[カテゴリー別保管庫] Papers_unclassified
2007年11月16日
■ 脊髄損傷からの機能回復には、回復時期ごとに違った脳部位が関与する
わたしが所属している生理学研究所・認知行動発達研究部門からついにhigh-quality journal paperが出ました。
Science 16 November 2007: Vol. 318. no. 5853, pp. 1150 - 1155 "Time-dependent central compensatory mechanisms of finger dexterity after spinal cord injury" Yukio Nishimura, Hirotaka Onoe, Yosuke Morichika, Sergei Perfiliev, Hideo Tsukada, Tadashi Isa
やってることはなにかというと、脊髄のC4-C5 segmentをtransectした動物での機能回復時に大脳皮質のどの部分が関わっているかを調べたところ、回復初期には同側のM1が、回復後に安定した状態では対側のpremotor cortexが関わっているということを示した、というものです。
この実験では大脳皮質から直接脊髄まで投射している経路だけを選択的にtransectionしています。つまり、partial lesionです。Transectされた以外の経路として、大脳皮質から脳幹、延髄などでいったん終止してそこから脊髄へ投射する経路がありますが、これらは正常に保たれています。よって、transection直後には指先を使ってつまむ動作(棒の先に付いたイモのブロックをスリットからつまんで食べる)が出来なくなりますが、訓練によって約一ヶ月程度で術前の成績に近くなります。(なお、手を伸ばす動作reachingに関しては大きく影響を受けることはありません。)
そういうわけで機能回復時には残っている脊髄への投射経路を使っていることはわかるわけですが、そこへの大脳皮質からの指令がどう変わっているか、これを調べたのです。PETを使って、[rewardの餌をスリットからつまんで食べる条件]から[control条件(棒の先についたrewardの餌を受動的に食べる)]の差分をとっています。よって、graspingとreachingの両方のactivationが寄与してます。PETですので、event-relatedではなくて、block designです。
術前のコントロールでは使っている手の対側のM1が活動します。手術後一ヶ月ではそれに加えて同側のM1が活動しています。手術後三ヶ月では同側のM1の活動は消えて、対側のPMの活動が見えてきます。この時期にはつまむ動作の成績はほぼ正常になっていますが、対側のPMにmuscimolを注入して一時的にその活動を抑制させると、また成績が悪くなります。術前の対側のPMにmuscimolを注入してもこのような効果はありません。
というかんじでいくつか手法を組み合わせてかなり堅い結論を持ってきているのがわかるかと思います。すばらしい。ここまでの苦労を傍目から見ておりましたので非常に感慨深いです。
「損傷後の機能回復に脳はどのように寄与しているのかを明らかにする」という意味において私が現在やっている仕事は同じ目的を共有しています。つまり意識の問題と基礎医学への寄与とを両方やってしまおう、というプロジェクトなのです。さあ、私もこれにつづいて行こう、というのが結論なのですが。
今回のScience論文の仕事はCRESTによるコラボレーションの産物です。じっさい、所属を見てもらうとわかりますが、生理研、浜松ホトニクス、理研、イエテボリ大学というかんじで並んでいます。
ちなみに先週のScienceでは理研の田中啓治先生のラボがOxfordのBuckleyと行ったWCST関連の仕事が掲載されています。日本のnhp studyがんばってます。こちらの仕事もコラボレーションの産物ですね。
Science 9 November 2007: Vol. 318. no. 5852, pp. 987 - 990 "Mnemonic Function of the Dorsolateral Prefrontal Cortex in Conflict-Induced Behavioral Adjustment" Farshad A. Mansouri, Mark J. Buckley, Keiji Tanaka
そういえば5年前くらいだったかBuckleyが日本に来ていくつかラボ回って話をしに来てましたが(前ラボにも来た)、そのあたりからこのプロジェクトが進んでたのでしょうね。
- / ツイートする
- / 投稿日: 2007年11月16日
- / カテゴリー: [Papers_unclassified]
- / Edit(管理者用)
2007年05月02日
■ 痛そうな写真
はてブで捕捉したasahi.com> サイエンス> 脳内メカニズム解明 写真見て「痛い」 群馬大院教授ら経由で、Cerebral Cortexに載ってる"Inner Experience of Pain: Imagination of Pain While Viewing Images Showing Painful Events Forms Subjective Pain Representation in Human Brain"というのを発見しました。柿木先生が著者に入ってます。
んで、痛みを想像したときに活動する脳部位をfMRIで見たって話なんだけど、その視覚刺激として使っている「痛そうな写真」ってのがFig.1(残念ながらopen accessではない)に載っていて、これがとてもいい感じ。この実験のために作った合成写真なんだろうけど、腕に刺さった注射針が突き抜けてるし! 50mlのシリンジに18ゲージのぶっとい注射針付けて腕に刺してて、その部分が赤くなっているとか。かなり最高です。
コントロールの刺激としては、IAPS (International Affective Picture System)という刺激セットがあって、これの中からvalence and arousalのレーティングが高かったものをemotion of fearの条件として使っているとのこと。それでもって、imagery of pain - emotion of fearでemotionalな反応とはべつの場所でもimagery of painのactivationが出てるのを見てます。Imagery of painにspecificなのがinsulaとかで、emotion of fearではamygdalaで、両方で出るのがACCという話。
心の痛みのimaging、というのでは以前(20040730)に恋人が痛い目に遭っているときに活動する脳領域という話がありました(Science 2004 "Empathy for Pain Involves the Affective but not Sensory Components of Pain")。あれもinsulaとACCでしたね。
{kind=link}
- / ツイートする
- / 投稿日: 2007年05月02日
- / カテゴリー: [Papers_unclassified]
- / Edit(管理者用)
2007年03月18日
■ ショウジョウバエのattention
"Attention-Like Processes in Drosophila Require Short-Term Memory Genes"
ショウジョウバエでもattention-likeなLFP応答があって、dunceのmutantではその応答がなくなるし、行動の指標でもattention-likeなものがなくなるとのこと。ただ、ここでのattentionってのはnovelty detectionみたいなもののことなので、短期記憶への影響と関連があるのはほとんど自明なようにも思えるのですが。
この分野もう少し押さえておきたいのだけれどなかなか手が伸びない。Referenceから追ってみると、ショウジョウバエでのchoice behaviorなんてのもありますね。Science 2001 "Choice Behavior of Drosophila Facing Contradictory Visual Cues"。中公新書から出ている水波誠氏の「昆虫-驚異の微少脳」も面白かったです。
無脊椎動物での認知機能を神経活動と絡めて明らかにするという方向性は、よりシンプルな系でニューロンのネットワークがどのようにして認知的な活動を支えているのかというのを明らかにするという点で興味があります。以前もヒルの意志決定の論文を読んだことがありましたが、このへんがいろいろ進んでいるのでもっと知りたいです。以前に研究会の企画をしていて、国内の関連する研究者の方を探していたことがありました。なんかの機会に再チャレンジしてみたいと考えているところです。
- / ツイートする
- / 投稿日: 2007年03月18日
- / カテゴリー: [Papers_unclassified]
- / Edit(管理者用)
2006年08月28日
■ LTPと学習
前回のエントリのBear論文についてryoさんがコメントしてます。Inhibitory avoidance learningという課題が鍵であるらしい。
私自身はEdvard I. Moserのsaturation論文でかたがついてたような気がしてたけど、そうでもなかったようです(20040731のエントリに短いコメント有り)。Moser論文のPerspectivesを当のT. V. P. Blissが書いていて、
"Striking though the results of Moser et al. are, skeptics may remain unconvinced. Almost half the tetanized animals demonstrated residual LTP; in fact, the average residual potentiation in this group was no different from that seen in previously untetanized control animals."
