[カテゴリー別保管庫] 上丘、FEFと眼球運動
2020年08月06日
■ 盲視にipRGCsは関わっているか? アップデート2020
2012年の記事になるけど、「盲視に関わるMPKチャネルの経路」という記事で、内因性光感受性網膜神経節細胞ipRGCsが盲視に関わるかどうかについて言及したことがある。
つまり、ipRGCsって元々はnon-image-forming vision、たとえば視野全体の光量の違いによって瞳孔反射が起こるとか、そういうのにしか関わってないと思われてた。
- Neuron2010論文 “Melanopsin-Expressing Retinal Ganglion-Cell Photoreceptors: Cellular Diversity and Role in Pattern Vision”
でもこちらの論文が出て、ipRGCしかないマウス(錐体、桿体はノックアウトされてる)でも縞模様とただの灰色(どちらも視野全体での光量は同じ)を区別できる、つまりipRGCはimage-forming visionにも関わっているということが提唱されるようになった。
それで、それ以降の研究の進展をフォローしてなかったので、調べてみた。
- ARVO 2018の学会要旨"How does melanopsin help us see?"
こちらで、ちょうど2014年以降の新しい論文がリストされてる。みたところ、ipRGCだけを選択的に刺激して、錐体、桿体は刺激しないような視覚刺激を使ってその応答を見るという研究が増えてる模様。
(S錐体についても、S-cone isolating stimuliというのが使われてた。あれと同じアプローチか。)
こちらではそのような刺激条件でマウスdLGNの20%の細胞が活動する、つまりipRGCからの入力を受けているという話になってる。これは意外というか、上記の記事で書いたように、ipRGCはLGNを経由せずに、上丘や視床枕に直接入力するから、盲視に関わる可能性があるかもと思っていたのだけど、もっとふつうにLGNにも入力しているらしい。
- Sci Rep 2018 “Melanopsin photoreception contributes to human visual detection, temporal and colour processing”
あと、同じ刺激をヒトに提示した実験があって、ipRGCはヒトでの色知覚にも貢献しているらしい。
しかし「ipRGCだけを選択的に刺激」というタイプの研究は、網膜の細胞の個人差を考慮した厳密な実験条件が必要で、そのうえでも、微妙なズレが効いているのではないかという疑いが消えないので、個人的には信用してない。(S-cone isolating stimuliのときにそれを痛感したので。)
というわけで、盲視にipRGCsが関わっているか?についてはあまり大きな進捗はないようなのだけど、このネタ、マーモセットでも使えるようにはしておきたいなと思った。
- / ツイートする
- / 投稿日: 2020年08月06日
- / カテゴリー: [上丘、FEFと眼球運動]
- / Edit(管理者用)
2015年12月03日
■ Eye Tribe + Processingで視線位置のデータ収集してみた
以前のブログ記事でも書いたことがあるけど、Eye Tribeをいじってる。Eye TribeというのはUSB3で接続するポータブルなアイトラッカーのことで、アイトラッカーってのは人の視線がどこを向いているかを計測する機械。
たとえばこのアイトラッカーをパソコンの画面の下にくっつけておけば、使用者が画面上のどこを見ているかを調べることができる。だから広告業界とかにとっては重要なツールになる。もしくはマウスの代わりの入力デバイスとして使うこともできる。たとえばこの映像とか見るとゲームでの応用とかもできるのがわかるかと思う。
入力デバイスとしてのアイトラッカーのもう一つの重要な用途は、筋萎縮性側索硬化症(ALS)などで発話、キーボード入力などが出来ないような状況でも眼球運動は可能であるような場合で、この場合アイトラッカーが唯一可能なコミュニケーションデバイスとなる。しかしアイトラッカーというのは市販のもので数百万円するようなものなのでなかなか個人用で利用できるものでもなかった。
そこで廉価なデバイスとオープンソフトウェアとの組み合わせでアイトラッカーを自作しようというムーブメントが起きた。たとえばEYEWRITERではALSを発症したグラフィティアーティストTony Quanのために、プレーステーション用のカメラPlayStation EyeとC++用オープンソフトウェアopenframeworksを組み合わせることによって、視線入力によって再びグラフィティを作成することに成功している。入力デバイスとしての使用に関しては、日本語での資料としてはこちらがまとまっている:視線入力装置は自作できます。
このような状況に大きく風穴を開けたのがThe Eye Tribeというわけ。The Eye Tribeはこれまでの市販のアイトラッカーが数百万円という状況で、99ドルつまり1万円程度という廉価なアイトラッカーを出してきた。これまでのアイトラッカー(Tobiiとか)が専用のPCを使って計算をした結果をイーサネットなどで通信してデータ取得していたのに対して、The Eye TribeではUSB3接続で直で使用者のラップトップに接続できる。ゆえにポータブルにはなるけれども、多分あんまり複雑な計算はできていないはず。さてそれではどのくらいの精度が出るものか調べてみようというわけです。ということでやっと本題きた。(<-いつもよりちょっと丁寧な執筆をしております)
(私の研究での真の目的は、廉価なアイトラッカーを複数様々な位置に設置することでヘッドフリーでの広い視野角でのアイトラッキングが実現できるのではないか、というところ。今回はそのへんには立ち入らない。)
まあ論より証拠で、実際に計測した映像をアップロードしてみた。
この映像でやっているのはサッカード課題というもので、赤い丸が画面の中央に点灯して、それが左に移動して、真ん中に戻ってきて、右に移動して、また真ん中に戻ってくる。被験者の課題はこの赤い丸を追っかけてみるというもので、白い点がEye Tribeで計測した被験者の視線位置を表している。視線計測のフレームレートは30fpsで獲得して、実際の課題中にリアルタイムで自分がどこを見ているかモニターすることができている。
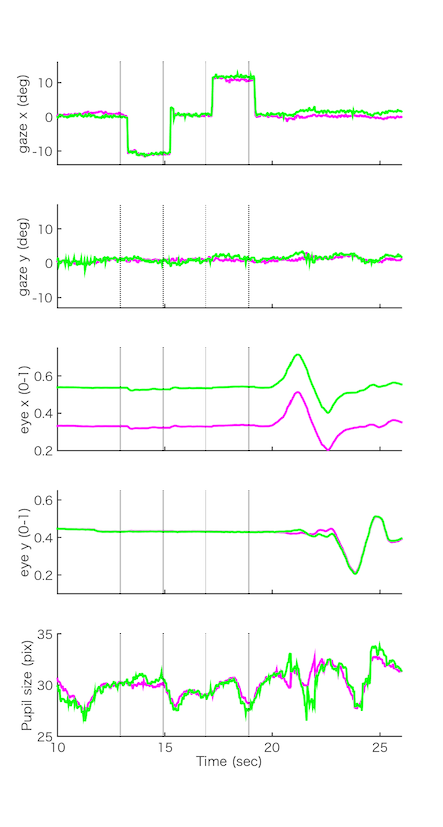
このときの計測データを横軸を時間にしてplotしたものがこちら。上からgaze(視線位置=眼球の動き)の(1)横方向の位置、(2)縦方向の位置、eye(眼球位置=頭部の動き)の(3)横方向の位置、(4)縦方向の位置、そして(5)瞳孔のサイズ。マゼンタが左眼、緑が右眼のデータ。Youtubeの動画では視線だけを動かしている10sec-22secあたりを切り出している。
眼球位置(3)(4)は安定して記録できてる。(3)を見ると左右の眼の間隔は一定なので20-23secあたりで左右に頭を動かしているときにこの間隔が変わらないことがわかる。(4)を見ると23-25secあたりに上下に頭を動かしているのがわかる。左右の眼は同じように動いている。
視線位置(1)(2)を見ると、横方向(1)に視線を移動させているのがわかる。縦線のある部分が赤点が移動したタイミング。こういう速い視線の動きのことをサッカードと呼ぶ。(<-やけに説明が親切) いっぽうで縦方向(2)には眼を動かしていないので変化はほとんど見られない。縦方向(2)のほうがノイジーなのがわかる。それでもばらつきは視野角にして1度くらいか。(真正面から真上までの視野角が90度だから、それの1/90、画面上だと27pixelに相当(測定時の画面からの距離に依存))
データ取得は30fpsなので眼球運動速度を計算するのはキビしいけど、それでもサッカードは検出できてる。マイクロサッカード(~0.5度くらいの小さいサッカード)を検出するのは無理そう。(The Eye Tribeは最大で60fpsで記録できるけど、ここでは安定性のため30fpsにしている。)
さてどうやって記録したかだけど、The Eye Tribe (99ドル)で記録したデータをオープンソフトウェアであるProcessingで表示している。Processingで視覚刺激の提示とThe EyeTribeから入力したデータの表示、記録を行っている。MacBook Air 11inchを使っているけど、Windows, Linuxでも同様な運用は可能。
The Eye Tribeが提供しているサーバプログラムを走らせると、JSON型式でのデータが30fpsで入ってくるので、それを扱えれば基本的にどんなプログラム言語でも処理可能。といいつつわたしはmatlabとpythonしか使えないので手をこまねいていたのだけど、processingでThe Eye Tribeのデータを読み込めるライブラリを作成してくれた人がいた:Processing用のEyeTribeライブラリをやっつけで作った
(ちなみに今ではほかにもProcessing3用のThe Eye TribeライブラリとしてEyeTribeProcessingというものある。こちらを使うとキャリブレーションのプログラムもProcessing上から動かすことが可能。)
このライブラリを使わせてもらって、この映像を作るためのprocessingのプログラムを書いた。ソースはgithubから入手可能なのでご自由に。(元のファイルを作成した方も「動作の保証もクレーム対応もしませんが、もしそれでもよければ使ってみてください」と書いてますので、それに準拠します。)
- / ツイートする
- / 投稿日: 2015年12月03日
- / カテゴリー: [上丘、FEFと眼球運動]
- / Edit(管理者用)
2014年09月08日
■ 生理研一般公開で視線計測の実演をします
生理研一般公開で視線計測の実演をします。2014年10月4日(土) 9:30-17:00 (受付終了16:00)場所:岡崎コンファレンスセンターおよび生理研 明大寺地区。
うちのラボでは毎回マッスルセンサー(簡易筋電計)だったのだけれど、今年はアイトラッカーを使った出し物をすることになった。はじめての試みなのでいろいろ思案してた。けっきょくあんま手をかけずに、ゴーグル型のアイトラッカー(Arrington Viewpoint)を持っているので、これかけてもらって視線計測しながらボールとか使って遊んでもらってシーンカメラの記録をリプレイすればじゅうぶん楽しめるなという考えに至った。
じつはもっと研究っぽく、投球の映像を流してそれをバットを構えて見てもらってとか考えていた。こういう研究は昔からあって、興味あったのだけれど、それのために投球の映像とかを作らないといけないのでたいへん。
そんなことせずともテニスボール投げるのをバッターボックスで見ているだけで行けるんではないだろうか。精度的に行けるか実地で試してみる。
ちょっと背景の話をすると、野球のバッティングでは「球をしっかり見ろ」と教えられるけど、じつのところ、打撃しながら球をずっと見続けることは出来ない。
まずは時間の側面から考えてみよう。ピッチャーのプレートからホームベースまでは18.44mで、球のリリースポイントから考えると17mくらい。急速が120km/hだとしたら、500msでホームまで届く。バットをスイングするのに最低150msはかかる。球のリリースポイントの視覚情報がV1まで来るのに50msくらいはかかる。このへんを合わせると、バットを振るか、どこに向けて振るかを決めるために使える時間は非常に少ないことがわかる。(参考:Sports Vision - Vision Care for the Enhancement of Sports Performance. By Graham B. Erickson p.30-32)
つぎは視線について考えるために視野角を計算してみよう。バッターからピッチャーを見た向きを0度として、球がホームベースを通るときの視野角を90度とする。ピッチャーが球をリリースした直後の視野角はほぼ0度で、球がホームベースに近づくまではこの角度はほとんど変わらない。視野角は打つ直前に急速に高くなるため、眼球運動で追従することは出来ない。このためたとえプロの選手でも球を打つ瞬間は球を見ていない。
ちょうどいいのがないのでプロじゃないけどyoutubeのこの映像:Mobile Eye Tracking: Improving Visual Perception in Baseball これよりもテニスの映像のほうがわかりやすいか:Playing Tennis with SMI Eye Tracking Glasses 原理的には同じなので。
