[月別過去ログ] 2011年05月
« 2011年04月 | 最新のページに戻る | 2011年06月 »2011年05月30日
■ Alva Noeの訳本「知覚のなかの行為」読んでます

Alva NoeのAction in perceptionについてちょぼちょぼ書いてきましたが(これまでエントリは「Alva Noeの知覚理論」へ)、訳本「知覚のなかの行為」が出ましたので読んでます。けっきょく訳本読む方が早いのよねー。
Alva Noeのsensorimotor contingency (-> dependency)は「環境への働きかけによってどのように環境が変化するかという「知識」が知覚をconstituteする」みたいな言い方なのだけれども、この「知識が知覚を構成する」って言い方が行動そのものが構成要因ではないとするための妥協の産物であって、超奥歯に物が挟まった感があるわけですが、「知識」とやらにそんなことができるだろうかと言いたくなるわけですが、「知識」を記憶、そして内部モデルとしてとらえればそんなに悪くない。
もう一度繰り返すけれども、知識=semantic memory=top-downのtemplate=内部モデルとしてとらえれば、それを知覚の構成要因とするのは悪くないんではないか。何度も書いてきたけど、ボトムアップのサプライズとそれを緩和するトップダウンの内部モデルという図式とじつはそんなに大きくかけ離れているようには思わない。ただし、内部モデルのアイデアはヘルムホルツ以降の「推論としての知覚」というアイデアと強固に結びついている。そしてNoeは直接知覚的立場からそれに強固に反対する。だから私は科学者としては内部モデル的アプローチにコミットしながら、手がかりとして哲学的な争点を脱臭するような方向へ進むことを考えながら、つまり、推論としての知覚をする際に内部に詳細な表象を作らないようにだけ考えればいいんじゃないかと。
じっさい、Raoの論文とかで出てくる図式は外部の推定をするように書かれてはいるけれども、それぞれの階層でローカルに誤差の計算をしているだけで、全体としての表象があるわけではない。だからこそ本当に問題なのはそのように分散して表象されているものをどのように行動に使うために読み出してゆくかということであって、視覚に閉じたモデルを行動に使うところまで行かなくてはいけない、っつうかもういつも書いていることと同じになってしまったのでやめる。
今日書いたことでの進歩はseosorimotor dependencyが「構成する」みたいな言い方であって、causationではないことに気づいた。あと「知識」という言い方をもっと計算論的にすればいいんだ、って考えたこと。
でもあれか、Raoのとかではまさに画像を再構成してやって、それがどのくらいうまくいっているかが議論されるんだよなあ。そこが階層ごとでなされているか、それとも全体としての画像ををpixelレベルで誤差を評価しているのか、とかうだうだ言ってないでやっぱじぶんでいじってみるべきだよな。
二つの視覚経路議論という意味では、Noeは触覚だけでなく、視覚に関しても視覚運動変換を媒介して経験が与えられるとしている。たとえば、距離や形態を「直接」経験として与えるのは要素の分析とかそういうのではなくて、「どのくらい手を伸ばしたらそれに届くか」とか「どういう風につかめるか」とかそういう「知識」によって構成されると考える。
これは神経科学者としてはけっこう興味深い提案で、ざっくりパラフレーズしてしまえば、ある物体の形態の認知はその物体のアフォーダンスの認知によって成立するという提案だ。これを神経科学と繋いでしまえば(Noeはそんなこと考えないだろうが)腹側経路の側頭連合野(IT)にある形態に選択的なニューロンの成立に、背側経路の頭頂連合野(AIP)にあるアフォーダンスに関わるニューロン(村田さんの仕事)の成立が先立つ、とまでは言わなくとも寄与する、なんて可能性を考えることができる。
これは反応性成立の前の発達のところで検証すべきことだけど。反応性がすでに成立しているadultで、AIPにムシモル入れてもITの反応性が変わるとは思えない。(もしそうなったらかなりインパクトのある仕事となるが。) LIPに平面の単純図形への選択性があるという仕事はすでにある: SerenoのNature98
