでFWEをかけたmapを出す」よりは「大きいnでrandom effectsでuncorrected p-valueのmapを出す」という研究者が多くなるのは自然なことでしょう。おそらく、fMRIの初期にはnを増やすのが難しくて(同意してくれるボランティアがラボの身内ぐらいしかいなかった時代)こういう苦労をしてきたのが、今は脳に興味を持つ人が増えてwebで公募してもたくさん集まってくれるようになって要らなくなったという部分もあるのではないかと。
・・・というfMRIにおける統計学の移り変わりを見るに、nhpの発火パターンを扱う統計学についてもいずれ同じような展開が出てくるのではないかという気がします。ただ、その前に大事なのはどの情報量を用いるのか、どの理論を用いるのかという以前に、「何が独立で何が独立でないデータなのか」をはっきりさせることなのかもしれません(nhpは全くの素人なので的外れかもしれません、ごめんなさい)。近接しているニューロンでも実は独立とか、離れたところにあるニューロン同士でも実は独立でないとか、そういうところにも解決のカギがあるように思えます。
2008年11月21日
■ Twitterに脳内がだだ漏れる20080918
Twitterに書いたことを断章形式でブログに再利用。前回は20080809まで。
8/9 質より量に学ぶ - Radium Software//まったくもってホント重要。//「泥のように働く」で荒れたけど、こういうことを指してたんじゃないのかな。
8/11 北島58秒台キター!!!
8/12 Proteus VXフリー化!!! スゲー!!!//さいきん、スゲー!!!とかキター!!!とかそんなのばっかり。//Proteus VX落としてきた!//スタンダロンで動かしてみた。音鳴らねえYO!!!//FL Studio 8からVSTで読み込んでみた。 音鳴らねえYO!!!
8/13 twitterfoxからテスト。//こ、こりゃええ(タモリっぽく)
8/25 "Twitter is over capacity"出た! もう引っ越すか。
8/27 つらいときには明るい歌を歌い、平和なときにブルースに絡まる。//言葉には二面性があり、僕らの主観性には二面性がない。//だから僕は、主観性は表象に先立つ、って言ってるんだ。//だからこんな日は、ビール飲みながら「バナナフィッシュにうってつけの日」を読む。//ビール、にが。
8/31 みえるものすべてが、灰色になって、//遠くにあるように見えて、//それはそれでかまわないのだけれど、//ちりちりとしたかんじが残るのだけが、//どうにもゆるせないのだ//そうぞうの世界で。//ぼくはひとりで立っていた。//とおくで音楽がながれていた。//トレモロをかけた、ひどく歪んだギターの音が、//遠ざかったり近づいたりするように、//トーンをびみょうに上げ下げしてゆく//やはりぼくはひとりで、//なつかしいことを思い出したりとかしながら、//ひっしに耐えていた。
9/1 ここはポエムを垂れ流すところにしたので、//フォロー全部切ったのであしからず。//「入滅」なんてかんじで。
9/6 今日みたいに寝苦しい夜は//カエルがゲコゲコ鳴いて//水に飛び込む音がしたりして//...家の中で。//って気づいたらスゲーでかくなってるし。//もう寝る!
9/17 Paradieswarts。xx-boundてかんじか。//ノースバウンド、アースバウンド。//だから、訳すなら「楽園行」で。//楽園へ、いこう。//いまの俺がなりたいのはただひとつ。//13th floor elevatorsの「壺男」。//あ、研究会の感想だけれど、私がいちばん印象深かったのは、//iPhone持ってる人にまったく遭遇しなかったってことです。//あ、冗談ですよ?(>>いちばん印象深かった)
9/18 「XXですよ?」が使いたかっただけなんだけど、いまいち。
2008年11月13日
■ 飛行機で読む本の候補200811
いよいよSFNですね。しかし今年はわたしは不参加です。ネタはあったんですが(JNS論文のdiffusion modelの部分とか、NCM2007で出したV1 lesionでのexpress saccadeとニューロン活動とか)、今年はちょっとあちこち行きすぎたので、実験に専念ということで。
……のはずだったのですが、HFSPミーティングでカナダのDoug Munozのところに行くついでにニューヨークのHakwan Lauのところに行って、コロンビア大学の心理学部のセミナーでトークをすることに。関係する方を訪ねたかったのですが、あいにくSFNの会期中なのでそれもかなわず。またの機会に!