ちなみに打撃とキャッチングでは話が違う。PLoS One 2014 Keeping Your Eyes Continuously on the Ball While Running for Catchable and Uncatchable Fly Balls を見ると、ほぼ最後まで球を見続けている。とはいえ時間スケールが違うので単純に比較できない。
クリケットの場合だとボールが近くに来る前に予想的なサッカードをしていることが知られてる。Nature Neuroscience 2001 From eye movements to actions: how batsmen hit the ball 一方で野球の場合はどうかというと私はこれまで関連論文を読んでなかった。というのもだいたいこういう論文はPerceptual & Motor Skillsに出版されるので、購読してないうちでは読めなかったもんで。
でも今回調べてたら、OPTOMETRY AND VISION SCIENCE 2014 A Method to Monitor Eye and Head Tracking Movements in College Baseball Players(pdf)という論文を見つけた。
これによると、大学野球レベルの選手では、球を見ているときはほぼずっと目は動かさず頭の動きだけで球を追従している。(眼は何もしていないということではなくて、主観的には、VORを打ち消すように眼も動かしていることになるはず。) それで、ベースを横切る最後の50msくらいだけ視野角にして10度眼を動かしている。
速度のレンジからしてこれは追従眼球運動ではなくてサッカードだろう。いろいろ面白い。ただしこの論文では打撃ではなくてテニスボールに書かれた数字と色を読めという課題なのでそれゆえの違いは出そう。つまり前者は行動のための視覚、後者は知覚のための視覚。
そういう意味では、打撃の状況で中心視と周辺視がどのくらい使われているかにも興味が出てくる。中心視と周辺視、reachingとgazeの関係に関しては五味先生がこの辺のことをやってる。JNS2006 Spatiotemporal Tuning of Rapid Interactions between Visual-Motion Analysis and Reaching MovementとかJNS2013 The Hand Sees Visual Periphery Better Than the Eye: Motor-Dependent Visual Motion Analysesとか。MFRやOFRと盲視との関係は私にとってずっと課題のままで、はやく盲視でreachingをやるべきなのだけれど。
あと、野球と周辺視というキーワードでPLoS One 2010 Transitions between Central and Peripheral Vision Create Spatial/Temporal Distortions: A Hypothesis Concerning the Perceived Break of the Curveballが引っかかってきた。これはVSSのイルージョンコンテスト2009のページで体験できる。
カーブが実際には滑らかに曲がっているにもかかわらず、なぜ急にギュンと曲がっているように見えるのかというと、gazeが周辺視から中心視に変わったからだというもので、面白いけど、だったら実地で試せば?と思う。つまり、カーブボールを見ている打者のgazeの動きは曲がったと感じたタイミングと関連しているか。
脱線しまくったけど、けっきょく一般公開の方は、頭動かして眼を測れるのだから、VORを体験してもらってもいいし、文章読んでもらってサッケードしている(追従眼球運動してない)のを知ってもらうとか、この分野の基礎を体験してもらうのがいちばん目的に叶っているんではないかと思った。
そういうわけで、カムカムエブリバディー。
- / ツイートする
- / 投稿日: 2014年09月08日
- / カテゴリー: [上丘、FEFと眼球運動]
- / Edit(管理者用)
2013年05月23日
■ 総研大講義 「注意と運動」レジメアップしました
総研大講義2013 認知と運動の脳科学 5/24(金) 「#4 注意と運動」レジメアップしておきました。
お題が「注意と運動」だったので、注意とサッカードとは同じ脳回路(FEF, SC)を使っている、とする「注意のpremotor仮説」とまではいかないけど「overt attentionとcovert attention」についての話を強調しました。時間配分を考えて、半側空間無視の話は抑えめ。
これからレジメを印刷して、ラストの部分(aberrant salience)のスライドにいろいろ足して、そして明日を迎えるというかんじ。
- / ツイートする
- / 投稿日: 2013年05月23日
- / カテゴリー: [上丘、FEFと眼球運動]
- / Edit(管理者用)
2012年12月13日
■ 盲視に関わるMPKチャネルの経路
2012/11/20
「V1から上丘へ行く経路はなにをしているか」これをアップデートした。まず、教科書的にはMPKはパラレルであるように考えられている。
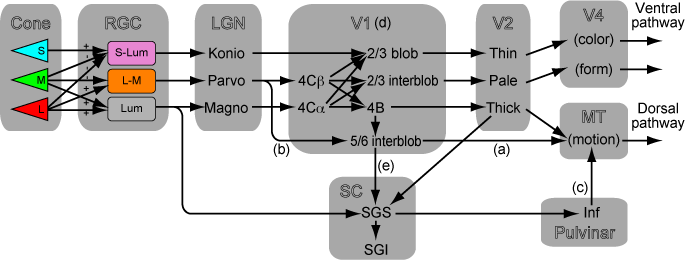
でも実際にはV1の中でかなり混ざっているってのが最近のCallawayの知見。Nat Rev Neurosci. 2009 "Parallel processing strategies of the primate visual system"
さらにV1をバイパスする経路を考えてみると、Koniocellularの系が活躍していることが分かる。
LesionしたマカクではW cell(=konio系)がマカクでSCに行っている。メラノプシン系はマウス。ラットでの知見。
ipRGCsが盲視に関わるかどうかの話をアップデートしておくと、Neuron 2010 "Melanopsin-Expressing Retinal Ganglion-Cell Photoreceptors: Cellular Diversity and Role in Pattern Vision" でipRGCsがpattern visionにも関わる(rot, cone欠失マウスでgrating vs uniformの弁別)ってのがある。
よく知られているSCNやOPNに行くやつはM1 ipRGCで(こっちはサーカディアンリズムとかに関わる)、それとはべつに、dLGNやSCへ投射するnonM1 ipRGCがある。TINS 2011 "Intrinsically photosensitive retinal ganglion cells: many subtypes, diverse functions"
Pattern visionに関わっているのであれば、ipRGCによる光への応答がどのくらい速いのかが重要だけど、PLOS Biology 2010 "Melanopsin Contributions to Irradiance Coding in the Thalamo-Cortical Visual System"を見る限りだと100msとかの潜時で出ているっぽいので充分速い。(斜め読み)
ただし、こちらはmRGCsって言ってるので同じクラスの細胞になるのかよく分からん。著者ごとに名前の付け方が違っているんだろうと思うんだけど。
Pupil reflexとかのproprioceptiveな感覚を盲視ではモニターしていて、ゆえに視覚ではなくて名状しがたい感覚になるのだ説(盲視の能力そのものではなくて「なにかあるかんじ」の説明)からすれば、ipRGC->pretectumがblink経由で情報与えてるかも。
V1から上丘への投射というのは、LGN(M)->4Calpha->4B->5/6interblob->SGSというルートがある。このため、TehovnikのようにV1 L6を微小電気刺激するとSCがドライブされてサッカードが起こる。
いっぽうでNassi et.al. Neuron 2006 "The Parvocellular LGN Provides a Robust Disynaptic Input to the Visual Motion Area MT"ではLGN (P)->V1 layer6 Meynert cells -> MTというルートでMTにもparvo入力が入る。 V1 layer6 Meynert cellsってのはSCにも投射しているかもしれない。
となると、LGN(M, P) -> V1 layer6 Meynert cells -> SC というルートでカラーの情報をV1経由でもらうことが出来るかもしれない。
昔のCoweyの論文 Neuroscience. 1995 "Nasal and temporal retinal ganglion cells projecting to the midbrain: implications for "blindsight"."を読むと、V1 lesion後に生き残ったRGCの細胞はW cell (koniocellular)であると書いてある。つまり、マカクでもretinotectalでS錐体の情報は入る。
ではなんでBrian WhiteのJNS2009 "Color-related signals in the primate superior colliculus"で上丘で速い色応答が出なかったのか。それはSGSから記録してないからじゃん?ってことになる。CatではM cellはSOへ、K cellはSGSへ入力する。Acta Neurobiol Exp 2004 "Motion sensitivity in cat's superior colliculus: contribution of different visual processing channels to response properties of collicular neurons"
なるたけコンタクト小さい電極で浅いところから記録して、DKLで刺激を作って、saturationをmaxにしてやる。つまり、backを灰色ではなくて、blueにしたうえでyellowへの応答を取る。このぐらいやれば応答が出てくるんではないだろうか?
「V1 layer6 Meynert cellsってのはSCにも投射しているかもしれない」というのは当てずっぽうではなくて、Meynert cellsだって言ってる人がいる。
上丘浅層にrabies virus入れて、V1経由で二次で染まるLGNのニューロンを調べてみればいいと思う。Konio, parvo両方が染まるはず。
Lyon 2010 "A disynaptic relay from superior colliculus to dorsal stream visual cortex in macaque monkey"でMTにrabies virus入れてdysynatpicにSCが染まる。これはinferior Pulvを経由している。2011年のIBROのときにCallawayがトークをしてたのでいろいろ質問した。うろ覚えだけど、SC->LGN->MTの可能性を聞いたら、染まっているSCの層はLGNに投射するlayerではないからunlikelyだという話を聞いた。たしかにあらためて Lyon et al 2010を読むと、そういう議論をしている(p.275左上)。SCのlayer2a->LGN(K)とlayer2b->infPVとなってる。
でもって、MTに入れて染まるのはSC layer 2b。 これでSC->PV->MT説にけっこう自信を持った。Leopordの言うようなSC->LGN->MT説は解剖学的にはunlikely。
それでもやっぱりパルボ系をSCに持ってくるのは難しい。しかし、実際にはLGNのレベルでも生理学的にはMPKは混ざっている。Logothetis et al Science. 1990 "Perceptual deficits and the activity of the color-opponent and broad-band pathways at isoluminance"ではこう書かれている:
These results suggest that impairment of visual capacities at isoluminance cannot be uniquely attributed to either of these systems and that isoluminant stimuli are inappropriate for the psychophysical isolation of these pathways.