まあここまで我田引水的に話を持って行かなくてもいいのだけれども、発達期の脳での相互作用というのはいろんな重要な問いがあって、NBRとかを使ってもっと積極的にやった方がよい。たとえば、幼年期の顔認識は皮質下から直接扁桃体へ入力する経路を使っているという可能性。
これもまず本当にそうか、そしてそれはどこで切り替わるのかという視点で考える。腹側経路と背側経路の相互作用についても同様な問いを複数考えることができる。ポイントはadultで反応性が成立してしまったあとで見てもわからないものを発達の過程を見ることで明らかにできないか、というような問いの作り方であって、発達自体を明らかにしたいということでもない。
本当は発達の研究というのはみんなそういった視点から捉え直すことができるはずで、ocular dominance columnとかorientation mapとかの生成の過程を明らかにすることは、どうしてそのような反応特性をニューロンが持つようになり、どのような刺激空間によってそのようなマップができたのかを明らかにすることは、adultでの応答特性とかそれがどのくらいoptimalであるかといった議論でなされていることをその形成過程からより直接的に示す仕事になるはずだ。
さらに神経科学的な捉え直しを進めてゆくけれども、Noeがあるものを(たとえば)球体として経験するのは「あるものの現れが運動の結果に応じて変化するから」と書くときに必要なのは反応選択性的なフィルターでは足りなくて、様々な向きからの見た方とかそういった確率分布みたいなものだ。
どうやってそのような分布を情報として持つことができるか。もちろんtextureの統計量みたいに次元圧縮してもいいのかもしれないけれども、それは反応選択性のパラダイムの中での簡便法だろう。(textureに関してはまた別エントリにて)
どうやってそのような分布を情報として持つことができるか。 もちろんNoeはこのような言い方はしないだろう。直接知覚派にとってはそのような多様性は現実世界そのものが持ってさえいればよいのだから。真にベイジアンな脳を考えるのであれば、確率密度分布を持つ必要があって、そのとき周辺尤度が計算できて、自由エネルギーが計算できて、フリストン仮説につながるはずなんだけど、まだちゃんと読めてないので正しいかどうか自信がない。
周辺尤度計算できるほどには情報を持ってなくて、ある程度近似的にposteriorを計算できるようになっているのだろう、というあたりが脳ネットワークにできることと考えるのが妥当なのかもしれない。ともあれこういうことがわたしはfundamentalなことであると思っていて、本当はこういうことを突き詰めていきたいし、間違っているのならさっさとその間違いに気づきたいと思っている。
Noeは「感覚運動変換のあり方についての知識」と言うとき、これが「命題的な知識」ではなくて「実践的な知識」であることを強調している。これも神経科学的な言葉へ変換可能だ。つまりこれは「宣言的記憶」ではなくて「手続き的記憶」なのだ。そうしてみると私がさっき書いた意味記憶としての捉え方は正しくなかったと言えるが、内部モデル説により近くなったと言えるだろう。まさに小脳などで内部モデルとして作られているものが手続き記憶なのだから。こうやって話は一挙に川人先生や伊藤正男先生の小脳意識説に近づいてくる。
ここで私はべつに小脳説にコミットするつもりもなくて、話は逆で、命題的記憶でない、実践的な記憶としての脳内メカニズムを探すというheuristicを使えばいいんだなということになる。そういう意味では小脳よりは頭頂連合野の方が興味深い。
“Conceptualism Revised: Through Criticizing Noë's Enactive Approach” Noeの理論と二つの視覚経路議論の関係というのはNoe自身も扱っているし、私の盲視の話も二つの視覚経路議論の範疇のヴァリアントであると言える。それをsensorimotor dependencyへの反論のように使うのはじつは私はあまり釈然としていない。
というのもそれはつまり意識としての脳は腹側経路で、それはsensorimotorとは独立している、みたいな感じで使われるからなんだけど、きっと事態はそんなふうにはなっていなくて、両者の相互作用を発達の段階で考えるべき。で、sensorimotor側が先に成熟して、それによってventralがshapeされる、みたいなのがたぶんホントのところだと思う(といいつつこれを証明すること自体が大ごと)。