んで、帰りのトロントから成田までの飛行時間が17時間ですよ。給油時間込みとはいえ。
仕方ないので読む本を探しました。さいきんはラノベばっかり読んでましたが、さっさと読み終わってしまって時間が保たないので、なんか長編を探してみる。
「鉄鼠の檻」 (文庫版) 京極 夏彦はいつも候補に入ってるんだけど後回しになりそう。
「ビューティフル・マインド」 シルヴィア ナサーも積んだままだったので読もうかと思ってアマゾンの批評を見たら(見ちゃうし!)、訳がひどいと書いてあったのでがぜん萎える。
「オウエンのために祈りを」 ジョン アーヴィング。アーヴィングは大学の頃に「ガープの世界」「熊を放つ」を読んだ以来で当分遠ざかっていたのだけれど、最後が一挙に来るみたいでスゲーよさそうだし、新潮文庫に入ってたので買いました。
「燃えよ剣」 司馬 遼太郎。自分はシバリョーとか、っていうか歴史物をまったく読んだことがないので、死ぬ前にここらでチャレンジ。楽しめるかどうかではいちばん間違いないはず。ちなみに岡崎には近藤勇の首塚があって(法蔵寺)、縁のある土地なのですが、まだ行ったことありません。
PRML訳本読むってのも急務なのだけれど、飛行機では無理だな。あー、ベイジアン、ベイジアン。
「全米NO1「こころの主治医」が教える なりたいカラダになれる7つのプログラム」 フィリップ・マグロー さすがにそろそろダイエットしないとまずいので。経験上(ダイエット成功歴2回! でも失敗歴も2回)、いかにしてわたしが「ダイエットモード」と呼んでいるマインドセットに入るかがダイエットには重要なのですよ。そこに入れば痩せるのはつらくない。逆に言えば、ダイエットがつらいような状態のときは成功しないんです。このあたりの体験談は昔書きました:20000816。ということでこの本が良さそうなので、図書館から借りてきました。次回のダイエットのテーマは「健康に痩せる」でいきたい。
というわけで「 オウエンのために祈りを」と「燃えよ剣」をかばんに入れてくことにしました。
2008年11月07日
■ 細胞外電極はなにを見ているか リニューアル版
京都で開催された脳プロ分科会の「皮質脳波」のセクションで「細胞外電極はなにを見ているか」ということで10分トークをしてきました。
以前のブログのエントリ(「細胞外電極はなにを見ているか」)を元ネタにして作成していたのですが、あれは電気生理はじめての方も含めたトレーニングコースでのチュートリアルでしたが、今回は神経生理、工学系、臨床系の先生方の前ですので、かなり変更を加えることになって、けっきょくのところほとんど新しいものを作ることになりました。
そうしたら準備した内容が10分ではとてもしゃべりきれないものになりましたので、当日はいちばん関連する部分だけしゃべることにして、増補したものをハンドアウトで配布しました。さらに増補したものをLaTeXを使ってPDFファイルとして文書を作成しましたので公開します。ダウンロードはこちらから:「細胞外電極はなにを見ているか」(pdf)
最重要ポイントは、ニューロンの活動から細胞外電位が発生する過程の「順問題」と細胞外電位からニューロンの活動を推測する「逆問題」とに分けて考えることができるということです。図にするとこんなかんじです:
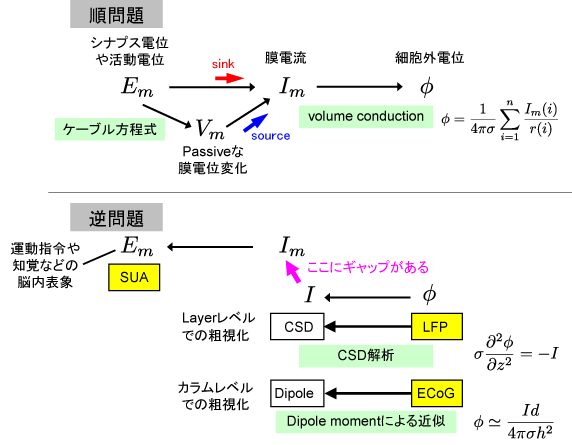
くわしいことはPDFの文書をご覧下さい。
今回のプレゼンとこの文書にはいくつか狙いがあって、1) BMIという文脈で期待されていることにきっちり答える。2) Decodingとデータベース作成に関する自分の立ち位置を明確にする。3) トレーニングコースとブログで再利用。4) チュートリアル記事の執筆依頼お待ちしております。というかんじだったのですが、当日はECoG電極のspacingの議論に質問が集中しました。わたしはECoG使ったことないんですけど、専門家面して意見言ってよかったんだろうか。
ともあれプレゼンの方はけっこうおもしろがっていただけたのではないでしょうか。あとで何人かの方に声をかけていただきましたし。私も第一旭と新福菜館と駅ビルの「すみれ」でラーメン三食食って満足。(<-食い過ぎ)