つまり、isoluminantにしても、LGN parvoのニューロンの応答はゼロにはならない。
2012/11/20
マーモセットではretina->inf Pulv->MTがLGN->MTやV1->MTよりも早く形成される。JNS2012 "The Early Maturation of Visual Cortical Area MT is Dependent on Input from the Retinorecipient Medial Portion of the Inferior Pulvinar" 呼び水説として尤もらしい。この著者は以前にもadultでこの経路を報告している。Front. Neuroanat. 2010 "Retinal afferents synapse with relay cells targeting the middle temporal area in the pulvinar and lateral geniculate nuclei"
Eur J Neurosci. 2007 Selectivity of human retinotopic visual cortex to S-cone-opponent, L/M-cone-opponent and achromatic stimulation Human fMRIでDKL(lum, R/G, B/Y)のそれぞれの刺激で視覚野がどのようにactivation起こるか。MTの活動パターンがとても特異。B/Yでboldが下がってる。
fMRIでのBOLDがベースラインから下がるのはけっして抑制とは言えないわけだが、とにかくほかの領野とはものすごく違っていることが分かるし、やっぱLGN->MTのkonioがfunctionalに効いてんだろう、とすごく印象深い。
2012/12/4
高等研カンファレンス二日目より参加。ポスター発表終了。狙い通りDavid Leopoldと議論をすることが出来たのでミッションコンプリートした。David LeopoldはRGCからLGNとSC両方へ投射しているニューロンの重要性を強調していた。これはJNS 2008 "Y-cell receptive field and collicular projection of parasol ganglion cells in macaque monkey retina"およびJNS 2008 "The smooth monostratified ganglion cell: evidence for spatial diversity in the Y-cell pathway to the lateral geniculate nucleus and superior colliculus in the macaque monkey"のこと。
これはmagnoであって、parvoではない。retina->SCにkonioがあるとする私の持論についてはけっこうふつうに肯定していたので驚いた。ただ、上記のJNS2008二つを見たら、"lacked measurable S-cone input"って書いてあった。うーむ、いまだにS-coneがSCに入っているという証拠は見つからず。
2012/12/5
まさに俺得論文。Cerebral Cortex 2012 "Bypassing Primary Sensory Cortices A Direct Thalamocortical Pathway for Transmitting Salient Sensory information"
- / ツイートする
- / 投稿日: 2012年12月13日
- / カテゴリー: [上丘、FEFと眼球運動]
- / Edit(管理者用)
2012年11月01日
■ 上丘についていろいろ(5) transsaccadic integration
とりあえずstaircaseで自分のluminance contrastのthresholdを決める。ギリギリになってくると、画面の刺激だか、画面のムラだか、自分内ノイズだかわっけわからん。昔の網膜出血の残りで飛蚊症があるので、自分内ノイズ高すぎ。ほんとにわっけわからん。
trassaccadic integrationみたいなことを自分でテストしている。サッカード前にperipheryにある刺激をidentifyしたのか、それともサッカード後にfoveaにある刺激をidentifyしたのか、自分でやっていてさっぱり区別がつかない。
SRTから推測するに、postsaccadicにfoveaで見ている時間はほとんど無いはずなのだけれど、presaccadicにperipheryで見ているような気がしてならない。ああ、こうやって日が暮れてゆく。
もちろん、これこそが連続性を作るメカニズムなのだけれど、それを使った課題を作ってみたかったって話。
サッカード前後の知覚について論文を調べていたんだけど、MorroneとかのストーリーとDuhamelとかのremappingの話とかに終始してなんか代わり映えしないなあと思ってた。 そしたら、@kanair さんの日本語レビューを発見した。これがすごい面白かった。連続性よりも断続性を強調する、predictive codingなどへの展開、サッカードと視覚の相互拘束的関係とかみどころたくさん。感激してメールした。
ほかにもいろいろ調べてくれば出てくるのだけれど、Transsaccadic memoryとかtranssaccadic integrationみたいなキーワードでいくつか見つけた。いちばん新しいあたりだとVis. Res. 2010 "Parametric integration of visual form across saccades"とか。
サッカードでなんか出したいのだけれども、盲視に本質的なところでないとつまらない。すぐに浮かぶのはWurtzのdouble saccadeだけど、これはparietal lesion向きだと思う。Saccadic adaptationは試行数がたくさん必要なので不向き。
盲視ではtarget localizationはできるけれども、contentはわからない。そこで、transsaccadic integrationができないことと視覚意識を持てないこととを繋げられないかとか考えてる。
Deubelの仕事で、saccade中にtargetが移動しても気づかないが、targetの移動時にgapを入れると移動に気づく、という仕事がある。これはやりやすいかも。また、内観報告と眼の動きとの乖離も期待できる。まずはsaccadeなしで移動が弁別できることを押さえてから。
ただ、それだったらWurtzのdouble step taskのほうに持って行った方がキャッチーであるとは言える。意識との関連ではなくなるが。もしSC->MD->FEFが必須なら、V1 lesionでも起こるはずだ。だが起こらなかった、という結果になればサプライズが生まれる。
でもって、internal modelの維持には意識が必要であるってストーリーとつなげる。よし行った!(<-毎度の皮算用)
saccadic suppression, visual stability, internal modelこのへんでWurtzストーリーに乗っかったうえで、どっかで異議申し立てが出来ればよい。
Wurtzのストーリー的には、saccadic suppression(SS)はSGI->SGS->Pulv->MTとなっていて、corollary discharge(CD)はSGI->MD->FEFとなっている。
WurtzストーリーのままならV1 lesionによって、CDもSSも変わらないはずだが、どちらかだけ変わってくれるとインパクトがある。たとえば、CDは使えるがSSは効かなくなるとか。もちろん盲視でSSやCDをどうやってみるかってところがchallengeであって、簡単ではない。
- / ツイートする
- / 投稿日: 2012年11月01日
- / カテゴリー: [上丘、FEFと眼球運動]
- / Edit(管理者用)
2012年10月26日
■ 上丘についていろいろ(4) motion selectivityつづき(+LGN)
生理研研究会初日、無事終了。だがそれとはまったく関係なく、書き溜めたエントリを公開モードにするのであった。
Neuron 2006 "The Parvocellular LGN Provides a Robust Disynaptic Input to the Visual Motion Area MT"これ見逃してた。
Callawayのrabies virusの仕事だけど、ふつうMTへの入力はmagno/konioだけだと思われているけれども、LGN(P) -> V1 layer 6 (Meynert cells) -> MTってのがあるそうだ。Schmidみたいな灰白質除去だと残るかも。
目的はそっちではなくて、LGNがどのくらいdirection selectivityを持っているのか、だった。つまり、motion energy modelの文献を読んだ限り、spatiotemporally separableな受容屋を持ったニューロンはどの方向にも応答する。
そうすると、たとえばon-centerのRFを持ち、posi-negaのinpulse応答を持つニューロンは白-黒のエッジがどっちに動いても応答する。ただし極性が反転すると応答が変わるが。あと、ST-orientedでないのだから、staticなものが最適刺激にはなる。
いっぽうで、経験上、SGSのニューロンはほとんどがon-off両方にtransientに応答する。つまり、on-centerとoff-center両方からの入力を受けている。だから、どんな方向の刺激でも同じように強く反応すること自体は説明できる。ただし、staticなものとの違いはわからない。このへんはSGSよりLGNのほうが研究が進んでいるはずなので、LGNのdirection selectivityを調べることにしよう。
catだけど、LGNでsecond-order motionに応答するってのがあった。"Temporal response properties to second-order visual stimuli in the LGN of cats" ただし、前述の通り、catとmonkeyではこのへんはずいぶん違うので留保が必要。spatiotemporally orientedであることは、spatial frequencyへのselectivityとconfoundするので、話がややこしい。
Vis Neurosci 2002 "Are primate LGN cells really sensitive to orientation or direction?" 20%で方向選択性あり。SGSとはずいぶん違う。
そんなにSGSとも違わないか。古いのに当たる。EBR 1979 "The responses of magno- and parvocellular cells of the monkey's lateral geniculate body to moving stimuli" 方向選択性はあまりない。Magno系はsharpなvelocity tuningがある。
@ichipoohmt direction(方向)は向きがあるから、もし0degを右向きに定義したら、180degは左向き。orientation(方位)は傾きだけだから、0degと180degの区別は無しで、横棒。
@ichipoohmt 神経生理学の場面だと、縦棒を右に動かしたとき(0deg)と左に動かしたとき(180deg)で反応に差があれば方向選択性がある。向きによらず縦棒に反応するけど、横棒には反応しなかったら方位選択性だけがある、という説明でどうでしょうか。
- / ツイートする
- / 投稿日: 2012年10月26日
- / カテゴリー: [上丘、FEFと眼球運動]
- / Edit(管理者用)
2012年10月04日
■ 「カエルである」ってどんなかんじかな?
カエルの視覚について調べてる。盲視について「カエル脳の復活」みたいなわかったようなことを言うわりにはカエルの視覚のことほとんど知らないことに気付いたから。
これまで知っていたのはせいぜい
- Lettvin et al 1959 ("What the Frog's Eye Tells the Frog's Brain" bug detector論文)
- David IngleのScience 1973 "Two Visual Systems in the Frog " (Goodaleの二つの視覚システム説の先駆け)
くらいだった。
ところが、カエルの場合の二つのシステムというのはtectum(=上丘->orienting)とpretectum-thalamus(->avoidance)となっている。pretectumといいつつ、こっちはgeniculocortical系の前進なんだろって思ってた。
でもざっと調べてて、JP Ewertの一連の仕事があるってのを知った。Wikipediaだとこのへん:
神経生理学の仕事なんだけど、行動的にはニューロエソロジーなんで、出てくるジャーナルが違ったりして観測範囲から外れてた。
でもって、この図を見て混乱してきた。Pretectumからtectumにpresynaptic inhibitionが来ている。これ自体はIngleのもうひとつのScience 1973で分かっていたことをトランスミッターレベルで明らかにしたということらしいのだが、ともあれ、これでtoadのpretectum-thalamusをprimateのLGNとは同一視できなくなった。PrimateではSC->LGNという結合はあるが、LGN->SCは知られてない。
{kind=link}
Goodaleの仕事にあるように、盲視ではobstacle avoidanceは残存している。(ただし、delayを入れるとその能力は消える) PNAS2009 これをIngleのストーリーとつなげようとすると、SC->orientingで、LGN->dorsal pathway またはSC->Pulv->dorsal pathwayでobstacle avoidanceなんて話になる。
Primateだとpretectumは視覚情報処理的にはあんまたいした仕事してないので(accommodationとかblinkとか)比較的無視されがちなのだけれども、盲視における役割として考えてみようと思う。
以前考えたことがあるのだけれども、盲視での「なにかあるかんじ」というのがなんらかの体の応答を(prorioceptiveに)モニタしている可能性というのはずっとあって、もしpretectumが寄与していて、accommodationとかがそのときに変わっているかもしれない。Pupil sizeの応答も知られてる。輝度揃えてもなるかは不明。
EwertのBBS1987ってのがあって、これは読む価値あるだろうけどアクセスできない。驚いたのはDaniel Dennettがこれにコメントしていることで、哲学者なのにすげー実験の知見を勉強している。
デネットのコメント、ざっと読んだが、要はホムンクルス/カルテジアン劇場批判で、カエルは単純な視覚運動変換しか持っていないのでZombieに見える。ヒトと脳はそんなには違っていないのだから、ヒトにホムンクルスがあるように思わせるのは脳ではなくてその行動なのだ、みたいな話か。(正確には、カエルの話だけに終始してる。) なんかカエルとprimateでの脳の可塑性の違いとかいろいろ突っ込みたいところはあるが、また読み返してみることにしてみよう。
- / ツイートする
- / 投稿日: 2012年10月04日
- / カテゴリー: [上丘、FEFと眼球運動] [視覚的意識 (visual awareness)]
- / Edit(管理者用)
2012年07月16日
■ 上丘についていろいろ(3) motion energy周辺
一年くらい前にツイートしたことを再編集して掲載。
なんでblindsightでmotion刺激が有効なのかをちゃんと考えるとどうもわからない。MotionはLGN->MTでもよいように思う。上丘のomni-directionalなニューロンからPulv経由でMTの応答ができているとは考えにくい。
IBROでトークの後のWurtzを捕まえて(みんな粘って質問してた)話を聞いてみた。Wurtzが言ってたのは、Pulvから記録して、MT刺激でantidromicが出るやつ(MT->Pulvの入力があるやつ)は方向選択性があって、アンチがないやつは選択性がない。
これはBerman 2008のfig.4になっている。Wurtzはsaccadic suppressionについてはSC->Pulv->MTを強調しているので歯切れ悪そうにしてたけど、これだと方向の情報はMT->Pulvと来てそうだ。
あと、盲視との関連に関しては、SC->LGN->MTの可能性もあるよと言ってた。これはSchmid論文にも書いてあることだけれども(Wurtzが言い出したことかもしれない)、前の日に聞いたEd Callawayのトーク(Neuron論文)でtrans-synaptic labelを使った仕事では、MT<-Pulv<-SGS(layer3b)はあるけれども、MT<-LGN<-SGS(layer3a)はないという話を聞いたばかりだったので、その話をしたらWurtzは意外そうにしていた。このへんは論文に書く必要が出てきた。
でもって、そういう目でもうすこしmotion processingについて勉強してみようと思ったらまた広大な領域が開けてしまって途方に暮れる。つまり、Adelson and Bergenみたいなmotion energyによる説明とdisplacementとをちゃんと分けないといけない。あと1st order, 2nd orderとか、phi motionとreversed-phiとかいろいろ重要な知見がある。(このへんの論文読んでると西田さんの名前が頻出する。)
盲視のGY氏はmotion刺激は定位や検出はできるが、方向弁別の能力はそれと比べると著しく低い(Azzopardi 2000)。つまり、上丘のomnidirectional cellっぽい。
reversed-phiに関してはneurophysiologyだと、AlbrightのJNP2005がMTの応答を記録してる。Rodmanの仕事で、上丘を抑制してもMTの応答は抑制されないってのはあるので、SC->Pulv->MTがmotionの情報そのものは持っていない。
Wurtzのストーリーは、SGIのsaccadeコマンドがSGSに戻ってきて、それがPulv->MTときて、MT/MSTで見られるsaccadic suppressionを作っているというもの。SGI->SGSの流れに関しては伊佐研のMeeさんのJNS&PNASを援用している。だから、motionの情報そのものを持っている必要はない。
いま混乱しているところ。いろいろパーツを並べてみる。motion energy vs. displacementと同様な話なんだろうと思うのだけれども、Vision research 2000 Brown & He (PDF)とか。
あー、わからねえ、Reichardt Detectorみたいな時間遅れつき差分計算と、motion energyモデルみたいなspatiotemporal filterでやってることは等価か?というかMarr的な意味で説明のレベルが違うという理解でよいのか?