だから、独立しているという議論よりはその相互作用を議論する方がたぶん良いだろうと思ってる。(<-もはや言及した話とは全然別だけど。)
(ついったに書いたことを元にして編集して作成した)
- / ツイートする
- / 投稿日: 2011年05月30日
- / カテゴリー: [Alva Noeの知覚理論] [視覚的意識 (visual awareness)]
- / Edit(管理者用)
2011年05月27日
■ BPT_モデル、意志決定の応答潜時モデル、自己組織化臨界現象
さいきんのBPT_分布とかそういう話題(たとえばBPT過程、更新過程メモ、さらに元ネタのpdf)で、応答潜時のdiffusion modelとかで見たことあるようなランダムウォークの図が出てきたので、なるほどこうやって繋がるのかとか思った。
ランダムウォークモデルを使うということは、閾値は一定で、毎度同じ大きさの地震が起こるということを仮定しているわけで、でも実際はそんなことはなくて、いろんな大きさの地震が起こって、その大きさの分布がべき乗則に従うわけだから、生成過程のモデル(process model)の部分をもっと精緻にしたりとかたぶんできるんだろう。
ある震源地で、M1からM9までいろんな大きさの地震が起こっていて、その頻度の分布はべき乗則に従っている(*)んだけれども、たとえばM7以上とかに絞るとけっこう周期があるように見える、というのが実情なのだろう。
(*あとで確認してみたら、これにはグーテンベルク・リヒター則という名前が付いていて、経験則として確立していることを知った。地震発生の頻度を自己組織化臨界現象として捉えるというのは、国内でも元神戸大の伊東敬祐先生のプロジェクトがあったのをwebで見つけたし、けっこう前から複雑系のメイントピックではあるらしい。ともあれ、ここでは意志決定の過程の話をしたい。)

以前に『歴史は「べき乗則」で動く』を読んだときに一時的にかぶれたんだけれども、「科学」の地震の特集の号(たぶん2010年8月号か?)とか読んでみたらぜんぜんそういった複雑系的な話題がなくて拍子抜けした覚えがある。
だから地震の発生頻度の予測にランダムウォークと閾値のモデルを使うというのはかなり簡略化したモデルといえるだろう、みたいな話にして終わるつもりだったのだけれども、逆に考えることができることに気づいた。
つまり、地震だけではなくって、応答潜時に関しても、ランダムウォークで一定の閾値を超えるとイベント発生というのはかなり簡略化したモデルなのであって、実際にはそんな閾値などどこにもない。急速眼球運動(サッカード)の開始コマンドを発生させるニューラルネットワークは試行ごとにいつバーストするかは入力の大きさとは一対一対応には決まっていない。つまり砂山が崩れるタイミングと同じ自己組織化臨界現象となっている。
そう考えると、一定の閾値を超えたらバーストするという心理学的モデルが実際のニューラルネットワークの挙動とずいぶんかけ離れているように見えたものだけれども、もうすこし生理学にできそうな気がしてくる。
もちろん、リアリスティックにすればいいってわけではない。CarpenterのLATERモデルが1/RTが正規分布するというモデルでかなりうまくいったことは、本質をうまいこと突いたことを示している。(LATERモデルは試行ごとにslopeが変わるだけで、試行の中では直線的にevidenceが蓄積する。よってこれはランダムウォークですらない、非常に簡略化されたモデルとなっている。)
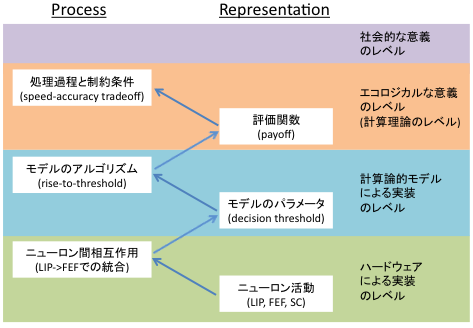
論点はそこではなくて、以前Marrの階層的に応答潜時の計算論モデルを考える、ということをブログに書いたことがある(20100317)けど、あれに関わっている。あのとき書いててぴんとこなかったのは「はたして閾値を作るニューラルネットのprocessとはじつのところいったい何だろう?」もっと簡単に「閾値の実体っていったい何だろう?」ってことだった。
そのときは個々のニューロンのintrinsicな特性とネットワーク的特性、みたいなかんじで自分を納得させていたのだけれど、ここまで考えてくると、そのネットワーク的特性というのを閾値の中に押し込まずに、自己組織化臨界現象がおこるニューラルネットワークの中でとらえる方がよいような気がする。そのうえで改めて、basal gangliaからの入力とかそういったものによる調節をとらえるならば、閾値の上下のような心理的な概念をもっとニューロンの言葉で語ることができるのではないだろうか。