なんて書かれているところを見ると、結果としてはそんなに強くなかったらしい。
あと、今回はじめて気がついたけど、Science '93 "Association between brain temperature and dentate field potentials in exploring and swimming rats."(water mazeとかをやらせたratのDGのfEPSPが上がるのは、体温上昇のせいである、としたやつ)はMoserの論文だったのですね。
つれづれと。
- / ツイートする
- / 投稿日: 2006年08月28日
- / カテゴリー: [Papers_unclassified]
- / Edit(管理者用)
2006年08月17日
■ In vivoでawake behavingでwhole-cell recording
Neuron 8/17号。Whole-Cell Recordings in Freely Moving Rats
この論文がどうかではないのだけれど、intraでもwhole-cellでも、awakeのbehavingでsystemと組み合わせると、subthresholdのmembrane potentialのfractuationがどういう情報を持っているか、とかマニアックめな方向へ行ってしまうのがなんか歯がゆいなあと思う。なんかもっとすごいことが出来そうに思うので。
Referenceからまとめ:
- Nature Neuroscience 4, 184 - 193 (2001) In vivo intracellular recording and perturbation of persistent activity in a neural integrator D. W. Tank
- Nature 427, 704-710 (19 February 2004) Whisker movements evoked by stimulation of single pyramidal cells in rat motor cortex Bert Sakmann
- Pflügers Archiv 2004 In vivo, low-resistance, whole-cell recordings from neurons in the anaesthetized and awake mammalian brain Bert Sakmann
- Neuron, Volume 27, Issue 3, Pages 461-468 Active Stabilization of Electrodes for Intracellular Recording in Awake Behaving Animals M. Fee
- / ツイートする
- / 投稿日: 2006年08月17日
- / カテゴリー: [Papers_unclassified]
- / Edit(管理者用)
2006年07月14日
■ 和音の幾何学、つづき
つづいてます。
Perspectivesの"Exploring Musical Space"のほうを読んでいたら、'tonnetz'自体がすごいおもしろいものであることがわかってきました。自作したのを使って説明してみます。(tonnetzでググってみたら、日本語で引っかかってくるサイトがみつからなかったんですが、日本語だとなんて言うんですかね。)
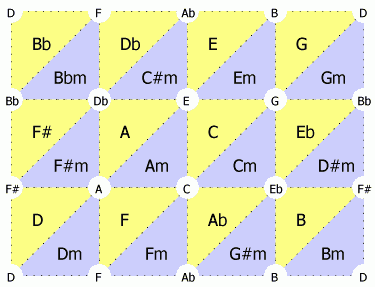
Fig. 1
そもそもtonnetzとはなにか、というと、音程間の関係を二次元上に表現した格子状のダイアグラムのことです。(Wikipediaのエントリから超訳。)
Fig.1はPerspectivesにあった図を説明のためにいくつか改変して作った図です。三角形が貼り合わされていて、中心にあるのが3和音を、頂点がその3和音を構成する音程を示しています。Fig.2aに例がありますが、CコードはC音(ド)、E音(ミ)、G音(ソ)からなるわけです。ふたたびFig.1に戻ってもらうと、黄色い三角形はメジャーコード(長調の3和音)を、水色の三角形はマイナーコード(短調の3和音)を示しています。共通の音を持つコードが貼り合わされています。あとは説明としてはこんなかんじ: (なお、説明で和音Cと単音Cとを混同しないよう、和音は「Cコード」、単音は「点C」とか「C音」と呼んでますので。)
- それぞれの三角形の頂点から真上に行くと、長3度(=半音4個ぶん)高い音になります(たとえば、点Cの上はE)。
- それぞれの三角形の頂点から右に行くと、短3度(=半音3個ぶん)高い音になります(たとえば、点Cの右はEb)。
- それぞれの三角形の頂点から右上に行くと、完全5度(=半音7個ぶん)高い音になります(たとえば、点Cの右上は点G)。
- それから、いちばん上の辺といちばん下の辺はつながってます。たとえば、点Cの上は点Eで、その上は点Abで、その上は点C、というかんじで一周します。
- それから、いちばん右の辺といちばん左の辺はつながってます。たとえば、点Cの右は点Ebで、その右は点F#で、その右は点Aで、その右は点C、というかんじで一周します。
- つまり、全体としてはドーナツ型(トーラス構造)をしているわけです。
- このトーラス構造を右上にずっと進んでいってみましょう。たとえば、点Cから右上に進んでゆくと、C-G-D-A-E-B-F#-Db-Ab-Eb-Bb-F-Cと重なることなく一周して戻ってきました。これが5度圏(circle of fifths)です。
- こんどは和音に注目して右上に進んでみましょう。Dmコードの右上はFコードで、その右上はAmコードで、その右上はCコードで、その右上はEmコードで、その右上はGコードです。つまり、Cメジャーダイアトニックコード(ドレミファソラシドだけからなる和音)となっているわけです。
- じゃあ、その先はどうよっていうと、Gコードの右上はBmコードでF#の音が入ってしまうし、Dmコードの左下はBbコードでBbの音が入ってしまう。つまり、Fig.2bのような事態になるわけです。5度圏で12音を一周することを考えたら、ドレミファソラシド以外の音も通るに決まっているから、そりゃあたりまえなわけです。
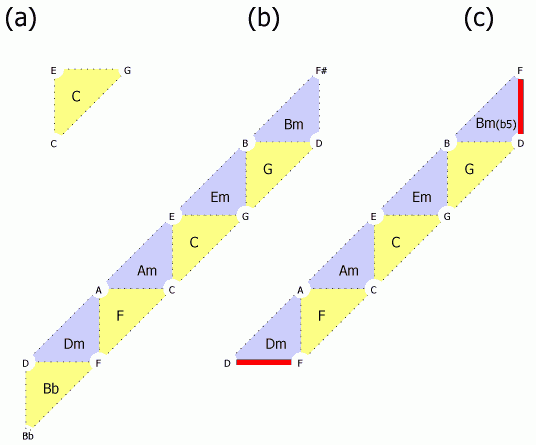
Fig. 2
とか思いながらFig.2bの絵を見てて気付いたのですが、BmコードのD音-F#音-B音の代わりに、D音-F音-B音という和音を持ってくると、Fig.2cにあるように、上下の赤線部分を繋げてやることが出来ます。これはBm(b5)コードもしくはBm-5コードと呼ばれる、Cのメジャーダイアトニックのコードのひとつです(*注)。Bm(b5)コードは、BmコードのF#音をF音に変えたものとも言えるし、BbコードのBb音をBに変えたものとも捉えることが出来るでしょう。つまり、Bm(b5)はBmコードとBbコードとの中間型みたいなものなんですな(これじたい今考えてみてはじめて気付いた)。ともあれ、これを使うと、右上のGコードと左下のDmコードとをつなぐことができるわけです。そして、Bm(b5)コードを加えたこの7つでCメジャーダイアトニックコードの全部が出そろいます。
つまり、トーラスを5度圏で一周する代わりに、横着して7つの3和音で一周してつなげてしまったのがメジャーダイアトニックコードなのですな! おもしろくなってきた。なんか矛盾を無理矢理つじつま併せて、メジャースケールというひとつの閉じた系として脳が捉えている様子がうかがえます。色の彩度が波長によって青から緑、赤まで一次元的に並んで終わりというのではなくて、なぜか紫を経てまた青に戻るという閉じた構造をとること(色環)と並べて考えてみるとおもしろい。(脳科学っぽくなってきた!) これはこの種の表象が持つ性質なんではないかと思うのです。いま、「表象」という言葉を使うことによって、言語を使った表象が持つ閉域形成の性質(指し示すものと指し示されなかったものとの二分法によって外部をなくしてしまう性質)と、色や音程が空間的に表現されるという意味での表象が持つ閉域形成とを結びつけてやろうとしたのです。脱線してきたけど、脱線こそがおもしろい。
話を戻します。(というか上のパラグラフの後半を加筆していたら話がつながらなくなってきた。) しかも、Fig.2cの赤線の部分を貼り合わせようとすると、点Dと点Fを対応させるためにひっくり返さないといけないから、メビウスの輪になってます。なんかよくわかんないけど意味深い。
ともあれ、そんなわけで、tonnetzひとつでいろいろ考えられておもしろい。ギタリストとしてはFig.1見ながらいかにメジャーダイアトニックコードから離れたコード進行を作れるか考えてみたりします。ニールヤングがよくやるF-D-C-Gみたいなコード進行はDmコードの代わりにDコードにしたものだな、とか。そういえば以前書いたDm7-Cmaj7-Bm7-Aの進行もAmコードの代わりにAコード持ってきたってかんじですな。ロックのパワーコード系のコード進行で3度の音をomitした進行、たとえばA-C-E-G(ニルヴァーナのスメルズライクとか)なんかもAm-C-Em-Gの変形と言えます。マイナーコードのところをメジャーコードにしたり、3度の音をomitしたり、というのはロックっぽいものでの常套手段ですな。そこからさらに外れるのは難しい。そういう意味では、Trafficの'Paper Sun'のG-E-Gm-D-Cm-Bb-G-Aなんてかなりいい感じです。こういう、一小節ごとに転調してるようなサイケなコード進行が大好きです。