@ykamit ありがとうございます。Adelson and Bergen (1985)を読んでみたら、fig.18でこの議論がなされていて、Reichardt Detectorのうちのあるバージョンでは等価になるとのことでした。
@ykamit ありがとうございます。そもそもわたしはReichardt Detectorがもっと単純な足し算だと誤解してたので(dendriteへの入力のイメージ)、神谷さんの指摘で道が開けました。
Reversed-phiがmotion energyで説明できるってのはGeorge Matherのサイトを見て納得した。子どもたちもこれ見ておもしろがってる。さらにtwo-stroke versionというのがシンプルかつ効果的で驚く。こちらもmotion-energyで説明できるらしい:"A motion-energy model predicts the direction discrimination and MAE duration of two-stroke apparent motion at high and low retinal illuminance" (PDF)
やっと思い出した。つまり、スタート地点はなんだったかというとPaul AzzopardiのPNAS2011で、盲視のGY氏にmotion illusionを見せたら、feature-basedが消えてmotion energyに応答した。
だからじつはretinotectalでmotion energyやってるかもよ、っていうけっこう衝撃的な主張なのだけれども、共著のHoward S. Hockという人が使っているこのillusionの説明がさっぱりわからないので難儀していたのだった。
supplementaryもなければ、著者のサイトにもなにも置いてない。勘弁してほしい。JoVとかだとwebにquicktimeとか貼ってあったりしていろいろと捗る。
Current Biology論文にも出したように、盲視でもsaliencyは使えるのだけれども、それをひっくり返して、盲視ではsaliencyしか使えない、という作業仮説を立てている。たとえば、色パッチ(灰色背景での等輝度の赤)の定位はできるけれども、色パッチの弁別(等輝度、等saturationの赤と緑)はできない。
そうすると、motionでも同じようなことが期待できる。motion刺激のcontent(方向)はわからないけれども、motion刺激の定位はできる。つまり、saliency mapでピークができるようなものは処理できる。そうして調べていたらぶつかったのが、Li and SperlingのNature 1995で、saliency-basedなapparent motionというのがあって、これが3rd orderで、あと1st, 2nd orderがあるというもの。わっけわからん。
George Matherのサイトにmatlabコード付きでmotion energy modelの解説があった。これで勝つる。
Ittiのsaliency modelでのmotion channelはこのmotion energy modelを使っている。つまり前後のframeの値を使う。ではIttiのbayesian surpriseのほうはどうか。基本的におんなじdetectorを使っているはずなので時間差分のさらに差分を取るようなことをしていることになって具合が悪いはずだ。本人に聞いてもそんなにformalな解説が期待できないので(というか私が理解できない)、いつかコードを読むべきか。
Azzopardi PNAS2011の刺激を理解した。というか刺激そのものが図に書いてあった。あまりに単純すぎて誤解してた。長方形が伸び縮みするだけだった。伸びる方向に動いているように見える。しかし、伸びる部分の輝度を反転させると、motion energy的には逆向きに動く。
そしてGY氏の損傷視野でだけ、この刺激が逆向きに動いているように見える。だいたいわかった。
- / ツイートする
- / 投稿日: 2012年07月16日
- / カテゴリー: [上丘、FEFと眼球運動]
- / Edit(管理者用)
2012年05月05日
■ 上丘についていろいろ(2) motion selectivity
nhpの上丘はmotionの方向に依存しないサプライズ検出器みたいになっている。しかしFigure-backgroundでのmotion detectionの場合にはMT損傷でこれが消える。Exp Brain Res. 1997 "Loss of relative-motion sensitivity in the monkey superior colliculus after lesions of cortical area MT"
ちなみにcatの上丘ではdirection selectivityがある。macaqueの上丘ではあまりdirection selectivityがないという話の元ネタを探すためには1970年代まで遡る必要があって、J Neurophysiol. 1977では"Movement in any direction was as effective a stimulus as any other in 125 cells (83%)" とか書いてあったり、EBR1979では"The pan-directional cells which comprise about 90% of the visual cells in the superficial layers" とか書いてあったり。
0なわけではないのだけれど。それから、velocity sensitivityは非常に強い。Catの場合だとこれを使ってmagno系(Y)かkonio系(W)かを分ける。
方向依存のないサプライズdetectorを作るためには、そのまえにdirection selectivityのあるところが必要で、先述の通りMTが必須かもしれない。すると、blindsightにおいても、RGC->LGN->MT->SCみたいな経路を使えるかもしれない。
@ichipoohmt ネコもウサギもハトもカエルもみんなdirection selectiveだったはずなので、サルの方がその能力を失ったと考えた方がよいように思います。ただし、上丘がその能力を失ったというよりは、ウサギとかではRGCのレベルでmotion detectorがあったのに、それがnhpとかでは失われたという風に考えた方がよいようにも思います。
もっとちゃんと調べておいたほうがよいのですが、1)RGCに方向選択性のある動物ではMTだろうがSCだろうがそのコピーで情報を持つことができた。2)RGCでの方向選択性がなくなって、V1->MTを使った処理が専門化した。3)さらにそこからinvariantな情報が上丘にできた?
@ichipoohmt MTのある視野位置のいろんな方向検出器を統合してSCの方向サプライズ検出器を作っているとしたら、受容野がMT並に広くなるはずなので、それで検出できそう。
そういう眼で先述のEBR1997を読み返してみたら、読みまちがえていることに気づいた! MT lesionで消えるのはrelative-motion (図と地のあいだでのmotion)の選択性だけで、単純なmotion関連の成分(motionによる)は変化しない! となると今まで書いていた話が全部吹っ飛んだ!
@ichipoohmt おつきあいありがとうございました。今書いてたのは、犬山で話をした、盲視ザルではmotion saliencyが非常に強い、これはなぜかっていうことを考えていたのでした。
Relative motionの話の大元はJ Neurophysiol. 1991 "Selectivity for relative motion in the monkey superior colliculus." ランダムドットを提示して、バックグラウンドが均一な灰色だと方向選択性がないor弱いけど、バックもランダムドットだと選択性が出てくるという話だった。
だから、私がやっているようなmotionによるsaliency/surprise検出に関してはじつはこのrelative motionが効いてそう。ただし、Laurentのモデルは単にローカルにmotion energyを計算しているだけだからrelativeは考慮に入れてない。もちろん両者は刺激としては相関しているので結果としては有意に出てくると考えるのは悪くない。
ん? そうしてみると、さっきの図式をちょっと修正すればよいのか。Macaqueでは、SGSでの単純な方向選択性は非常に弱くて、RGC->SGSもMT->SGSもあまり寄与していない。Relativeな方向選択性、つまりcenter-surroundの計算をするときだけMTに依存。
だから、マカクSGSではRGCが方向選択性を失った分をMTからの入力で補完したりしてない。でもCat SGSには強い方向選択性があるから、これがMT由来なのか、RGC由来なのかが重要。もしMT由来だとしたら、じゃあなんでマカクではそれを失ったのよ?って問いになる。 ここまで来れば次はcat RGCでの方向選択性を調べてみることにしよう。
ざっと見るに、Vis Neurosci. 1995 "Spatio-temporal receptive-field structure of phasic W cells in the cat retina."ではcat RGC W cellで10/54はdirection selectiveと出ている。
だいたい満足できた。
- / ツイートする
- / 投稿日: 2012年05月05日
- / カテゴリー: [上丘、FEFと眼球運動]
- / Edit(管理者用)
2012年04月28日
■ 上丘についていろいろ(1) オシレーション
BBS2007で上丘が意識に関連してる説がある。"Consciousness without a cerebral cortex: a challenge for neuroscience and medicine" 水頭症で大脳皮質のない幼児が意識を持っているとおぼしき行動をする、というショッキングな例も出ている。外在説的考え方からすれば、可能な知覚運動変換の能力に従ってさまざまなレベルの意識経験があってよいように思うので、意識の生成要因を脳のある特異な構造に結びつけるアイデアにたいする反論みたいになる。私の仕事でもどっかで言及する必要が出てくるかもしれない。
このBBS論文の中では、Watkins et al 2006 (Geraint Rees)というのが言及されていてaudio-visual illusionを経験したときだけSCの活動が上がる、という話が出ている。cortico-tectalでなんか統合やってるんでしょうというのはcortico-pulvinarでbindingやってるって説と同じで、あってもいいけど本当のことはわからない。
ただ、上丘でのoscillationに関してはやっぱやっときたいなあと思う。CatではBrecht et al (W Singer)の一連の仕事というのがあって、麻酔下ではcortico-tectalでalpha-beta辺りが出る。JNP1998 "Correlation Analysis of Corticotectal Interactions in the Cat Visual System"
面白いのは、awakeのときだけgammaが出るって話。"Synchronization of visual responses in the superior colliculus of awake cats." これでalphaとgamma両方あるとなると、Tallon-Baudryのこれを思い出す。JNS2009 つまり、gammaがawarenessでalphaがattentionで、どっちもpre-stimulusで出てくるけど、影響の出方が違う。
これはあくまでvisual cortexでの話だけれども、私の話でも上丘ニューロンのhit-missの差はprestimulusにも出てきていて、しかもphasicなので、なんらかoscillationっぽいことが起こっていて、それとの位相によって行動が決まっている節がある。
あいにくdetectしたスパイクのデータしか残ってないので、いまは議論のネタにしかならないけど。ともあれ、前述のJNS2009はprestimulusとvisual responseのcorrelationをとってきっちりとした議論をしているので、これは参考にしたい。これは先日書いた、Victor Lammeの質問へのいちばんしっかりとした答えになるはずだ。
いま気づいた。上丘ガンマ論文のファーストのMichael Brechtってこの人だ。Nature 2004 "Whisker movements evoked by stimulation of single pyramidal cells"
- / ツイートする
- / 投稿日: 2012年04月28日
- / カテゴリー: [上丘、FEFと眼球運動]
- / Edit(管理者用)
# tkitom
>あいにくdetectしたスパイクのデータしか残ってないので、
Instantaneous firing rateをアナログデータとして扱ってやってみるという手は使えないでしょうか?取っ掛かりとしてでも。
ども。たしか元基生研の廣川さんにもそう言われました。最近の小林康さんの仕事とかもそういうかんじですよね。論文のデータにはしにくいけど、行けるかどうか見るためには使えると思います。じつはpre-stimulusは見た感じあんまoscillatoryではないです。
# pooneilつーか、FBユーザーはぜひFB用コメント使ってください。そのほうが手元に残るし。けっしてエロいサイトへのリンクとか飛ばしたりしないので。
# tkitom>つーか、FBユーザーはぜひFB用コメント使ってください。
それは作法を知らずにすみませんでした。こちらはこちらなのかと思っとりました。
今度からは気をつけます。
いや、失礼、どっちでもいいんですがたぶんそっちが便利ってだけの話です。
2012年04月03日
■ 上丘と周辺視
リモートカメラだと頭がある程度動いていいとはいっても、30度とか動いたらもうデータが取れなくなる。複数のカメラを横に並べてやれば、広い画面の探索に対応できる。そういうのはコマーシャルレベルでは見ないが、研究レベルではググると出てくる。生理学出来ない環境のにネタのストックにしとく。