LaTeXはインストールが面倒なのでなるたけmimeTexとかでなんとかしてきましたが、今回はさすがにそういうわけにもいかないなと思ってひさびさに調べてみたら、TeXインストーラ3というのでW32TeXを含むすべてが一括インストールできるようになっていて感激しました。ほんとはアンインストールも一発でさせてほしいけど。
今回powerpointに数式を貼りつけるときにTeXclipにお世話になりましたが、これは素晴らしいですね。(いま気づいたけど作者の方はMouseoverDictionaryの作者でもあるんですね。こちらもお世話になってます。)
こんなかんじでlatexの原稿ファイルをアップロードするとPDFが作成できる、みたいなwebサービスが出てこないですかね。本一冊書くとかでなければ、そういうので全然じゅうぶんだと思うんですけど。精神としてはhtmlみたいに、コンテンツ作る人はコンテンツ作りに集中すべきで、タグ打ちとかもなるたけ省力化する方に行くのがあるべき方向だと思うんですけど。数式書くときしか必要ないからなあ。論文書くのだってバージョン管理システムを使うよりはワードの履歴機能使う方が圧倒的によいですからね。
って話がずれてきたので、終了。
# mmrl
ども、おひさしぶり、mmrlです。
現在新しい多点電極で深いところの細胞外記録チャレンジ中なのですが、スパイク波形が従来のやり方と異なってとれてきていて、スパイクソーティングに悩んでいました。
むか〜し勉強した電磁気学の教科書と、Kochのを引っ張りだしてきてうんうんうなっていたのですが、大変参考になる資料を作っていただきまして非常に勉強になります。
ご無沙汰しております。
>>現在新しい多点電極で深いところの細胞外記録チャレンジ中
おー、たぶん深いへんだとニューロンのdendriteの形状が違うからpyramidal neuron用のアルゴリズムだと最適ではないのかもしれませんね。
"Biophysics of Computation"とか"Methods in Neuronal Modeling: From Ions to Networks"とかも読んでおかないといけないのですが、けっきょくHolt-Koch論文とかですませました。
すみません。気になったので投稿します。
以前のエントリですが、
Vext = R/(4*pi)*Σ(Ij / rj) ---(3)
ただし、Vextは細胞外電極で測定された電位、Rは細胞間隙の抵抗でだいたい200-300ohm/cm。Ijはある場所jで流れた電流の大きさ、rjは場所jから細胞外電極までの距離。Σはすべての電流源jでの総和。(Nunez and Srinivasan(2006)より)
なんとなく意味は通じるのですが、空間上の一点を間接的に表現している r と一点では表現し得ない I が混在している時点で(3)の表式に違和感あるんですが。
生物系では慣例的にこういう表式するんですか?
psycho-resiさん、こんにちは。はてなダイアリー拝見しました。
物理出身で精神科医をされていて、ECTの効果を脳内でのマクスウェル方程式を解いてシミュレーションしてるということは、私よかよっぽどこのへんについてくわしいと思うんでこっちの方が教えていただきたいです。
ご質問いただいた件ですが、何段階かポイントがありまして、わたしもしぼりきれないのでそれぞれ書いてみます。
1) まず、ご質問への直の答えですが、上記の式自体は言及している教科書Nunez and Srinivasan(2006)の中でじっさいに使われている式ですし(p.168)、他の論文などでもニューロンの活動から周りの電位をシミュレーションするときによく使われる式です。
2) 新しい記事の方もそうですが、わたしの言葉遣いとして、currentとcurrent sourceや、currentとcurrent densityを厳密に使い分けていないところがあります。たとえば上記のIjはcurrent sourceと言うべきでした。このへんを正しくしたら問題は解決するでしょうか?