そろそろ話がとっちらかってきたんでこのへんで。(まとまった話にしようとしないのは芸風、ということで。)
*注 ここでは3和音のことを考えているので4和音のBm7(b5)コードではなくて、7度のA音を抜いたBm(b5)コードと表記しました。ジャズやロック向けの本とかだとメジャーダイアトニックコードは4和音で書かれていますが、4和音に関しては、上記の3和音を構成する右上の頂点(Cコードだったら点G)のさらに上の点の音(Cコードだったら点B)を付加してゆけばよいことがわかります。これは、Cmaj7 = C + Em、Em7 = Em + Gというふうに考えれば納得。
- / ツイートする
- / 投稿日: 2006年07月14日
- / カテゴリー: [Papers_unclassified]
- / Edit(管理者用)
# HZ
コードの話、たいへん興味深かったです。高校生の時「大学への数学」の「宿題」コーナーでトーラスの問題を解いたこと、大学生の時、ギターを弾き始めたことなど思い出して、いろんなことがつながっているんだと感慨を覚えました。
脳研究を経て今は精神科医の修行の身ですが、楽しく拝見させていただいております。今後も幅広くクオリティーの高いブログを続けられるのを応援しております。
# pooneilコメントどうもありがとうございます。お返事が遅れました。
脳の中のランダム・ウォーカーhttp://hz.cocolog-nifty.com/blog/拝見しました。Anthony Graceの話題もありましたね。岡崎でもこれから統合脳レクチャーコースの一環としてAnthony Graceが話をします。運営側なのではたして講演を聞けるかどうか不明なのですが、可能ならレポートしてみたいと思います。では。
2006年07月09日
■ 和音の幾何学
Science 7/7号。"The Geometry of Musical Chords"
和音を空間上の点として幾何学的に表現して、和音の進行を空間上の動きととらえる、とかそういう話で、ものすごく惹かれるのですが、読んでいる時間がない。そのうちScientific Americanあたりででも紹介されたら読んでみることにしようかと思います(なんか、きれいなグラフィックで表現されてる様子が浮かびます)。単行本出ないですかね。
Perspectivesの"Exploring Musical Space"はenhancedバージョンになっていていろいろリンクされてます。それをみると、そもそもこのような音楽の数学的表現という分野がすでにあって、tonnetzという単音間の関係(=音程, pitch)の空間表現(ドーナツ型=トーラスになるらしい)があるらしい(Wikipediaのエントリ)。そういうのを踏まえた上で音程から和音へ拡張して、和音をorbifold(多様体をさらにgeneralizedしたもの、Wikipediaのエントリ)の中の一点として表現する形式を作りました、というのが今回の論文らしい。幾何学的、といってもかなり抽象的で、多様体とかトポロジーとか群論とかそういう世界らしい。
著者のサイトではChordGeometries 1.1っていうソフトウェアも配布してます。Movieもいろいろあって、おもしろいです。
部分部分で興味を惹かれるネタがあります。Table 1での、オクターブを3等分、4等分、5等分、6等分、7等分していった時を近似する音の組み合わせがXmajor, X7, X9, X11, X13と基本3和音の上にテンションノートを重ねていったものになるんだーとか、たぶんこれ自体は新しいことではないんだろうけど、知らなかったんでおもしろい。
Supporting InformationのFig.1ではcircle of fifth(5度圏: Cから完全5音ずつ上がってゆくと、G,D,A,E,...と重複せずにすべての音程を巡ってまたCに戻ってくるやつ。Wikipediaのエントリ)について言及してるようですし。
ま、わたしはロックンローラーなので音楽理論のことはわかりませんが(<-えー?)。
- / ツイートする
- / 投稿日: 2006年07月09日
- / カテゴリー: [Papers_unclassified]
- / Edit(管理者用)
# yureisoul
はじめまして、yureisoulといいます。いつも参考にさせていただいています。私は音楽理論について無知ですが、こういう話は大好きです。和音の遷移パターンの解析にも郡論が役に立つんですね。しかも曲がった商空間を導入しているところがなかなか興味深いです。快-不快と協和音-不協和音は関連があると言われていますので、たぶん音楽の幾何学構造と人間の知覚の構造にはなんらかの関連があるのではないかでしょうか。将来的にこのような研究と脳研究がつながったら非常に面白いと思います。
# pooneilどうもありがとうございます。「機械仕掛けのこころ」http://yureisoul.blog49.fc2.com/巡回先に入れてます。「君と僕とのKLダイバージェンス」おもしろかったですよ。ふたりの距離は同じように見えて違っている、味わい深すぎる。んでもってコメント見て、あれ、みたいなかんじで。
協和音-不協和音についてもこの論文でやってるみたいですがまだ読んでません。その手前のところでおもしろくなってきたので、またエントリ作りました。ぜひごらんいただければ。
2006年02月15日
■ Gamma-band synchronization, Science '05
昨日のエントリのつづきというか。んで、そのPascal FriesはこのあいだもScience ("Neuronal Coherence as a Mechanism of Effective Corticospinal Interaction")にM1とspinal cordのmotoneuronとのあいだでgamma bandのcoherenceがある、っていう話を出してました。
スゲー、と言いたいところだけれど、どこでもgamma bandってところがどうなのよ、とも思います。じっさい、SN Baker and RN LemonのThe Journal of Physiology '97 "Coherent oscillations in monkey motor cortex and hand muscle EMG show task-dependent modulation"なんかだと、M1からのLFPとmuscleのEMGとのあいだで見られるtask-dependentなcoherenceはbeta band (ここでは20-30Hzあたり)なわけだし。もしくはJNS '05 "Existing Motor State Is Favored at the Expense of New Movement during 13-35 Hz Oscillatory Synchrony in the Human Corticospinal System"とか。
このへん、motorの人はどう考えますか?
- / ツイートする
- / 投稿日: 2006年02月15日
- / カテゴリー: [Papers_unclassified]
- / Edit(管理者用)
2006年02月14日
■ Gamma-band synchronization, Nature 2/9
この分野でひさびさに出たnatureはPascal FriesによるV4 gamma oscillationのattentionによるmodulationの話。
Nature 439, 733-736 "Gamma-band synchronization in visual cortex predicts speed of change detection" Pascal FriesとRobert Desimone。
タイトルに"speed of change detection"なんて言い方してるけど、たんにreaction timeを調べましたということのようですな。以前のScience '01 "Modulation of Oscillatory Neuronal Synchronization by Selective Visual Attention"からの違いは、trial-baseでのreaction timeとgamma oscillationとの相関を見たところにある模様。Oscillationのようなノイジーなデータでtrial-baseの議論ができるというのはすごいことだと思うけれども、それでNatureか?とも思います。
しげさんのところでも採りあげられてます。
- / ツイートする
- / 投稿日: 2006年02月14日
- / カテゴリー: [Papers_unclassified]
- / Edit(管理者用)
2006年01月10日
■ 視床が自発的行動に関わっている?