モニターの中を見ているくらいの狭い探索はマカクやチンプにとってはたぶんそんなにエコロジカルには重要でなさそう。上丘のマップを考えてみても、頭と目を動かして広い範囲を探索するのに最適化されているだろう。
上丘のマップの話を補足すると、JD Crawfordの一連の仕事 (たとえばNature Neurosci 2001) があって、上丘の眼球運動のベクターをコードしているのではなくて、網膜のどの位置に目を動かすかをコードしている。上丘のベクターマップでは偏心度の大きいところは非常に小さいけどこれは端的に60degとか大きいサッカードは普通はしなくて、頭を動かしてorientするからで、頭+目=網膜の位置の変位で考えると、大きな角度変化も上丘でコードしていることがわかっている。
Fovea付近の視覚の詳細な分析というのはたぶん大脳皮質の機能なので、saliency mapに基づいた視覚探索における上丘の役割ということを考えると、fovea中心主義は大脳皮質の機能を前提とした視覚科学の流儀をそのまま安直に使っているのではないかという恐れもある。
そういう点から考えると、色とか方位とかによるsaliencyってのはほとんど中心視野でしか使えてなくて、周辺視野で使っているsaliencyってのはmotionかflicker (onset, offset)くらいに単純化されるだろう。
つまり、staticなsaliency (frameごとにpixel間の関係だけで計算できる)はほとんど中心視野用で、dynamicなsaliency (luminanceやらいろんなものでdefineされた時空間的なエッジ)だけが周辺視野で使える。
たぶん心理物理でこういう研究はされているのだろうけど、motion saliencyが使えるとして、それは輝度、色、orientationみたいにいろんな属性でdefineされるわけで、周辺視でどのくらいのものが使えるかとかそのへんを抑えておく。
周辺視では赤背景の中を動く等輝度の緑丸とかの弁別能がどのくらい悪いかとかはvision researchの昔の論文とか探せば出てきそう。
- Second-order motion perception in the peripheral visual field.
- Psychophysical evidence for separate channels for the perception of form, color, movement, and depth
- Position-based motion perception for color and texture stimuli: effects of contrast and speed.
そういう目で見るとこれとか面白い。 Peripheral vision: Good for biological motion, bad for signal noise segregation? 周辺視はバイオロジカルモーションが使える。
つまり、周辺視は皮質下のシステムが使えて、荒い空間解像度で早い時間応答でバイオロジカルモーション検出したり、affective blindsightみたいに感情の識別が出来たりする、なんて話になったら面白い。根拠がそんなにあるわけでもないけど。
前述の論文は渡邊克巳さんの論文 [Vision Res. 2005] の結果にたいする反論でもあって、ノイズに埋もれている場合は周辺視ではバイオロジカルモーションは使えない。
[Neuropsychologia. 2009] これを見ると、STSのbiological motionに関わる領域とEBA(extrastriate body area)はぜんぜん離れてる。うーむ、まだよくわからないな。
- / ツイートする
- / 投稿日: 2012年04月03日
- / カテゴリー: [上丘、FEFと眼球運動]
- / Edit(管理者用)
2012年01月09日
■ ハトの首振り
ハトの首振りを止める方法 - Yahoo!知恵袋 これ面白い。あのハトが地上歩いているときに頭を前後にヘコヘコするやつ。あれはヒトの眼球運動で起きていることと似た現象なのだな。
ヒトがなにかものを注視しているときに不意に頭を回すと、眼球は網膜像を変えないように頭の動きとは反対側に回る。これを前庭動眼反射VORという。また、動いているものを見るときにはヒトでは眼球が動きに追従して流れてゆくフェーズとずれた分を戻すフェーズとがあって、これをOKNと言う。
これと同様にして、ハトの首振りも歩行による視野の動きを打ち消すように後ろに動くholdフェーズと後ろに下がった頭を前に戻すthrustフェーズがあるらしい。ヒトのVORで頭の動きを眼球で補正するのと同様に、ハトの首振りでは体の動きを頭で補正している。
ハトの首振りについてのレビュー論文を見つけた。"Head-bobbing"って言うそうな。 PDFもリンクされてるけど、本文とビデオがあるこのページを読む方が役に立つ。
これのFig.1を見ると、holdフェーズとthrustフェーズの違いがよく分かる。Holdフェーズが平らになっているのは、視野の動きをほぼ0に抑えていることを意味している。つまり、視野のぶれを無くすることによって、地面にあるもの(エサとなる穀物とか)を見つけるのに役立つ。
面白いなと思うのは、thrustフェーズの方も無駄ではないということだ。運動視差を作るのに役立っているって説があるそうだ。ハトは目が横に付いているので両眼立体視がほとんど役に立たない。だから、奥行き方向の視覚情報を得るためには運動視差を使うしかない。
運動視差というのは、電車に乗っているときの景色の動きを思い出してくれればよいのだけれども、自分が動いているときには近くのものは大きく動いて、遠くのものはあまり動かない。これがあれば奥行き方向の視覚情報を得られる。
だからthrustフェーズは頭を前に動かすことで運動視差を作るのに役立っているのではないかという話があるんだそうな。これは面白い! じゃあ、逆にヒトでのfixation-saccadeの連関のうち、saccade自体はそういう意味で視覚に積極的に関与している可能性はないだろうか。
とは言っても、transsaccadic memoryとか、サッカードによって全体を把握するとかそういうのはあるとして、サッカードの動き自体を使ってこそpick upできる視覚情報というのはないのだろうか?
このレビュー論文にはほかにもいろんな面白いことが書いてあって、ではなんでハトは首振りするのにアヒルは首振りしないか。いろんなトリで比較した研究があって(322種!)、目の位置とか、歩きながらエサを探すかとか、いろんな条件が考察されてる。このようなデータから首振りの機能を推測する。
大元のYahooの記事の話に戻ると、こういった学問的な話はさておき、それを「どうやったらあのキモイ首振りを止めることができるだろうか? たとえば目隠ししたらどうなる?」みたいなかんじで料理してあるのでナイスだと思った。プレスリリースとか大学の授業とかそういうところで役立つ技だ。
- / ツイートする
- / 投稿日: 2012年01月09日
- / カテゴリー: [上丘、FEFと眼球運動]
- / Edit(管理者用)
2007年09月15日
■ V1から上丘へ行く経路はなにをしているか 補足
Retinal ganglion cellはLGNだけでなく、上丘へも投射しているわけですが、RGCの細胞の形態と発火特性による分類というのは非常に進んでいるので、すこしこれについて補足を。
まず、元ネタはThe Cognitive Neurosciences III (2004)の"Origins of Perception: Retinal Ganglion Cell Diversity and the Creation of Parallel Visual Pathways" Dennis Daceyより。ちなみにこのDennis DaceyというひとがLGNのkonio layerに入力するblue-on cellがsmall bistratified cellという細胞グループであることをを示したNature 1994のファーストです。
LGNおよび上丘にretrograde tracerを注入して、retrogradeにlabelされたRetinal ganglion cellの反応特性を見たあとでdendriteの走行などのmorphologyを見てやる、というような仕事があって、macaqueのRGCは形態から17通りとかに分類されていて、メジャーなのがparasol cell (magno系)とmidget cell (parvo系)とsmall bistratified cell (konio系)で、これらでRGCの全populationの約75%を占めてます。これらはみんなLGNにしか行ってない。
上丘へ行ってるのは、recursive cellというやつで、これはdirection selective。SGSのニューロンはmacaqueではあまりdirection selectiveなものはないので、おそらくこのrecursive cellというのはSGSの応答のsourceとは考えにくい。もちろん、SGSへの入力にconvergenceがあればdirection selectivityがなくなってもいいんだけど。それから、べつのpopulationでmoderate monostratified cellというのがあって、これはあまり生理学的にはcharacterizeされていない。
以上の、上丘へ投射している細胞は全RGCのpopulationのうち約9%です。とても少ないと考えた方がよいでしょう。また、foveaかperipheralかによってもちがうでしょうし、上丘への投射は視野のnasal側かtemporal側か(鼻側か耳側か)によってassymmetricになっていることが知られています。(Rafalのhumanの仕事はこれを論文のロジックの重要なところに使っています。)
あと、現在の文脈で重要なのは、Melanopsin-containing cellです。これはさいきん一挙に話題となってますが、ほかのRGCとちがって、自分でmelanopsinを持っているので光感受性になってるのです。ふつうのRGCはrodやconeが光を受容して、それをhorizontal cell, bipolar cell, amacrine cellなどで処理したあとの情報を受け取るだけなのです。それで、このMelanopsin-containing cellは視交叉上核(SCN、サーカディアンリズムを持ってる)を介して松果体に情報を送っていて、サーカディアンリズムとか、pupil reflexとかに関わっていると考えられています。
このMelanopsin-containing cellが自分のmelanopsinだけでなく、cone, rodからも入力を受けていて、L+M-SのシグナルをLGNに送っていることを示したのがDaceyのNature 2005 "Melanopsin-expressing ganglion cells in primate retina signal colour and irradiance and project to the LGN"です。この細胞は上丘へは投射していませんが、pretectumへは投射しているので、もしかしたらこれがextrageniculate pathwayでcolorの情報を担っているのかもしれません。
さて、おつぎの元ネタはAnnual Review of Neuroscience 2000 "The Koniocellular Pathway in Primate Vision" Stewart Hendry and Clay Reidです。上丘のSGSからLGNへの入力というのはLGNのkonio layerにのみ終止しているということが知られています。LGNのKoniocellularのlayerというのはmagno, parvoのlayerの隙間にあるわけですが、このうちいちばんventral側、つまりmagno layer側のkonio layerだけにSGSからの入力が来ています。
あと、"The ventral-most K layers, however, are innervated by types of γ and ε ganglion cells, similar in several ways to the types of cells that send their axons to the superior colliculus and pretectum."なんて書いてある。またややこしくなってきた。いったいどういう種類のRGCの入力が上丘に来てて、それがさらにLGN konio layerにどういう情報を送っているか、という問題ですね。上丘にkonio系の入力が来ているとは思えないのだけれど。
あと、こんなふうに言ってる。
"Zeki (1995) has proposed, however, that a direct geniculo-extrastriate path could account for the same phenomenon. That proposal is particularly attractive given the robust innervation of the foveal representation of V2 and the likelihood that this region has been little explored in studies of V1 ablation or cooling because of its proximity to V1. Thus, a relatively rich innervation of V2, with its feed-forward projections to most other extrastriate areas, as well as a much poorer direct geniculocortical innervation of those other areas, could well give the K layers a central role in blindsight."