3) もし上記の式が∑ではなくて積分形で、微小領域dSで流出入するcurrent source I(x,y,z)の足し合わせ(∫I/R dS)だったら違和感は解消するでしょうか?
4) 電磁気学の教科書によく出てくるのは上記のcurrent sourceがpotentialを作るものではなくて、chargeがpotentialを作るものですが、こちらの場合も上記の教科書では φ = 1/(4*pi*ε)∑(qj/Rj) という表記がされています(φは電位、qjは位置jでのcharge sourceの量で、εは誘電率)。これには違和感はありますか? 積分形なら問題ないですか?
5) 細胞外電位で見ている現象は、細胞膜が静止膜電位で分極しているchargeによって影響を受けるようなミクロな系ではないので、4)のchargeの式ではなくて上記のcurrentの式を使うこと、上記のcurrentの式はあくまで細胞外電位で見ているようなmacroscopicな物理現象でのみ妥当であること、が上記の教科書には書かれています。このあたりが論点でしょうか?
以上です。ほかにもぜひこれらのエントリや文書で直すべき点、改良すべき点などありましたらご指摘いただけるとありがたいです。
いえ、あんまりわかってないです。
ECTの科学的な基礎ができてないと感じたので着手したんですが、できたのは静解析のみです。マクロの大雑把な電位勾配や電流密度はそれらしく計算できたんですが、ミクロのつまりニューロンとそれらがどう相互作用するのかは課題として残ってます。
質問に関してですが、私、その教科書持ってないので印象でしかいえないんですが、膜上の局在した領域でのみ電荷の移動がおこるという前提があるようですね。ならばそのn番目の領域dSでの電流密度ベクトルjnを定義して
In=∫jndS
細胞外電極で測定された電位Vextはこれらの足し合わせだから
Vext = R/(4*pi)*Σ(In / rn)
とすると私の場合は激しく納得します(少なくとも表記の上では。式自体はどうやって誘導されたかわかりません)。
前提の理解が違ってたらたぶん見当違いのこといってます。
(4)は違和感なし。
(5)は難しいんでまた考えさせてください。
たぶん、問題の式がわかりにくい理由の一つは、Imがcurrentやcurrent densityではなく、current sourceをあらわしていることによると思われます。current sourceは-div(Jsource)(Jsourceは膜を通過するcurrent densityのうち、passiveな抵抗性電流で説明されないもの)と定義される量で、マクロ系の電磁気学でみかけるものです。
currentやcurrent densityを考えると、電荷の偏りがないかぎり、電流は閉じているはずなので(d(rho)/dt=0 -> divJ=0)、電流がある点にしかないというのはありえないことです。しかしながら、-div(Jsource)は、点状に分布し得ます。
ここのところは、マクロ系の電磁気学の議論に類似しています。すなわち、current density(J)をオームの法則の法則に従うpassiveな抵抗性電流(sigma*E)と、それ以外(=容量性電流 + active conductanceなど)に分解します。この後者をJsourceと呼ぶことにします。J = sigma*E + Jsource。Jについては、電流は閉じているはずなので、divJ=0。
active conductanceや容量性電流などがなければ、(sigma=const.として)常に、div(E)=0で、磁場でもあたえない限り、rot(E)も0で電場もなにも発生しないのですが、active conductanceが働くと、div(E)は0とは限らず、電場および電位が発生することになります。
ここでdiv(J)=0、およびJ = sigma*E + Jsourceより、
div(E)= - div(Jsource) / sigma = (current source) / sigma。
したがって電場(phi = - grad(E))は、
phi = -Laplacian (current source) / sigma
となって、rot(E)=0より、r->infinity でphi=0の境界条件のもとで
(3)式のように解くことができます。
psycho-resiさん、どうもありがとうございます。基本的にはわたしのコメントの2)での「たとえば上記のIjはcurrent sourceと言うべきでした。」というのが論点だったようですね。
あと、教科書(Nunez and Srinivasan(2006))での記載では、たしかにI_mは点ではなくて、ある一定の大きさを持った小領域になっています。つまり、球をどんどん小さくしていって、その外側ではdiv(J)=0が成り立ち、その球面より内側ではdiv(J)=s(source current)になっていて、この球の中心からr離れた場所での電位φを計算する式となっています。この関係を使って、膜に広がって分布しているcurrent source/sinkを小領域に分割して足し合わせる、というのがここでやっていることであると理解しました。