田中真樹さん@北大の論文がNature neuroscienceに出てます。
Inactivation of the central thalamus delays self-timed saccades - Nature Neuroscience
Central thalamus (おもにVL核、すこしMD核)にムシモルを注入して興奮性伝達を抑制すると、self-timedヴァージョンのmemory-guided saccadeでreaction timeが遅くなった、これが今回あきらかになったこと。
ふつうのmemory-guided saccadeではfixation pointの消灯がgoシグナルとなってsaccadeを開始させるが、このself-timedヴァージョンのmemory-guided saccadeでは、delay期間が800-1600msの間だったらいつsaccadeを開始させてもよい。いわば、ふつうのmemory-guided saccadeは「外的要因によってトリガーされた」なsaccadeであり、self-timedヴァージョンのmemory-guided saccadeのほうは「内的要因によってトリガーされた」saccadeであるわけです。
そういうわけで、self-timedヴァージョンでは、被験者は自分でdelay期間の経過時間を見積もって、最小のdelay期間800ms以降でsaccadeをしようとする。よって、saccadeの開始時間は800-1000msあたりに集中します。そこで、central thalamusへムシモルinjectionすると、saccadeの開始時間は800-1600msくらいに遅くなったと。いっぽうで、ふつうのmemory-guided saccadeのreaction timeもムシモルによって多少遅くはなったけれど、self-timedヴァージョンへの影響よりは弱かった、と。
そういうわけで、reaction timeの意味づけがふつうのmemory-guided saccadeの場合(fixation pointの消灯からsaccade開始までの時間)とself-timedヴァージョン(cueが消えてから、delayの経過を待つ時間があってから、saccade開始するまでの時間)とで違っているので、直接的な比較はあまり意味をなさないように思うのだけれども、ispi-contraの選択性も明確です。
Timingやdurationの測定にcentral thalamusが関わっている、ということでしょうか。でもただ単に時間感覚がずれた、とかいうことだと、ipsi-contralでの選択性があることが説明できないわけです。というわけで、ある行動に選択的であることから、その行動(contraへのsaccade)を自発的に開始するところにcentral thalamusが関わっているのではないか、と言えるわけです。Doug Munozがこの論文をF1000に選んでます。Munozが注目する理由はよくわかります。行動開始がvoluntaryであるか否か、という問題に関わるのではないか、ということでしょう。Monoz自身はprosaccadeをreflexive saccadeとして、antisaccadeをvoluntary saccadeとして扱ってこの問題に取り組んでいます(Nature Reviews Neuroscience 04 "LOOK AWAY: THE ANTI-SACCADE TASK AND THE VOLUNTARY CONTROL OF EYE MOVEMENT")が、今回の論文はそれと問題意識を共有しているように思います。
Memory-guidedであるが故に、where to sacccadeは決まっていて、when to saccadeが未定である、という状況であるところがポイントなのですな。Visually-guidedでself-timedであるような条件ではなにが起こるでしょうか。たとえば、targetはつきっぱなしなのだけれど、target-onからの経過時間によってrewardの量とかが変化するようにしておいて、タイミングを測らないようにしないといけないとか、そんな感じで。
あと根本的問題として、タイミング測定とかなしに、genuinely intrinsic triggerにするにはどうすればよいか、という問題についてとか。
なお、MD核の方はSommer and WurtzのScience '02 "A Pathway in Primate Brain for Internal Monitoring of Movements"によるSC-MD-FEFがSCからのsaccade開始情報のcollorary dischargeを運んでいる、という説で有名になったのだけれども、このへんはほんとうにいろいろなものがあるのだなあ、と。
なあ、Sommer and Wurtzとの関連では、JNP03 "Contribution of Signals Downstream From Adaptation to Saccade Programming"が、central thalamusのrecordingでsmooth pursuitでの結果はJNS05 "Involvement of the Central Thalamus in the Control of Smooth Pursuit Eye Movements"にあります。
追記:2005年06月28日のエントリが関係あるのでリンク。
- / ツイートする
- / 投稿日: 2006年01月10日
- / カテゴリー: [Papers_unclassified]
- / Edit(管理者用)
# viking
はじめまして。友人にこのサイトを教えてもらって以来、興味深く拝見しております。近いテーマで研究しておりますので、特に論文に対するコメントの数々は非常に参考になります。
ところで、こちらのblogではtrackbackの機能は使用されておられないのでしょうか?僕のblogからtrackbackを送ろうとしたのですが、「トラックバックする」をクリックしてもURIが表示されませんので少々戸惑っております。もし機能を停止されているようならば、コメントで代用といたしますので。
どうもはじめまして。trackbackに関してですが、いろいろいじっている過程で使えなくなっていたようです。道理でtrackbackが全然なかったわけでした。というわけで、お知らせいただきまして、どうもありがとうございます。はやめに直しておきたいのですが、すぐには対処できませんので、必要でしたらコメント欄の方で通信をお願いします。お手数かけまして恐縮です。
なお、リンクされていたvikingさんのサイトに行ってみようとしたのですが、ユーザー認証がかかっていて、入れませんでした。
お返事ありがとうございます。しばらくはコメント欄を利用させていただきます。
ところで、認証がかかっていた件ですがSPAM対策等で色々といじっておりましたもので大変失礼しました。設定を変えてみましたので、お試しください。万一アクセスできないようでしたら、またご連絡いただければありがたいです。
2005年10月20日
■ JNP 11月号
- "Laminar Variation in Threshold for Detection of Electrical Excitation of Striate Cortex by Macaques" Edgar A. DeYoe, Jeffrey D. Lewine and Robert W. Doty
- "Psychophysics of Electrical Stimulation of Striate Cortex in Macaques" John R. Bartlett, Edgar A. DeYoe, Robert W. Doty, Barry B. Lee, Jeffrey D. Lewine, Nubio Negrão and William H. Overman, Jr V1へのmicrostimulationによるphosphoneに関する論文2連発。"Preliminary communications of these data have been presented (Bartlett et al. 1977; Lee et al. 1973)." なんて恐ろしいことが書いてある。30年ですよ! Tehovnikの論文とか、さいきんの流れであることを認識したのでしょうか?
- "Low-Frequency Oscillations Arising From Competitive Interactions Between Visual Stimuli in Macaque Inferotemporal Cortex" この発火パターンには見覚えがあって、なんか意味はあるのだと思います。菅生さんのNature '99とかでの早い部分の発火と遅い部分の発火でのコーディングの違いとか、Richmond and OpticanのITニューロンでの一連の仕事とか、そのへんはみんなたぶんこの問題と関連しているわけで。
- "Dynamic Circuitry for Updating Spatial Representations. I. Behavioral Evidence for Interhemispheric Transfer in the Split-Brain Macaque"
- "Dynamic Circuitry for Updating Spatial Representations. II. Physiological Evidence for Interhemispheric Transfer in Area LIP of the Split-Brain Macaque" Carol L. Colbyによる連報。JNPはあいかわらず連報が多いですね。JNSとかはそういうのはdiscourageさせる、とか書いてあった気がする。Split-Brainを使ってInterhemispheric Transferの議論をしてる。要旨にも書いてあるけど、split-brainにすればinterhemisphericなremappingは起こらないだろう、と予想して実験を始めたんだけれどそうでもなかったんで、なんとか議論して論文にした、というかんじなんだと思います。Tomita et al Nature '99では複雑なshapeを使うことでsubcorticalなtransferがあり得ないだろう、ということをよりどころにしていたのに対して、今回のようなパラダイムではsubcorticalな領域、pulvinarなどのthalamusやbasal gangliaの影響をまったく取り除くことができない。そのへんに問題があったのではないかと読まずにコメント(読んだ人がつっこんでほしい)。
- / ツイートする
- / 投稿日: 2005年10月20日
- / カテゴリー: [Papers_unclassified]
- / Edit(管理者用)
2005年08月11日
■ Anticorrelated random-dot stereograms
重要なのだけど、とりあえず論文リストだけ作成。
- Nature "Responses of primary visual cortical neurons to binocular disparity without depth perception" CUMMING AND PARKER
- Perception '98 "Disparity detection in anticorrelated stereograms" Cumming BG, Shapiro SE, Parker AJ.