Blindsightはfoveaだけによらないこと、それからnhp studyでは多くの場合V1 lesionによってV2もablationされていること、などからして賛成できませんが、residual visionを支えるpathwayがM, P, Kのどれか、という視点は大変重要であると思います。
今日はこのへんで。
- / ツイートする
- / 投稿日: 2007年09月15日
- / カテゴリー: [上丘、FEFと眼球運動]
- / Edit(管理者用)
2007年09月13日
■ V1から上丘へ行く経路はなにをしているか
今回もサーベイとJCを兼ねて。上丘への視覚入力にはretinaから直接来るretinotectal (retinocollicular)な経路と大脳視覚皮質を介してくる経路があります。V1からも来るし、V2からも来る。LGNからは来ないみたいだけど。Schillerの論文でJ Neurophysiol. 1974 Jan;37(1):181-94. "Response characteristics of single cells in the monkey superior colliculus following ablation or cooling of visual cortex."というのがあります。nhpのV1をcoolingもしくはlesionして急性で上丘の視覚応答を記録すると、SGSの応答はcoolingまたはlesionによって影響を受けないけれども、SGIの視覚応答は全くなくなる、というものです。つまり、SGSはretinaからの直接入力によってdriveされるけれども、SGIはV1経由の信号がないとvisual responseを引き起こすことができないのです。解剖学的にはV1のlayer5,6からSCへの投射があることが知られていますし、LIPやFEFを回ってくればいくらでも上丘へ行く経路はあります。ただ、麻酔下なんで、あんまり高次なところは回ってないんではないかと思うんですが。
ひきつづき彼らはJ Neurophysiol. 1976 Nov;39(6):1352-61. "Quantitative studies of single-cell properties in monkey striate cortex. IV. Corticotectal cells."にて、上丘(表面から0.5mmあたりと書いてあるので基本的にSGSからSoあたり)を刺激しながら、V1からunit記録をして、上丘の刺激によってantidromicに刺激されるunitを見つけます。こういうニューロンはV1のlayer 5,6にあって、antidromicの応答潜時は4.6ms。でもって、そういうニューロンの性質を調べると、complex cellで、orientation tuningはbroadで、direction selectivityがあって、spontaneous activityが高い。そういう意味ではSGSのニューロンの性質には似ていません。そこで彼らのdiscussionとしては、V1から上丘へ来る入力は「SGSの応答特性を作っている」というよりは、「SGS->SGIの情報の伝達をゲートしている」のではないか、と言っています。これは重要かつ面白い指摘です。
さて、このあとで彼らはJ Neurophysiol. 1979 Jul;42(4):1124-33. "Composition of geniculostriate input ot superior colliculus of the rhesus monkey."を発表します。これは上丘SGIからunit記録して、LGNのmagnolayerをlidocaineでblockすると応答が消えるけど、parvo layerをblockしても応答が消えない。どっちをblockしてもSGSの応答は消えない、という論文です。彦坂先生の仕事以降だとmuscimolになるのですが、この時代は局麻でblockしているのでpassing fiberもblockしてしまうという問題があります。ただ、SGSの応答は消えない、というのがcontrolで効いているので、だいたい妥当なのではないかと思うのですが。というわけで上丘はV1から受けている入力はmagno系だけだ、というわけです。色の情報とかはもらってない。
さて、この論文の意義を説明するためには視覚情報がRGC->LGN->V1->V2と進む情報伝達にはM,P,K経路の三つのチャネルがあるということの説明が必要です。図を作っておきました。
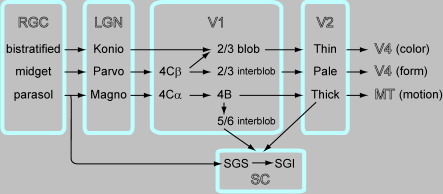
Schillerのだとcatの論文を引いて、X-type, Y-type, W-typeって話になるんでそこは翻訳しないといけないんですが、最新のレビューをもとにしてこれまでの知見を再構成してみました。図の元ネタはAnnu Rev Neurosci. 2005;28:303-26. "The circuitry of V1 and V2: integration of color, form, and motion." Sincich LC, Horton JC.および "The M, P, and K Pathways of the Primate Visual System" Ehud Kaplan in The Visual Neurosciences Ch.30です。図の表現上、一番上がkonio、真ん中がparvo、一番下をmagnoとしてあります。それぞれがcolor, form, motionのべつべつのチャネルとなってventral pathway, dorsal pathwayに行っていることはよく知られたことです。
それにさらに上丘への投射を組み込んでみました。LGNのparvocellular layerに終止しているRGCのX-type cell (midget cell)は上丘へは投射していません。LGNのmagnocellular layerに終止しているRGCのY-type cell (parasol cell)は上丘へ投射しています。あと、catではRGCにW-cellというのがあって、上丘に選択的に投射しているという話があります。(Klaus-Peter HoffmannによるBrain Res. 1972 Aug 25;43(2):610-6. "Very slow-conducting ganglion cells in the cat's retina: a major, new functional type?"およびJ Neurophysiol. 1973 May;36(3):409-24. "Conduction velocity in pathways from retina to superior colliculus in the cat: a correlation with receptive-field properties.") あと、conduction velocityが非常に遅い8m/secのはこのW-cellです。
ちょっと話が細かくなるのでここで改パラグラフ。CatのW cellがnhpになにに対応するかですが、koniocellularと考えた方がよいようです。Shapleyの80年代のTINSとかではそんな書き方をしています。そうすると、hnpではW-cellに対応するものは上丘へは投射していないと考えて方が良さそうです。ただ、Visual neurosciencesにもありましたが、W-cell、konioてのはどちらかというとmagno系、parvo系以外を全部入れたヘテロな集団と考えた方がよいので、これからの研究の進展によって話が変わってくる可能性はあります。なお、Schiller 1979では、W-like cellという言い方はintroでしているけど、それ以降ではまったく言及していません。また、それ以降の論文をしらべてみるとnhpでもW-like cellと表現しているのがあって、PNAS 2007 "Geniculocortical relay of blue-off signals in the primate visual system"とかではW-like = blue-off = koniocellularと書いてます。上丘にS coneからの入力が行っていないことはS cone isolating stimulusによる刺激による注意の効果の論文(Current biology)などから示唆されていますが、実験的には証明されていないようです。SGSからvisual response記録して、L-, M-, S-cone isolating stimulusで刺激するという実験をしたらいいと思います。きれいにS coneだけinsensitiveであることが示せたらJNSあたりいけるんではないでしょうか。
話を戻します。視覚皮質から上丘への入力はいくつかありますが、細かいことがわかっているのはV1,V2からのものです。
上丘にretrograde tracerを入れてV1で染まったニューロンの位置と、V1をCO染色してCOバンドとの位置とを比較したのがVis Neurosci. 1996 "The distribution of corticotectal projection neurons correlates with the interblob compartment in macaque striate cortex." Lia B, Olavarria JF.です。Layer 5/6のinterblobのニューロンが上丘へ投射しています。残念ながらinjectionは限局してないので、terminalはSGSなのかSGIなのかは不明。それ以前にもW. FriesがEBR 1985でlayer 5/6のMynert cellで上丘とMTの両方に投射しているものがあることを示しています(retrograde tracerによるdouble labellingを使用)。
同じグループが同様にしてV2で染まったニューロンの位置と、V2をCO染色してCOバンドとの位置とを比較したのがJ Comp Neurol. 1997 "Distribution of neurons projecting to the superior colliculus correlates with thick cytochrome oxidase stripes in macaque visual area V2." Abel PL, O'Brien BJ, Lia B, Olavarria JF.です。V2のthick band、つまりmagno系からおもに入力が上丘に来ています。
以上をまとめると図のようになるのでは、というわけです。
補足。ちなみに近年はganglion cellのレベルではなくて、coneのレベルでL+M-Sとかいったやり方で機能を描写するので昔の書き方を翻訳しないと使えません。このへんに関する最新の成果がEdward M. CallawayのNature 2003 "Parallel colour-opponent pathways to primary visual cortex"です。これを踏まえてconeからRGCまでの変換を書くと、parasol cellはcone全部の信号を足していて、L+M+Sで色に関わらない明るさをコードしていて、midget cellではL-Mでred/greenのシグナルを作っていて、bistratified cellではS-(L+M)でblue-onの受容野を持っています。そこでblue-offのニューロンを見つけてそれがV1のlayer 4Aに入力しているというのを見つけたのがNature 2003の仕事でした。これは図に組み込んでないけど、bistratifiedがblue-on cellでlayer 3Bあたりにメインで行ってて、それとはべつにRGCにblue-off cellがあって、これが4Aに終止していて、この二つを合わせてblue-on,offに関わるkoniocellular系を作っている、というのが最新バージョンではないでしょうか。
意外に充実してきたので今度のセミナーはこれにするかも。ちと専門的すぎるけど。
- / ツイートする
- / 投稿日: 2007年09月13日
- / カテゴリー: [上丘、FEFと眼球運動]
- / Edit(管理者用)
2007年09月05日
■ 上丘の細胞構築
もうすぐ神経科学大会ですね。私は一日目の午後に口演です。眼球運動のセッションの一番最後で、視覚1の直前。ちょうどいい位置に入れてもらいました。ぜひ見に来てください。
毎年7月末にあるトレーニングコースではラットの上丘、視覚皮質、バレル皮質などから急性で記録をしてあとでhistologyで記録部位を同定するということをやっております。わたしはだいたい電気生理の解析とhistologyの解析の担当をこの3年くらいやってます。上丘の細胞構築(cytoarchitecture)をきっちり把握しておく必要がありますので、元ネタにあたっておいたものをまとめておきます。
まず、上丘とはなんぞや、から始めてると時間がありませんが、上丘は中脳にあります。つまり、視覚皮質よりも腹側(ventral)で、視床よりも尾側(caudal)です。上丘は大きく分けて、retinaからoptic nerveを介して視覚入力が入ってくる入力層と、脳幹のサッケードジェネレーターに眼球運動の指令を送る出力層とに分かれます。おおまかには入力するoptic nerveが入ってくるSOと、メインの入力層であるSGSと、メインの出力層であるSGIとがわかれば充分です。
教科書的な図はありませんが、PNAS 2007 "Identity of a pathway for saccadic suppression"がopen accessなので、これの図を見てもらえると良いのではないでしょうか。Fig.3を見るとSGS,SO,SGIの順に並んでいることがわかるかと思います。さらにFig.2をみてもらうと上丘はさらに6層に分かれることがわかります。いちばん表面にある薄い層であるSZを入れると全部で7層構造です。ちょっと大脳皮質とは細胞構築が違う。ただ、SO=4層、SGIの大きい出力ニューロン=5層のpyramidal neuronくらいに当てはめてイメージすることはできるかもしれません。
解剖学の論文ってのは細胞構築、つまりニッスル染色でみた、細胞(主にニューロン)の大きさや密度などが層ごとにどのようなパターンを示しているか、の記述というのがずらずら並んでたりするので、それを読んで顕微鏡で標本を見てどこがどの層なのか「目を慣らす」という作業が必要となってきます。
Ratの上丘でのこのような細胞構築の分類の元となった論文は以下のJCN 1981です。これはさらにcatでの細胞構築JCN 1974 Kanaseki and Spragueを下敷きにしています。
J Comp Neurol. 1981 Sep 10;201(2):221-42. "Trigeminal projections to the superior colliculus of the rat." Killackey HP, Erzurumlu RS. PMID: 7287928
論文から抜き書きで7層についての記述を。
-
SZ: the stratum zonale
"a thin, loosely patched layer, which contains small neurons" -
SGS: the stratum griseum superficiale
"densely packed with cellular elements and is relatively thick" -
SO: the stratum opticum
"primary composed of fibrous elements with some scattered cells" -
SGI: the stratum griseum intermidiale
"band of large neurons readily detectable in Nissl preparations"
"densely packed with cells and is thick, like the stratum griseum superficiale" -