式(3)の表現の仕方としては、ケーブル方程式で求まるtransmembrane current I_mという細胞の中から見た値が、細胞外からはsource current s (≡-div(J_source) )として捉えられていて、中からと外からの表現が混ざっているというあたりの問題なのかなとも思いました。
OK さん、どうもありがとうございます。ご指摘の「細胞膜ではdiv(J)=-I_m、それ以外ではdiv(J)=0」については、今回のPDFファイルの注5で多少言及しておりますが、以前のエントリのときはまだこのへんがわかっていませんでした。
J = σ*E + Jsourceの式に関してはOKさんの解説で非常にすっきりわかりました。どうもありがとうございます。(じつはこの式はNunez and Srinivasan(2006)のp.166で扱われていまして、該当部分を読んではいたんですが、十分理解できていませんでした。) Active conductanceとcapacitative currentだけがdiv(J) ≠ 0の点を作る、というのはとてもクリアーで、なんで膜にだけcurrent source/sinkがあるのかが明確になりました。(なんとなく、電極からは膜の向こうは抵抗が大きすぎて「見えない」から、その境界から出入りする電流はみな無から有が生まれたように見えるので、current source/sinkになる、というような理解の仕方をしていました。)
そうするとひとつ新たな疑問が生まれてきたのですが、わたしの図式では、ケーブル方程式で transmembrane potential V_mを計算した後に、transmembrane current I_mを抵抗性成分g_m*V_mと容量性成分cm*dV_m/dtの和として計算していました。その上でI_m/rを足し合わせる(式3)ということをしていました。しかし、電位に影響を及ぼすのが容量性成分だけであるなら、source currentの値としては、抵抗性成分g_m*V_mを差っ引くべきではないのでしょうか? つまり、ある位置での膜のsource current s = I_m - g_m*V_m であって、&phi = ∑ (I_m - g_m*V_m)/r とすべきではないのでしょうか? ただ、今回参照したPettersen KH, Einevoll GT. Biophys J. 2008 Feb 1;94(3):784-802では、式(3)のI_mがlinear leak currentとcapacitative currentの両方を含んでいると書いていますので、たぶんわたしがいま言ったことは間違っているんだろうと思いますが、このへんがまだよく理解できていません。
"Jsourceは膜を通過するcurrent densityのうち、passiveな抵抗性電流で説明されないもの"と書いたのは間違いでした。膜を通過する電流は抵抗性成分、容量性成分どちらも含める式で正しいと思います。
div(E)= - div(Jsource) / sigma とするときに、sigmaは一定でなければいけないので、細胞外空間だけを考えて、細胞膜のところは境界条件として扱うので、膜を通過する電流の抵抗性成分はsigma*Eには含まれず、Imのほうに入ってくるのだと思います。
OKさん、長々とおつきあいくださいましてどうもありがとうございます。これは細胞外の(σ1)と細胞膜の(σ2, ε)というふたつの領域があって、元々の話は細胞外の(σ1)の環境でのポアソン方程式を解くのに細胞膜の表面を境界条件として使うという話であって、私がケーブル方程式云々で言っていたのは細胞膜の(σ2, ε)の環境でのポアソン方程式を解くときの話であって、両者をごっちゃにしてしまったということのようです。またもう少し時間を取って勉強してみたいと思います。とりいそぎ御礼まで。
# 元物理系学生このスレ長いですね。
この話題に微妙に関係するご相談なのですが、先日投稿した論文にレビュアーからコメントが返ってきました。
かなり見当違いなコメントで正直相手にしたくないのですが、なかでも凄いのが
between the field strength in the brain tissue with ECT and possible neurophysiological responses
の関係を明らかにせよというもの。
それがなかなかできないからシミュレーションしているんでしょうに…。
それでなんですが、仮に実験系を組むとしたらどんな感じがいいかご相談よろしいですか。
考えようによっては、この分野、基礎から臨床までシームレスですね。
>>このスレ長いですね。
以前はコメントが15個並んだこともあったのですが
(two-photon in_vivo imagingのカテゴリ)、これだけ長いのも久しぶりです。
>>between the field strength in the brain tissue with ECT and possible neurophysiological responsesの関係を明らかにせよというもの。
ちょっとこれだけだとよくわかりませんが、rodentとかで電気刺激によるseizureの誘導と記録の実験とかそういった過去の論文を引いて議論すればよいのではないですか?