とここまで書いたところで、計算論のひととかだったら当然思いつくことであるような気がした。たぶんそういったアプローチはあるのだろう。だから私としてここで強調したいのはそういったテクニカルなところよりかは、閾値という概念から砂山が崩れるタイミングのような現象への移行だ。
でも意志決定のneural dynamics的なモデルってXiao Xing Wangの(2002それから2006)とかあるなあ。でもNature neuroscience 2006を見直してみたところ、あらかじめthresholdを決めてあるようだ。まだそんなにメジャーなものはないのかもしれない。
ニューロンのバーストをself-organizing criticalityとして見るってのは”Neuronal avalanche”でやってる。neuronal avalancheじたいはスライスだけではなく、最近ではmacaqueのECoGでも見られるらしい(PNAS)。ただしここでやってることは、ECoGから大きい同期発火とおぼしき波形を検出してそれのamplitudeの分布がべき乗則に従っているとかそういう話。
つまり、まず事実レベルとして意志決定の過程での閾値のバーストというのがSOCなのかどうかを検証する段階にあるので、モデルとしてはまだ神経科学内で受容されるほどのところまでは来ていないということなのかも。(物理系のジャーナルとかには出ていてもおかしくない。)
これ以上は論文読んで考えた方がよいことだけど、ふつうにニューラルネットワークのモデルで近接したニューロンどうしてreciprocalに結合した状況で個々のニューロンに閾値があってバーストするようにしてあるだけではSOCにはならない。Criticalであるちょうどよい状態を保つためにはネットワークの相互の結合強度をそのように調整する必要があるし、それがどのくらいロバストなのかとか、そもそもcriticalである必然性があるのかとかいろいろ気になるところはある。
でも、以前言及した"Self-organized criticality occurs in non-conservative neuronal networks during ‘up’ states" Nature Physicsとか見てて、consciousな脳はcritical stateであるってのはなんだかすごく重要であるような気がしてきた。Buzsakiもたぶんなんか言っているよね。
"Rhythms of the brain"見てみた。128ページだ。これはバーストの話ではなくて、脳波の話だけど。なんだか収拾つかなくなってきたので、5章のまとめを貼って終わりとします。
Order in the brain does not emerge from disorder. Instead, transient order emerges from halfway between order and disorder from the territory of complexity. The dynamics in the cerebral cortex constantly alternate between the most complex metastable state and the highly predictable oscillatory state: the dynamic state transitions of the brain are of the complexity-order types. When needed, neuronal networks can shift quickly from a highly complex state to act as predictive coherent units due to the deterministic nature of oscillatory order.
(ついったに書いたことを元にして編集して作成した)
- / ツイートする
- / 投稿日: 2011年05月27日
- / カテゴリー: [知覚的意思決定 (perceptual decision)]
- / Edit(管理者用)
2011年05月25日
■ 僕らの人生って空飛ぶブタくんサーカスみたいだね?