SAI: the stratum album intermediale
"characterized by patched of fiber bundles intermixed with a few cellular elements" -
SGP: the stratum griseum profundum
"has a layer of large neurons at its dorsal border" -
SAP: the stratum album profundum
"a fibrous layer that separates the superior colliculus from the periaqueducal dray"
細胞構築を決めてやるために必要なことは基本的にはこのような記述がすべてなのです。もちろん、immunohistochemistryとかでどこかの層がなんか特異なマーカー(CaMKIIだとかIh channelだとかcalcium binding proteinだとか)を持っているかを目印にしたりとか、トレーサーを注入して、投射関係などからさらにくわしく調べられ、consistencyの確認や、さらなる層の細分化が起こるわけです。たとえばの例として、LGNがmagnocellularとparvocellularに二分されていて、層のあいだの隙間はinterlaminarなんて言われていたところをCaMKII α-unitで染めることによってkoniocellularの層が確立する、ということもありました(Science 1994 Hendry and Yoshioka)。Koniocellularという言葉自体はその前からあったんですが。
ちなみに、ネットで探してみたらヒトでの上丘の細胞構築については「中脳蓋-上丘を主として-」を見つけました。
- / ツイートする
- / 投稿日: 2007年09月05日
- / カテゴリー: [上丘、FEFと眼球運動]
- / Edit(管理者用)
2006年01月12日
■ antisaccadeを使った自発的眼球運動の研究
060110のエントリでわたしは以下のように書きましたが、
Munozが注目する理由はよくわかります。行動開始がvoluntaryであるか否か、という問題に関わるのではないか、ということでしょう。Monoz自身はprosaccadeをreflexive saccadeとして、antisaccadeをvoluntary saccadeとして扱ってこの問題に取り組んでいます(Nature Reviews Neuroscience 04 "LOOK AWAY: THE ANTI-SACCADE TASK AND THE VOLUNTARY CONTROL OF EYE MOVEMENT")が、今回の論文はそれと問題意識を共有しているように思います。
コメントをしてくださったvikingさんのところでオリジナル論文の方(Nature Neuroscience '02 "Human fMRI evidence for the neural correlates of preparatory set")が解説されてます。こうやって話題がつながってゆくのはすばらしいことです。
以前に読んだときは、Gap periodが長すぎて、express saccadeが出るような、いわゆるgap paradigm (gap = 200ms)とは違うよなあ、とか思いましたが、このパラダイムも"when to saccade"に関わってくるので重要です。あと、なぜFEFであってIPSでないのか、というあたりは、parietalでやっているらしい目標設定と、FEFでやっているらしい運動準備やら行動選択やらの違いが反映しているようなので、そういう目で精読してみよう、と思いました。
- / ツイートする
- / 投稿日: 2006年01月12日
- / カテゴリー: [上丘、FEFと眼球運動]
- / Edit(管理者用)
2005年11月01日
■ Pursuitのエラーは視覚由来
Nature 9/15。"A sensory source for motor variation" Osborne, Lisberger and Bialek。F1000で挙げられていたのですこし読んでみました。
Pursuit=追従眼球運動、では動いている視覚刺激を追いかけて目を動かすのですが、この運動を正確に行うには目からの入力でたえずフィードバックをかけながら眼球運動の軌道を修正してゆく必要があります。そういうわけで、sensorimotor coordinationを研究するには良い系なわけです。んでもって、それでもこういう運動をするときには視覚刺激の位置と実際の目の位置とにはズレが起こります。このズレがなにに起因するかをモデルを作って計算した、というのがこの論文。視覚刺激自体の位置の推定によるズレと眼球運動のプログラムの段階でのノイズに起因するズレとがあるわけですが、ほとんどのズレは視覚刺激自体の位置の推定によるズレであったと。
F1000ではWilliam Kristanがコメント付けていて、ほかの課題でも応用可能であろうと。というわけでもうすこし詳しいことを読んでみる必要あり。Carpernterの仕事関連とか、さいきんのJNS 10/26 "The temporal impulse response underlying saccadic decisions"とかとも併せて。
Lisbergerはpursuit関連の仕事をやり続けてきた人。PubMedでさかのぼってみると、どうやらFuchs AFの弟子らしい。Bialekは"Spike"の著者。同じ著者グループでJNSにMTニューロンの記録も出してる。今回の論文は、そのついででこれまでのpursuitの行動データを解析してみたらこんなことがわかりました、ということなのでしょう。きっと次は同じメンツでMSTニューロンの記録で今回の解析を適応、ということでしょう。- / ツイートする
- / 投稿日: 2005年11月01日
- / カテゴリー: [上丘、FEFと眼球運動]
- / Edit(管理者用)
# V1の人
ごぶさたしております。
> PubMedでさかのぼってみると、どうやらFuchs AFの弟子らしい。
こんなのご存じですか?↓
http://neurotree.org/neurotree/
Lisbergerだったら
http://neurotree.org/neurotree/tree.php3?userid=guest&sessionid=my78IKkmW1ix6&pid=93
まだまだデータベースに偏りがありますが、「AさんとBさんは何親等(?)なのだろうか」とかいうのも調べられてなかなか面白いです。
# pooneilどうもお久しぶりです。neurotree見ました。おもしろいですね。思わずうちのボスにも教えてしまったり。これをネタにエントリを作ってみようと思います。
それから、所属変わったようですね。スケジュールがあえば、Washinton DCで会いましょう。ではまた。
2005年08月23日
■ JNP 9月号
"Chronometry of Visual Responses in Frontal Eye Field, Supplementary Eye Field, and Anterior Cingulate Cortex" Pierre Pouget, Erik E. Emeric, Veit Stuphorn, Kate Reis and Jeffrey D. Schall
Visual Response latencyをFEF,SEF,ACCで比較した。以前のJNP '98 "Signal Timing Across the Macaque Visual System"の続編みたいなもん。このJNP '98はLGN-V1-V2-V3-V4-MT-MST-FEFでvisual Response latencyを比較した、というもので、データは寄せ集めでしかないのだけれど、こうやって比較したものはほかにないので貴重な資料。Response latencyは視覚刺激が最適なものであるかどうかに大きく依存する、とかいろいろつっこみどころはあるにせよ。FEFのほうがV4よりもlatencyが早い、とか重要なことが書いてある。
前回のデータと照らし合わせてみると、SEFはFEFよりは遅くて、V2と同じくらい。ACCはSEFよりもさらに遅くて、V4と同じか遅いくらい、というかんじらしい。
- / ツイートする
- / 投稿日: 2005年08月23日
- / カテゴリー: [上丘、FEFと眼球運動]
- / Edit(管理者用)
2005年06月28日
■ Double step task関連
なんかやけに視床づいていますが、Sommer and WurtzのDouble-step task関連についてまとめておこうと思います。
というのも、Brain 2005 5月号に視床にlesionのある患者さんでdouble step taskをやった、というものがあるのを知ったからです。
"The role of the human thalamus in processing corollary discharge" C. Bellebaum, I. Daum, B. Koch, M. Schwarz and K.-P. Hoffmann
で、話は簡単でなくて、MD核に損傷がある場合にはその同側でsecond saccadeに影響が出る(n=1)のに対して、VL核の損傷の場合にその対側でsecond saccadeに影響が出る(n=3)、というのです。
Sommer and WurtzのときはMD核を抑制するとその対側でsecond saccadeに影響が出るわけですから、どうも話が合わないのです。
Sommer and Wurtzは昨年JNPにfollow-up paperも出したことですし、ここらでまとめてみよう、というわけです。
DuhamelのBrain 1992についても併せてまとめてみましょう。(しかしそれにしてもDuhamelの1992年というのはすごい年で、いっぽうでScienceにsingle-unitでのLIPのremappingについての論文("The updating of the representation of visual space in parietal cortex by intended eye movements")をファーストオーサーとして書き、またいっぽうでpatient studyでBrainにファーストオーサーとしてcorollary dischargeの寄与に関する論文("Saccadic dysmetria in a patient with a right frontoparietal lesion. The importance of corollary discharge for accurate spatial behaviour")を書いているわけです。余談ですが。)
それから、田中真樹さん@北大のJNP 2003 "Contribution of Signals Downstream From Adaptation to Saccade Programming"もメモメモ。
というわけでそのうちつづきます。
- / ツイートする
- / 投稿日: 2005年06月28日
- / カテゴリー: [上丘、FEFと眼球運動]
- / Edit(管理者用)
2004年12月22日
■ Neuron 12/16
Julio C. Martinez-Trujillo, W. Pieter Medendorp, Hongying Wang and J. Douglas Crawford "Frames of Reference for Eye-Head Gaze Commands in Primate Supplementary Eye Fields." Volume 44, Issue 6, 16 December 2004, Pages 1057-1066
SEFがどういう座標系をコードしているか、という問題についてですが、眼球運動と首の動きと両方の自由度のある条件でSEFを微小電気刺激しています。
可能性としてはbody-centeredか、head-centeredか、eye-centeredか、ということがありうるのですが、結論としては、いろんな座標のがあった、だからSEFはこれらを変換している場所なのだろう、というきれいな結果が分離しなかったときのための常套手段のまとめ方をしています。あんまり印象よくない。きれいに分かれないなら分かれないなりに変換の過程を追うような解析をしているかどうかでしょうな。
- / ツイートする
- / 投稿日: 2004年12月22日
- / カテゴリー: [上丘、FEFと眼球運動]
- / Edit(管理者用)
2004年07月08日
■ Nature Neuroscience 7月号
"Deficits in saccade target selection after inactivation of superior colliculus." Edward L Keller @ The Smith-Kettlewell Eye Research Institute。
上丘はサッケードのinitiationだけではなくていろんな認知機能(Horwitz and Newsomeによる"target selection"やPeter Thierによる"covert shift of attention"など)に関わる情報を持っていることが主にsingle-unit recordingからわかっています。しかしそれらの研究に必ず付きまとう問題はそのような機能は上丘が担っているのではなくて、単に大脳皮質で表象されている情報が伝わってきているだけではないのか、といわれてしまう難点があることです。そのためには上丘のサッケードの機能を阻害せずに上丘での"target selection"などの機能だけを選択的に阻害できることを示すことが必要です。これは主にmuscimolを局所的に注入することで行われています。今回の論文もサッケード自体には影響がなくてtarget selectionだけが阻害されるような結果を提示しています。そういえばPeter Thierも玉川のシンポジウムで話をしたときには上丘のlesionやmuscimolによるreversible inactivationが重要である、というような言い方をしてました。あれの場合はサッケードのターゲットとattentionのcueの位置が離れていることから、両者を分離してinactivationすることが可能です。そのうち論文になって出てくるかもしんない。
- / ツイートする
- / 投稿日: 2004年07月08日
- / カテゴリー: [上丘、FEFと眼球運動]
- / Edit(管理者用)
2004年04月16日
■ Sommer and Wurtz関連
このあいだのセミナーで関連論文が採り上げられてたので整理しておく。
Science '02 "A Pathway in Primate Brain for Internal Monitoring of Movements."で、上丘から視床のMDを通ってFEFへとcorollary dischargeが帰ってきていることを示唆した。何よりよいのはFEF->SCの向きはintactなままでSC->FEFの向きだけMDへmuscimolを注入することで止めることができるために、主張が強くできるところだ。SCやFEFへの注入ではそうはいかない。これが以前さかんに調べた内部モデル関連の話のサッケード系のアナログだといえる。これがsequentialな運動のplannningに使われていることは示されたわけだが、これが視覚像自体をmodifyしている可能性は残る(presaccadic remapping的なことが起こっているかどうか)、とScience論文の最後に書かれている。で、これの続報というかfull paper化したものがJNP '04の連報の
"What the Brain Stem Tells the Frontal Cortex. I. Oculomotor Signals Sent From Superior Colliculus to Frontal Eye Field Via Mediodorsal Thalamus."