- JNS "Rejection of False Matches for Binocular Correspondence in Macaque Visual Cortical Area V4" Seiji Tanabe, Kazumasa Umeda, and Ichiro Fujita
- JNP "A Stereoscopic Look at Visual Cortex" Peter Neri
- Vision Research "Unconscious adaptation: a new illusion of depth induced by stimulus features without depth" Ryusuke Hayashi, Yoichi Miyawaki, Taro Maeda and Susumu Tachi
- Vision Research '93 "Depth in anticorrelated stereograms: effects of spatial density and interocular delay"
- / ツイートする
- / 投稿日: 2005年08月11日
- / カテゴリー: [Papers_unclassified]
- / Edit(管理者用)
2005年08月09日
■ fMRIのBOLDシグナルと神経活動
Science 8/5でback-to-back。
"Hemodynamic Signals Correlate Tightly with Synchronized Gamma Oscillations" Wolf Singerたち。麻酔下のcatではBOLDシグナルはスパイクよりはLFPのgammma oscillationのpowerの方によく相関していた。
"Coupling Between Neuronal Firing, Field Potentials, and fMRI in Human Auditory Cortex" Rafael Malach。去年出たintersubject correlationの手法を用いて、awake humanの聴覚野ではBOLDシグナルがspikeとよく相関している(r=0.75だと!)ことを強調しています。Gamma frequencyのLFPもBOLDとよく相関している、としているので前者の論文や以前のLogothetisらの論文とかと直接対立するものではなさそうです。
ちょうど1年前のエントリ(20040809)で関連する事項を扱っています。Logothetisの論文についてとか。ご隠居の最後のコメントで言及されているLauritzenらの仕事をリストしておきましょう:
- JP '98 "Modification of activity-dependent increases of cerebral blood flow by excitatory synaptic activity and spikes in rat cerebellar cortex"
- JP '00 "Temporal coupling between neuronal activity and blood flow in rat cerebellar cortex as indicated by field potential analysis"
- JNS '03 "Brain function and neurophysiological correlates of signals used in functional neuroimaging." (Review)
- / ツイートする
- / 投稿日: 2005年08月09日
- / カテゴリー: [Papers_unclassified]
- / Edit(管理者用)
2005年08月04日
■ Science Anosognosia(疾病否認)論文
そういえば昨年のエントリ(20040708)でPassinghamのScience '04 "That's My Hand! Activity in Premotor Cortex Reflects Feeling of Ownership of a Limb"に言及しましたけど、これも関係してきますな。自分の腕に対するfeedling of ownershipがあるかどうかはpremotorが関わってくる、というのは今回のScience論文とつなげて考えると納得のいくかんじがあります。つまり、半側空間無視と疾病否認との関係にさらに「身体失認」を入れて考えるということですな。あと、PassinghamのScience '04のPerspectivesが"Probing the Neural Basis of Body Ownership"にあります。
追記:最新のJNSにも関連論文あり。"Awareness of the Functioning of One's Own Limbs Mediated by the Insular Cortex?" Hans-Otto Karnath。例の、hemineglectの原因部位はparietalではなくてtemporalだとするNature論文の著者です。(参考:Spatial hemineglectスレッド)
Patients with anosognosia for hemiplegia/hemiparesisとhemiplegic/hemiparetic patients without anosognosiaとを比較して、その損傷領域の差はright posterior insulaであるとしてます。
- / ツイートする
- / 投稿日: 2005年08月04日
- / カテゴリー: [Papers_unclassified]
- / Edit(管理者用)
2005年07月22日
■ Science Anosognosia(疾病否認)論文
7/20のエントリからの続きです。ラマチャンドランの「脳のなかの幽霊」から抜き書き。
脳卒中で入院しているドッズ夫人は自分が入院していることなどの状況に関してはよくわかっているのに、自分が左手を動かせないことをけっして認めようとしない、というところの記述です。
「手はどうですか? のばしてみてください。うごかせますか?」さらに、ラマチャンドランはドッズ夫人が言っていることと視覚情報とが矛盾していることを指摘したらどう対処するかを調べます。(前にも書きましたように、疾病否認は半側空間無視を伴うことが多いので、ふだんは患者さんはその視覚情報自体を無視しています。そこで医師が無視している視覚情報に注意を向けるように患者さんを誘導したら何が起こるか、というわけです。)
ドッズ夫人は私の質問にややむっとしているようだった。「もちろん動きますよ」
「右手を使えますか?」
「ええ」
「左手はどうですか?」
「ええ、左手も使えます」
「両手とも同じくらいしっかりしていますか?」
「ええ、両手とも同じようにしっかりしてます」
p.175
「ドッズさん、右手で私の鼻をさわれますか?」ドッズ夫人は視覚情報よりも、自分の左手は動くという信念を重視し、その矛盾を作話によって対処しています。
彼女は何の支障もなくそうした。
「左手で私の鼻をさわれますか?」
彼女の手は麻痺したままからだの前におかれていた。
「ええ、もちろんさわっていますよ」
「実際にさわっているのが自分で見えますか?」
「ええ見えます。先生の顔から1インチと離れていません」p.176
私はもう一つだけ聞いてみることにした。「ドッズさん、手をたたけますか?」
彼女はあきらめたように辛抱強く答えた。「もちろんたたけます」
「たたいてもらえますか?」
ドッズ夫人は私の顔をちらっと見て、右手で手をたたく動作をした。体のまんなかあたりで想像の手とたたきあっているようなしぐさだった。
「いま手をたたいていますか?」
「ええ、たたいています」p.176
このシーンからこの章の題名「片手が鳴る音」が付けられています。 白隠の公案の「隻手の音声」ですな。
この記述のほかにも、「今日は肩が痛いので左手を動かせない」という言い方で対処する例、麻痺している左手を自分の手と認めず、死体の腕がベッドの中に入っていると主張する患者さんの例などの記述があります。
この症例で重要なのは、視覚情報を凌駕するほどに、自分の腕は指令通りちゃんと動いている、という信念が強いということです。そしておそらくそれは、麻痺した腕に運動指令を出したにも関わらず、それが正しく実行されたというふうに内部でモニターしている機構が感受したことに基づいているのでしょう。視覚って他のモダリティよりもものすごい強烈なものだと一般的には考えられているわけですが、それがひっくり返るところに驚きがあります。(「百聞は一見にしかず」もいったん見てしまったものの信憑性の強さを強調しているわけです。もっとも、このばあいの視覚と聴覚が示している情報はそもそも直接性が違うのでmisleadingですが。)
そういうわけで、身体図式の問題としても、out-of-body experienceとか、もっと身近な例でいえば、金縛りとかで起こっていることとつきあわせて考える必要があるでしょう。金縛りを同じモデルで考えてみましょう。[運動の目標]と[実際の運動をsensory feedbackとしてモニターしたもの]との誤差を修正するようなループと、[運動の目標]と[運動指令から実際に実現したであろう運動を内部モデルによってfeedforward的に作ってやったもの]との誤差を修正するようなループとがあるというわけです。金縛りでは運動目標を立てて、運動指令は出ていて、しかし実際の筋肉の収縮は起こっていなくて、動いていないというsensory feedbackは戻ってきているのだけれども、内部モデルでモニターしているところでは動いたように感受している、という感じではないでしょうか。二つのループの間での矛盾が私たちをあわてさせるというわけです。なんつーか金縛りの時の空振り感ってのは、内部モデルとしては動いている感じがあるからではないですか、たぶん。このへん微妙ですな。というかそもそも金縛りって英語ではなんと言うんでしょうか。