昨晩次男に読んでやった絵本が心に沁みた。
アザラシが司会の動物サーカスで観客もみんな動物。最後の最後でさるくんが怪我をしたため空中ブランコができないのでこれでサーカスを終わります、と司会が言ったら観客が大ブーイング。
司会が「それでは観客のどなたかにやっていただきましょう」シーン。それではそちらのぶたくんお願いします。「ブ・タ・くん!」「ブ・タ・くん!」大成功しました!みたいなのなんだけど、なんかぶたくんに感情移入した。
たぶん、ブタくんは仕込みだったんだと思うんだけど、そのあたりのフォローもなく終わったので絵本って怖い。
ともあれ、そうやってわけもわからずひっぱり出されて、命がけの飛躍をして、しかもそれにもかかわらずに予定調和的に収束して、なんてむりやり意味を見いだそうとなんてする必要はないんだけれど、「あー、ぼくらのじんせーいーってえー、ねー、そらとぶブタくんサーカスみたーいーだーね?」「みたーいーだーね」とか思った(<-それパクリ)
昨日の絵本で、「ブ・タ・くん!」ネタが我が家で大流行。トイレから出てくるなり次男が「ブ・タ・くん!」、寝っ転がってる子どもたちをどかすために俺が「ブ・タ・くん!」、そのむかし水泳部の頃、ネタに走っていたのとやっていることはまったく変わっていない。
@a_kbysh タイトル失念してたんで補足していただけてどうもありがとうございます。ブタくん仕込み説に関してはこのページで謎が判明しました。 えほんおじさんのぶろぐ:どうぶつサーカス はじまるよ 裏表紙見逃してた。
よろこびいさんで次男に「ブタくんってサーカスの仲間だったんだけど知ってた?」って聞いたら、「いっしょにバスに乗ってたってことでしょ」と言われて、知らないのは俺だけだったということが判明。
次男の空手教室の帰りを車で迎えに行ったら生暖かい夜で、オケラがジージー鳴いていて、窓を開けてどちらから聞こえるかきょろきょろしていた。あとで次男に「何をしていたの?」と聞かれたから「オケラを探していたんだよ」と答えたら「探してどうするの?」と聞かれて、問いの深遠さに笑いこけた。
今日はフットベースのコーチ今年の第一回目。朝6時起きで6時半から8時半まで。お子さんたちに心を開いてもらうのが最初の仕事なんだけど、ぜんぜんこっちの言うこと聞かねえ。とくにうちの娘(<-だめじゃん)
Google Calenderで「6:30 フットベース」って入力したら勝手に6:30pmに設定されたので、温厚な俺様でも「屋上へ行こうぜ…久しぶりに…キレちまったぜ」とか思った。
(ついったに書いたことを元にして編集して作成した)
お勧めエントリ
- 細胞外電極はなにを見ているか(1) 20080727 (2) リニューアル版 20081107
- 総説 長期記憶の脳内メカニズム 20100909
- 駒場講義2013 「意識の科学的研究 - 盲視を起点に」20130626
- 駒場講義2012レジメ 意識と注意の脳内メカニズム(1) 注意 20121010 (2) 意識 20121011
- 視覚、注意、言語で3*2の背側、腹側経路説 20140119
- 脳科学辞典の項目書いた 「盲視」 20130407
- 脳科学辞典の項目書いた 「気づき」 20130228
- 脳科学辞典の項目書いた 「サリエンシー」 20121224
- 脳科学辞典の項目書いた 「マイクロサッケード」 20121227
- 盲視でおこる「なにかあるかんじ」 20110126
- DKL色空間についてまとめ 20090113
- 科学基礎論学会 秋の研究例会 ワークショップ「意識の神経科学と神経現象学」レジメ 20131102
- ギャラガー&ザハヴィ『現象学的な心』合評会レジメ 20130628
- Marrのrepresentationとprocessをベイトソン流に解釈する (1) 20100317 (2) 20100317
- 半側空間無視と同名半盲とは区別できるか?(1) 20080220 (2) 半側空間無視の原因部位は? 20080221
- MarrのVisionの最初と最後だけを読む 20071213