実験系、というとちょっとわからないですね。
今回作ったPDF文書の中ではLogothetisのNeuron 2007論文というのを言及していて、それとかは微小電気刺激をしたときの応答からconductanceを計算してたりします。ただ、たぶんECTの場合はseizureまで起こすのが必須なんだと思うんで、ちょっと違いそうではありますが。
ではまた。
どもです。現在LFPとスパイクの関係を調べてる者で、共同研究者が吉田さんのレジメを発見して来てくれたのは良かったのですが、僕も式(1)のphi = (1/4pi sigma) I_m/rで混乱してしまい、このスレで納得しました。感謝感謝。それで細かい事で申し訳ないですが、OKさんの
>>したがって電場(phi = - grad(E))は、
>>phi = -Laplacian (current source) / sigma
>>となって、rot(E)=0より、r->infinity でphi=0の境界条件のもとで
>>(3)式のように解くことができます。
はE = -grad phiと
current source = -(1/sigma) Laplacian phi
の間違いですよね。
Kensukeさん、どうもありがとうございます。ちょっともう内容を忘れかけてますが、「E = -grad phi」の式はたしかにひっくり返ってましたね。
PDFのファイルのほうも、ここで皆様が書いてくださったことを元にしてアップデートしておきたいのですが、頭がそっちに戻らないので止まってます。
Kensukeさん、物理屋さんですよね。もしよければ、PDFの式(1)のあたりの表現をどう変えるといいかsuggestしていただければ、acknowledgement付けてPDFをアップデートしたいと思いますので、ぜひご考慮いただければ。
もしくはappendixに新たに項目を設けるとか。pLatexで書いてくだされば対応できますのでぜひ。
お勧めエントリ
- 細胞外電極はなにを見ているか(1) 20080727 (2) リニューアル版 20081107
- 総説 長期記憶の脳内メカニズム 20100909
- 駒場講義2013 「意識の科学的研究 - 盲視を起点に」20130626
- 駒場講義2012レジメ 意識と注意の脳内メカニズム(1) 注意 20121010 (2) 意識 20121011
- 視覚、注意、言語で3*2の背側、腹側経路説 20140119
- 脳科学辞典の項目書いた 「盲視」 20130407
- 脳科学辞典の項目書いた 「気づき」 20130228
- 脳科学辞典の項目書いた 「サリエンシー」 20121224
- 脳科学辞典の項目書いた 「マイクロサッケード」 20121227
- 盲視でおこる「なにかあるかんじ」 20110126
- DKL色空間についてまとめ 20090113
- 科学基礎論学会 秋の研究例会 ワークショップ「意識の神経科学と神経現象学」レジメ 20131102
- ギャラガー&ザハヴィ『現象学的な心』合評会レジメ 20130628
- Marrのrepresentationとprocessをベイトソン流に解釈する (1) 20100317 (2) 20100317
- 半側空間無視と同名半盲とは区別できるか?(1) 20080220 (2) 半側空間無視の原因部位は? 20080221
- MarrのVisionの最初と最後だけを読む 20071213
# 徳生 健太郎
こりゃウケた。オリジナルのビデオでスゲえと思っただけに、パロディが爆笑。ブログ本題の「ごっこ」はどうだったかは知らんけど(笑)
# pooneilどもひさしぶり。
やっぱあの黒タイツがポイントだね。
「ごっこ」のほうはお父さんと幼児だからバランスが悪いのだけど、昨日も下の子に「へんなドボットごっこしよう」とせがまれたから、面白かったみたい。
それではまた、クラス会にて。