というあたりで中途半端にこの項ネタ切れ終了。
- / ツイートする
- / 投稿日: 2005年07月22日
- / カテゴリー: [Papers_unclassified]
- / Edit(管理者用)
# いととんぼ
関連の話題に、他人がわき腹とかを急にさわってくるとくすぐったいのに、自分でくすぐっても、ちっともくすぐったくないという現象がありますね。あれも、自分でくすぐるときには、運動司令とその帰結の内部モデルと、さわれたという感覚情報のすりあわせができるのに対して、人からくすぐられたときには前者がないことが関係しているという主張があります。20日に引用されている論文のBlakemoreさん(かのColin Blakemoreのお嬢さんだそうです。1、2年前、淡路島でのシンポジウムにこられていました)がそのような研究をされていました。しかし、小さいころは、なんであんなにくすぐったかったのか。最近は、くすぐってくれる人もいないなあ、ナンテ。
# pooneil自分で自分をくすぐる話はSarah-Jayne BlakemoreのNature neuroscience '98 "Central cancellation of self-produced tickle sensation" http://www.nature.com/neuro/journal/v1/n7/abs/nn1198_635.htmlですね。このへん追記しておきました。Sarah BlakemoreはColin Blakemoreの娘さんなんですか。Colin Blakemoreのほうは現在は社会に向けての活動の方が忙しそうですね。なお、我が家はまだ子どもが小さいので毎朝子どもたちとくすぐりあいっこしてます。
2005年07月20日
■ Science Anosognosia(疾病否認)論文
Science Anosognosia(疾病否認)論文
"Shared Cortical Anatomy for Motor Awareness and Motor Control"
昨日のつづき。
著者の結論は疾病否認のモデルとして提出されているいくつかの論文とつきあわせて考える価値があるでしょう。とくに重要なのは、内部モデル(forward model)によるefference copyのような、sensory feedbackそのものよりも速い機構を想定しているものです。
Heilman KM, Barrett AM, Adair JCのPhilos Trans R Soc Lond B Biol Sci. '98 "Possible mechanisms of anosognosia: a defect in self-awareness"
Frith, Blakemore and WolpertのPhilos Trans R Soc Lond B Biol Sci. '00 "Abnormalities in the awareness and control of action"
Cortex '05 "Anosognosia and body representations forty years later" (pdf file)
Mind & Language '05 "Anosognosia and the Two-factor Theory of Delusions"(pdf file)
あ、いま気づいたけど、"Cortex" ("Cerebral Cortex"ぢゃなくて)ってpdfみれますね。PubMedでリンクがつかないので無理かと思って論文読まずにきたのですが、これからはどんどん読めそう。Cortex誌にはほかにも疾病否認に関する重要そうな論文として
"ANOSOGNOSIA FOR PLEGIA: SPECIFICITY, EXTENSION, PARTIALITY AND DISUNITY OF BODILY UNAWARENESS" Anthony J. Marcel, Richard Tegnér and Ian Nimmo-Smith
"ANOSOGNOSIA: THE NEUROLOGY OF BELIEFS AND UNCERTAINTIES" Patrik Vuilleumier
"THE ANATOMY OF ANOSOGNOSIA FOR HEMIPLEGIA: A META-ANALYSIS" Lorenzo Pia, Marco Neppi-Modona, Raffaella Ricci and Anna Berti
があります。
また、Ramachandranが「脳のなかの幽霊」で書いていることは非常に興味深いので、原著に当たる価値があるかもしれません:
Conscious Cogn. '95 "Anosognosia in parietal lobe syndrome"
Nature '96 "Denial of disabilities in anosognosia"
また、疾病否認はbody schemaについて考えるにあたっても重要な事例であり、幻肢とともにメルロ=ポンティの「知覚の現象学」で扱われています。
メルロ=ポンティ『知覚の現象学』を読む(前半のみ)
身体図式と実存 メルロ=ポンティの身体論 竹内 幸哉
明日は「脳のなかの幽霊」から関連部位を抜き書き、ということで。
- / ツイートする
- / 投稿日: 2005年07月20日
- / カテゴリー: [Papers_unclassified]
- / Edit(管理者用)
2005年07月19日
■ Science Anosognosia(疾病否認)論文
"Shared Cortical Anatomy for Motor Awareness and Motor Control"

そんでもって、疾病否認の原因部位がどこかということはこれまでそんなには明らかになっていませんでしたが、右頭頂葉の障害時に左視野の半側空間無視を伴って見られるということはよく知られていました。そこで今回の論文では[半側空間無視があり(N+)、疾病否認がある(A+)患者さんのグループ]と[半側空間無視があり(N+)、疾病否認がない(A-)患者さんのグループ]とで脳の損傷部位を比較しました。その結果、N+A+グループで選択的に補足運動野(BA6)に損傷があるということを見いだしました。N+A+グループおよびN+A-グループ両方に関わるのは頭頂葉でした。というわけで、原因部位はじつは頭頂葉ではなくて前頭葉だった、というわけです。
もっとも、これだけの結果からは、疾病否認の原因部位は頭頂葉から補足運動野へとつながるネットワークだ、ということでもよいわけですが、著者たちは[半側空間無視がなく(N-)、疾病否認がある(A+)という患者さん(一人)]でもこの補足運動野あたりに損傷があることを示しています。
そういうわけで、著者らの結論としては、疾病否認の原因部位である補足運動野が実際の運動を生成させるだけでなく、生成した運動のモニターもしている、ということになります。
うーむ、表現に正確さが欠けているのは印刷した論文がいま手元にないからです。
明日に続きます。
- / ツイートする
- / 投稿日: 2005年07月19日
- / カテゴリー: [Papers_unclassified]
- / Edit(管理者用)
2005年06月27日
■ 木村研CM核論文
20050618のエントリにmmmmさんからコメントをいただきました。ありがとうございます。埋もれないようにここで続けます。
で、なるほど、[response bias +/-] * [CS+/-] のinteractionこそがneuesである、ということですね。わたしが書いてたことはけっきょく「negative prediction errorをコードしている」ということでしかありませんでした。ラボのサイトでの説明文の「期待とは異なる状況が発生した場合に,CM核は企画済みや実行中の行動をリセットしたり,切替えたりすることに深く関わる」という表現はそのへんのニュアンスをくんでいるように思えてきました。
さて、CM/Pfニューロンは彦坂研の1DR task / ADR taskのときにどう活動するか考えてみます。1DR taskではrewardが出ない方向のときに活動して、ADR taskではどの方向でも同じくらい活動する、とたんにcaudateの逆みたいに反応する、というところでしょうか。だとすると、hikosaka taskで見られるものよりもより多くの情報を今回のgo/nogoタスクが持っているとしたらそれは何か、というところがポイントで、まだ私よくわかっておりません。それがmmmmさんご指摘のPavlovian conditioningとの比較なのでしょうか。なんにしろ、もうすこし読みこんでみようと思います。
- / ツイートする
- / 投稿日: 2005年06月27日
- / カテゴリー: [Papers_unclassified]
- / Edit(管理者用)
2005年06月21日
■ 木村研CM核論文
20050618のエントリ(木村研CM核論文)にさっそくコメントが。どうもありがとうございます。痛みと心の痛み、とかいらんこと書いてみました。
さらにいらんことばかり書きますが、この話をプロスペクト理論とつなげる、ってのはどうでしょうか。我々が同じ大きさの損失を利益よりも重く見積もってしまうというところにCM核からのシグナルに影響される度合いをみる、というわけです。いわば、中脳ドパミン系は合理的な意思決定に関わり、CM/Pfは非合理な意思決定に関わる。主観的な期待効用と客観的な期待効用とのあいだのずれには個人差があるけれど、それがCM/Pf核のactivationの大きさの差と相関するとか。昨年のSFNでのGlimcherのfMRIではなんか関連すること言ってませんでしたかね。ちょっととばしすぎでしたか。
ま、中身読んでから書け、って話でもありますが、詳しい方にコメントしていただけると助かります。CM核とamygdalaはつながってますか?