"What the Brain Stem Tells the Frontal Cortex. II. Role of the SC-MD-FEF Pathway in Corollary Discharge."
として出てきた。
連報IでMDから記録して、上丘からorthdromicに刺激され、FEFからantidromicに刺激されるところを同定。
連報IIで同定したMDにムシモルを入れて、double step taskがどう影響を受けるかを調べた。
このネタで書いたレビューが
Current Opinion in Neurobiology '03 "The role of the thalamus in motor control."
および
"Identifying corollary discharges for movement in the primate brain."
前者では視床を通ってmotor関連領域からcortexへ帰る三種類の経路について議論している。(1) 小脳 -> VPL/VL ->M1/PM (2) globus pallidum/SNr -> VL/VA -> SMA (3) 上丘 -> MDmf -> FEF。で、彼らの実験では(3)の経路について検討した。
で、上丘からFEFへ帰ってゆくのが順キネマティクスで小脳からFEF(?)または上丘へ帰ってくるのが順ダイナミクスであるとすると尤もらしい。これこそCOEシンポで川人先生と話をしたときに出てきた、眼球運動系でのダイナミクス系の回路への解答なんだと思う。また、逆モデルと順モデルとがカップルせずにそれぞれが別経路を通ってるかもしれない、という意味で重要と思われる。
Sommer自身は前者の論文のConclusionで、(1)がvisually-guided eye movementに、(2)がinternally generated eye movementに関わっているかどうか(arm movementで示されているように)検討すべきであるとしてまとめている。
WolpertはCurrent biology '02 "Cerebral Carbon Copies."でScienceについてコメントし、(3)の経路について、サッケード中の眼球運動の位置を予測するのに使われている、としている。
後者は入手できなかったので読んでない。
ほかにこの著者の論文へのコメンタリとして、
"Seeing and Acting at the Same Time: Challenges for Brain (and) Research."
"Neurophysiology: Cerebral Carbon Copies."
がある。
- / ツイートする
- / 投稿日: 2004年04月16日
- / カテゴリー: [上丘、FEFと眼球運動]
- / Edit(管理者用)
2004年02月07日
■ JNPつづき
"Comparison of Memory- and Visually Guided Saccades Using Event-Related fMRI."
この論文の結果(2/5のところを参照)をsingle-unit recordingの結果と比較してみるのは面白いことであろう。Visually-guidedでdistractorとして出てきた刺激とmemory-guidedでcueとして出てきた刺激との間ではdistractorのほうが頭頂連合野やFEFでより強い反応をしているのだ(activationのあるvoxelは小さいけれど)。まったく同じ条件のものはないけれど(とくにreceptive fieldの概念を考慮しなければ意味がないのだが)似たことを扱っているsingle-unitでのLIPニューロンからの記録の論文にGoldbergのJNP '00 "Response of Neurons in the Lateral Intraparietal Area to a Distractor Flashed During the Delay Period of a Memory-Guided Saccade."やGlimcherのJNP '97 "Responses of Intraparietal Neurons to Saccadic Targets and Visual Distractors."がある。
Goldbergのはmemory-guidedのdelay中にdistractorが出るというものだが、targetと同じ位置に出るdistractorよりも違う位置に出るdistractorの反応のほうが大きい。あんまり関係ないがとにかくtask-irrelevantでもattentionをかけているところ以外にabruptに現れると反応は大きい。
Glimcherのほうは二つのpotential targetが提示されてからあとでcueによってどちらがtargetでどちらがdistractorかが決まる。当然potential targetが提示された時の応答は同じわけだが、cueによってどっちがtargetか決まると、tagetに大きい反応をして、distractorへの応答は抑制される。
というわけで答えは出ないのだが、刺激のabruptさ(Goldberg)という意味では今回の論文ではcueとdistractorでの反応は同じと予測できるが、goal-relatedであるかどうか(Glimcher)という意味ではcueのほうが反応が大きくなってしかるべきと思える。
- / ツイートする
- / 投稿日: 2004年02月07日
- / カテゴリー: [上丘、FEFと眼球運動]
- / Edit(管理者用)
2004年02月06日
■ JNPつづき
"Comparison of Memory- and Visually Guided Saccades Using Event-Related fMRI."
で、もう少し読んでみた。ブロックデザインでmemory-guidedとvisually guidedをやらせてそのコントラストを見ると、いろんなところでmemoey-guidedでのほうがactivationが高い。これはmemory-guidedでのcue、delay、saccadeの成分を全部足し合わせているからなので、続いてevent-relatedにしてcue、delay、saccadeのそれぞれの成分ごとにmemory-guidedとvisually guidedの比較をしている。このため、visually-guidedのタスクではmemory-guidedのcueの代わりにsaccade targetとは無関係な刺激(distractor)を出して二つのタスクでの条件の違いをなるたけ少なくしようとしている*1。しかしそれでもsaccadeをするときにtargetがあるかないかの違いは当然残る。この意味ではたいして工夫のない実験デザインであると言える。
*1:これが昨日書いた謎な、刺激への応答のコントラストだった。
- / ツイートする
- / 投稿日: 2004年02月06日
- / カテゴリー: [上丘、FEFと眼球運動]
- / Edit(管理者用)
2004年02月05日
■ JNP
"Comparison of Memory- and Visually Guided Saccades Using Event-Related fMRI."
GoodaleとEverling @ University of Western Ontario。
Humanのvisually-guided saccade (標的にサッケードする)とmemory-guided saccade (標的の出た位置を思い出してサッケードする)とでの脳活動部位をfMRIで調べたペーパー。前にはblockデザインでPETをやったものがあるが、fMRIで論文になったのはこれが最初らしい(SFN abstractが一つ引用されている)。Event-relatedでやっている。
で、かつてのPETスタディではPEF(nearly equals LIP)、FEF、SEFの活動はmemory-guidedのときのほうが高かった。今回の論文ではvisual responseに関してはfrontal(FEF含む)とparietal(right rost. IPS)では刺激(cueとtargetの比較?)による反応はvisually-guidedでやや高く、サッケード時の反応はright rost. IPSではmemory-guidedのとき高かった。
- / ツイートする
- / 投稿日: 2004年02月05日
- / カテゴリー: [上丘、FEFと眼球運動]
- / Edit(管理者用)
2004年01月05日
■ JNS 2003年1月号。 Journal club前の予習
"Controlled Movement Processing: Superior Colliculus Activity Associated with Countermanded Saccades"
Paré @ Queen's UniversityとHanes @ National Eye Institute。
HanesはJeff Schallとcountermanding task中のFEFのニューロン活動を記録してきた。今回は同じパラダイムで上丘からsingle unitを記録した。Hanesの出世作はScience '96
"Neural Control of Voluntary Movement Initiation"
で、今回の論文と直で関わるのはJNP '98。
"Role of Frontal Eye Fields in Countermanding Saccades: Visual, Movement, and Fixation Activity"
まずかんたんにcounermanding taskの説明:
- まずコントロールのタスク。動物はタスクの開始で注視点を見つめる。すると注視点が消え、サッケードの標的の点が現れる。こちらにサッケードすると報酬が貰える。
- いっぽう本命の課題では、注視点が消えて標的が現れたしばらくあと(25-275ms)に、ふたたび注視点が現れる。このとき動物は標的へサッケードせずに注視点を見つづけなければならない。しかし持ちこたえられずにどうしてもサッケードしてしまう試行がある割合で出る。この行動の結果と神経活動との相関を調べる、というのが基本的なアイデア。
で、JNS'03は同じ課題で上丘から記録しているわけだが、それでどういう新しいことが見出せたかが重要。FEFにある情報が上丘に行くのはあたりまえなので、FEFで見られるような神経活動が上丘で見られるのはあたりまえであり、それではJNSは通らない。JNPレベルでしょう。というわけで何か新しいものがあるはず。
要旨を読む限りだと、
- [サッケードをキャンセルするかしないかによる神経発火パターンの差が出る時間]のほうが[サッケードをキャンセルするかしないかがいちばん瀬戸際のときの時間]より10msだけ早い
- そしてこの10msという時間は上丘の発火から実際のサッケードまでにかかる時間よりも短い
- / ツイートする
- / 投稿日: 2004年01月05日
- / カテゴリー: [上丘、FEFと眼球運動]
- / Edit(管理者用)
2003年12月31日
■ Nature Neuroscience AOP祭り again (4)
"Neuron-specific contribution of the superior colliculus to overt and covert shifts of attention."
Peter Thier @ University of Tuebingen。
Rizzolattiのpremotor仮説というのがあって、「サッケード指令」(overt shift)と「目を動かさずに注意だけ向けること」(covert shift)とが同じメカニズムを共有している、というもので、Kustov and RobinsonのNatureでの上丘刺激とかShadlenのNature '01でのFEF刺激とかで支持されてきた。この論文はsingle-unit recording。
- / ツイートする
- / 投稿日: 2003年12月31日
- / カテゴリー: [上丘、FEFと眼球運動]
- / Edit(管理者用)
お勧めエントリ
- 細胞外電極はなにを見ているか(1) 20080727 (2) リニューアル版 20081107
- 総説 長期記憶の脳内メカニズム 20100909
- 駒場講義2013 「意識の科学的研究 - 盲視を起点に」20130626
- 駒場講義2012レジメ 意識と注意の脳内メカニズム(1) 注意 20121010 (2) 意識 20121011
- 視覚、注意、言語で3*2の背側、腹側経路説 20140119
- 脳科学辞典の項目書いた 「盲視」 20130407
- 脳科学辞典の項目書いた 「気づき」 20130228
- 脳科学辞典の項目書いた 「サリエンシー」 20121224
- 脳科学辞典の項目書いた 「マイクロサッケード」 20121227
- 盲視でおこる「なにかあるかんじ」 20110126
- DKL色空間についてまとめ 20090113
- 科学基礎論学会 秋の研究例会 ワークショップ「意識の神経科学と神経現象学」レジメ 20131102
- ギャラガー&ザハヴィ『現象学的な心』合評会レジメ 20130628
- Marrのrepresentationとprocessをベイトソン流に解釈する (1) 20100317 (2) 20100317
- 半側空間無視と同名半盲とは区別できるか?(1) 20080220 (2) 半側空間無視の原因部位は? 20080221
- MarrのVisionの最初と最後だけを読む 20071213