- / ツイートする
- / 投稿日: 2005年06月21日
- / カテゴリー: [Papers_unclassified]
- / Edit(管理者用)
2005年06月18日
■ Science 6/17
Complementary Process to Response Bias in the Centromedian Nucleus of the Thalamus 木村研@京都府立医大より。
視床のCM核からのsingle-unitとmicrostimulation。Go / Nogo x Large reward / small rewardの組み合わせがブロックごとに変わる。
Large rewardのときよりもsmall rewardのほうがresponse latencyは遅くなるし、CM核のニューロン活動はsmall rewardの時の方がよく発火する。
CM核へのmicrostimulationでGo responseが遅くなるのだけれど、同じGo trialでもlarge rewardのときのほうがsmall rewardのときよりもmicrostimulationの効果が大きい。
というわけで、ようするにrewardが少なくて不本意でいやいや行動するときにCM核が活動するのだ、というわけです。Asahi.comでは「すし屋でトロを注文したら品切れで、しかたなく赤身を頼む」なんて表現をしてます。木村實先生は「期待通りに物事が進むことは少ない。ベストの選択肢が選べなくても、パニックに陥らず、次善の選択をするのは知的な行動だ。その脳のメカニズム解明の突破口になる」とコメントしてます。どうあれ、CM核が中脳ドパミン系とはresponse biasに関して違った役割を果たしている、というこの論文の主張は明確です。
2003年の名古屋での神経科学大会のときにこの話のポスターが出ていて、ファーストオーサーの方にいろいろ聞いた覚えがあります。そのときはmicrostimulationのデータは無かったのではないでしょうか。(追記:ラボのウェブサイトの説明を見る限り、アブストには載っていないけど発表はした様子)
関連する前報におなじファーストオーサーによるJNP '02 "Participation of the Thalamic CM-Pf Complex in Attentional Orienting" Takafumi Minamimoto and Minoru Kimura、それからJNP '01 "Neurons in the Thalamic CM-Pf Complex Supply Striatal Neurons With Information About Behaviorally Significant Sensory Events"があります。(ラボのウェブサイトに説明あり。)
木村實先生によるNeuroscience Research '04 "Monitoring and switching of cortico-basal ganglia loop functions by the thalamo-striatal system"のFig.2とFig.3が今回の論文のストーリーの下敷きになっているようです。Cortex->Striatum->GP/SNr->CM/Pf->Cortexというループがあって、Striatumはドパミン系(SNcとか)およびCM/Pf核からmodulationを受けます。つまり、Corticobasal ganglia loopsでのresponse biasのmodulationには中脳ドパミン系によるものと、CM/Pf核による相補的な過程とがある、というストーリーらしいです。CM/Pf核のもつ「相補的な過程」が、中脳ドパミン系がTD errorを出力するといった強化学習的枠組みに対してはどういう位置づけになるのか、というあたりが重要となるでしょう。TD errorのようなrewardの大小に対して中立的なものではなくて、rewardが少ないとき>多いとき、という極性を持っているわけですから。
Neuroscience Research '04ではJNP '02を元にして、salient eventの情報がCM/Pf核を通してresponse biasを変えるということを強調しているようですが、さいきんのRedgraveらによるSNcのドパミンニューロンが上丘から視覚情報を受け取っているという話(Science '05 "How Visual Stimuli Activate Dopaminergic Neurons at Short Latency"およびNature Neuroscience '03 "A direct projection from superior colliculus to substantia nigra for detecting salient visual events")とつきあわせると、SNcとCM/Pf核のどっちがStriatumにsalient eventの情報を持ってきているのか、というあたりを考えるのがおもしろいかもしれません。
- / ツイートする
- / 投稿日: 2005年06月18日
- / カテゴリー: [Papers_unclassified]
- / Edit(管理者用)
# Striatumの人
CM-Pfは痛みの中継核でもあり、SalientなEventの情報といってもドーパミン系とは対極にあると思います。Redgraveらの主張と対比して考えると確かに面白いかなと思います。
# pooneil>>Striatumの人
コメントありがとうございます。おお!「CM-Pfは痛みの中継核」なのですか。知りませんでした。言ってみれば、損失という「心の痛み」でもCM/Pfが活動してしまう、というわけですね。おもしろくしすぎでしょうか。
お久しぶりです。先日はお会いできず残念でしたが、お子様がお誕生とのこと、おめでとうございます。
さて、Minamimoto et al., 2005, Scienceですが、Thalamostriatal systemの役割について一つの魅力的な仮説を提出した点で面白い論文だと思いますし、CM核がそこで中核的な役割を果たしているという点にも同意できるのですが、この論文のみを読んだ当初、negative prediction errorをコードしているという以上のことを示せているかどうかに疑問が残りました。
が、Matsumoto et al., 2001, JNPで、Response biasとは無関係のPavlovian conditioning課題での応答を見ており、ここではCS+に対する応答とCS-に対する応答との間に差がないことを報告しています。これをコントロール実験だと思えば、確かにResponse biasが必要な課題遂行中に限って、CS- > CS+という顕著な傾向が出るようです。同じ細胞でこのコントロール実験をして比較結果を示すのがベストだったと思いますが、少なくともPavlovian conditioningでの以前の結果をきちんと論文に書いてそれとの違いをdiscussしてくれないと、CM核に詳しい読者以外からは、正当な評価を受け損ねる恐れがあると思いました。
どうもご無沙汰しております。おかげさまで、もう三人目です。毎晩三人の子供を風呂に入れてます。このあいだの出張はそういうわけでドタキャンとなってしまい、残念でした。次の機会は神経科学大会でしょうか。お話できるのを楽しみにしております。つづきは上の方に書きました。
2005年01月24日
■ JNS 1/19 Out-of-Body Experience
"Linking Out-of-Body Experience and Self Processing to Mental Own-Body Imagery at the Temporoparietal Junction." Olaf Blanke, Christine Mohr, Christoph M. Michel, Alvaro Pascual-Leone, Peter Brugger, Margitta Seeck, Theodor Landis, and Gregor Thut [EndNote format]
Olaf Blankeはこれまでにも
Brain '04 "Out-of-body experience and autoscopy of neurological origin." Brain 127: 243-258 Olaf Blanke, Theodor Landis, Laurent Spinelli, and Margitta Seeck [EndNote format]
Nature '02 (brief communications) "Stimulating illusory own-body perceptions." Nature 419, 269 - 270 (19 September 2002); OLAF BLANKE, STPHANIE ORTIGUE, THEODOR LANDIS and MARGITTA SEECK [ris format]
などを立て続けに出して、'Out-of-body' experiences (OBEs)に関する仕事を続けています。
cogniさんのところの1/20にも記載がありますように、temporoparietal junction (TPJ)のはたす役割、という意味でも重要な知見です。うちのサイトのOptic ataxiaに関するスレッドやSpatial hemineglectに関するスレッドともおおいに関係があります。
- / ツイートする
- / 投稿日: 2005年01月24日
- / カテゴリー: [Papers_unclassified]
- / Edit(管理者用)
2004年09月27日
■ Nature 9/23
"A frequency-dependent switch from inhibition to excitation in a hippocampal unitary circuit." BEAT H. GÄHWILER
おお!Gähwilerとはなつかしい名前が。私は以前slice cultureで電気生理をやっていたのですが、Gähwilerはslice cultureのオリジネーターです。スライドガラスにplasma clotで海馬スライスをくっつけてやって、ぐるぐる回しつづけながら培養するとスライスの厚みが一層だけになって、よりたくさんのニューロン間の結合が形成されます。この系を使って彼らは以前もCA3とCA1の錐体細胞でシナプス結合のあるペア(unitary circuit)からパッチクランプをして、ひとつのCA3からの入力でCA1の錐体細胞にどれだけの影響があるかとか可塑的変化を起こせるかとかを検証していて、すげーと思った覚えがあります。スライスが一層になる過程でacute sliceと比べてシナプス結合が多くなってしまうことをどのくらいポジティブに捉えるかが問題なわけですが。これが約10年前の話*1。
今回はこの系を使って、dentate gyrusのgranule cellとCA3の錐体細胞とからpair recordingをして、preのgranule cell一個の発火頻度によってpostのCA3がどのようにmodulateされるかを調べたら、preの発火頻度によってpostへの影響が全体として興奮性になったり抑制性になったりすることがわかった、というものです。これがNature級なのかどうかはワシにはもはやわかりゃせんのじゃがの(ご隠居になってみる)。
*1:ちなみに当時私はミリポアのmembrane上での静置培養のほうを使っており、こちらのほうはスライスが一層になることもなく、pyramidal layerが広がることもないので、Gähwilerらのやり方よりもよいのではないかという信念を持っていました。とくにfield potentialを記録するにあたっては。
- / ツイートする
- / 投稿日: 2004年09月27日
- / カテゴリー: [Papers_unclassified]
- / Edit(管理者用)
2004年08月06日
■ Current Biology
"Pigeon Homing along Highways and Exits."
ハトが遠距離飛行をするときに高速道路や鉄道の線路に沿って飛ぶ習性があることはこれまでに経験的にわかっていたけれどそれを系統的に調べた人はいません。そこで著者らはGPSを駆使してそれを検証した、というものです。
Fig.1を見るとたしかに、ハトの軌跡(赤)は高速道路(緑)に沿っています。そしてこれが偶然によるものでないことは、Fig.2のシミュレーションから明確です(道路を無視して飛んでいたら、もっと軌跡はバラける)。
しかもFig.8の通り、飛ぶ回数ごとに道路をランドマークにして飛ぶ比率が増えてゆくわけで、つまり経験と記憶によってこの習性が形成、強化されているわけです。
しかもこの行動は目的地に対して一番近道を行っているか、という意味において最適解ではないのですな。つまり、飛ぶ回数ごとに、道路に沿って飛ぶ比率が増えるので、実のところ回り道になってしまっているのです。とはいえ、そのような行動は他の側面から見て最適である可能性はあるはずですが(他のハトと軌跡を同じくすることによる安全性の面でのメリットとか。もしくはランドマークなしに飛ぶとうろうろと目的地を探して、けっきょく飛行距離が長くなってしまうとか)。
なんにしろこの話は"navigation"の話であり、spatial memoryの話なので、将来なんかそういうneurowthologicalな展開があると面白いと思います。
- / ツイートする
- / 投稿日: 2004年08月06日
- / カテゴリー: [Papers_unclassified]
- / Edit(管理者用)
2004年08月05日
■ Current Biology
Publishした頃に書いたんだけれど、最近書くネタが多かったので貼りそびれてました。
"Experience in Early Infancy Is Indispensable for Color Perception." 杉田先生@ 産総研。
これまでの杉田先生の業績はすばらしくかつ独特で、プロジェクトごとの成果をNatureにshort paperとして出しつづけていて、JNSやJNPのようなfull paperはまったく書いておられません。今回の論文も含めて、どの論文もアッと言わせられるような大胆かつ鋭いところがあります。
- Nature '03 "Implicit estimation of sound-arrival time." (産総研のプレスリリース)
- Nature '99 "Grouping of image fragments in primary visual cortex."
- Nature '96 "Global plasticity in adult visual cortex following reversal of visual input." 逆さめがね(左右反転)をかけたsubjectの適応段階でのV1からのrecording。
乳幼児期の視覚体験がその後の色彩感覚に決定的な影響を与える
ポイント
- 色の恒常性を含めて色彩感覚は生まれながらに持っているものと考えられてきたが、乳幼児期の視覚体験によって獲得されることが明らかになった。
- また、視覚体験が受容器官(網膜)ではなく大脳皮質に効果を及ぼしていることも同時に明らかになった。
これまでも生後直後の「臨界期」の視覚経験がV1ニューロンの左右の眼優位性(ocular dominance)や線分の方位などへの感受性に影響を及ぼすことについては盛んに調べられてきましたが、そのような発達時の可塑性が色の知覚に関してどうなっているかを扱った人はおそらくいないはずです。まったくの盲点だったというか、この論文のこの点がまずアッと言わせられるところです。そしてしかもその結果として、色弁別はとりあえずできるけれども色の恒常性が障害されるということを示したわけです。これもまた驚きで、そのような結果になるということはおそらく、retinal ganglion cellの回路(L-Mによるred-green軸の形成や(L+M)-Sによるyellow-bue軸の形成など)あたりはおそらくintactで、大脳皮質のどこかのレベル(V4あたり)での可塑性による結果であるわけです。(たぶんV1ではないであろうことも方位選択性の可塑性とかと比べた特色であると言えます。)
今回の結果にそれのneural correlateを見つけてくっつければこんどもNature級の論文だったと思われますが、電気生理のデータに関しては今後のこととなりそうです。
また、そもそも、色というのは視覚意識の問題にとって重要な位置をしめています。この実験で実現している色の識別とはわれわれが体験しているような色とその識別なのでしょうか。色の恒常性がないときにわれわれの色の視覚意識と色間の関係で作り上げられる空間はどのように変化するのでしょうか。
- / ツイートする
- / 投稿日: 2004年08月05日
- / カテゴリー: [Papers_unclassified]
- / Edit(管理者用)
# mds
興味深い論文ですね。早速読んでみます。「〜乳幼児期の視覚体験によって獲得されることが明らかになった」の行はプレスリリース特有の言い過ぎ、というか精確さが欠けている感がぬぐえなくもないですが、色の恒常性が発達過程で獲得されるものであることを示したとは・・・着眼点の鋭さにあっとさせられます。これから後の実験が多く続きそうな、非常に発展性のあるテーマですね。弁別というか波長への反応性については、鳥居修晃先生の報告(ISBN:4130111116)などで、先天盲開眼者の初期視覚が色に対して強い鋭敏性を持っている事が明らかにされていますが、やはり単純な色の感覚と電磁波の波長とは一対一対応するものであり、恒常性のような「高次過程」のみが発達によって可塑的に変化するものなのでしょうか?それとも感覚レベルの色の意識体験でさえも、初期経験により変化させることが可能なのでしょうか・・・?これらは飼育環境の変化で、有用なデータが得られそうです。また、これらのサルを通常環境下で飼育していった時に、どのような変化が起こるのか(初期経験ではなくlate onsetな可塑的変化)も私としては興味深いなので、是非取り組んで頂きたいテーマですね。もう、飛び跳ねたいくらい(!)面白い。ところで使いやすいです。このコメント欄。
# pooneilおひさしぶりです。http://d.hatena.ne.jp/mds/見てきましたよ。> やはり単純な色の感覚と電磁波の波長とは一対一対応するものであり、> 恒常性のような「高次過程」のみが発達によって可塑的に変化するものなのでしょうか?このへんが一番面白いところなのではないかと思います。線分なら単にコントラストがぼやけて見えるようなことが容易に想像できるわけですが、色の経験が変容されるとしたらいったい何が起こるんでしょう。色空間の位相的関係は保持されたままそれが縮んだりするんでしょうか。色と電磁波の波長とは一対一対応はしないのではないでしょうか。黄色の単波長光だろうと緑と赤の混色だろうと同じ黄色として感じますよね。そのような混色の関係が変わって色空間が歪むであろうことは予想できるわけですが。ところで「先天盲開眼者の視覚世界」、ちょうど別件で読んでいたところです。重要ですよね。しかしこれ読みすすめててもやっぱりどう感じているのかはよくわからないんです。色の識別はだんだんできるようになるし、色名もどんどん付加されていって分化しているわけだけど、その分化する前に見えていた色(たとえば、名づけることができなかった緑)は、分化した後に呈示された同じ色と同じように見えていたのか。クオリア自体が変わったのか(暗くてよくわからないものからビビッドな緑へ)、それともその緑を緑たらしめる赤や緑とも相対的関係が変わっただけなのか(緑-赤の軸ができるまでは緑は青-黄または白-黒の軸からは分離されえない)。うーむ、もっと読み進めてみます。
2004年08月03日
■ Nature 6/17
"Structural basis of long-term potentiation in single dendritic spines." 河西研。
がJCで採り上げられたときのディスカッション項目が"Ryohei’s Neuroscience Notes"の7/30にあります。うーむ、さすが。(iii)と(iv)はラボ間のプロトコルの違いみたいなところでうやむやになってしまうかもしれないけれど、(ii)についてはよくよく検討しなければならなさそうです。Imagingと電気生理の対応はFig.4だけのようですので。テタヌス刺激のspine間でのspecificityとかはimagingのデータと電気生理のデータの対応という意味で重要そうです。
ところでlarge spineとsmall spineとはどういう関係にあるのでしょうか。もうpotentiationしてsaturateいるのがlarge spineだとしたら、LTDを誘導したらlarge spineはsmall spineになるのでしょうか。このへんは(i)のどのくらいパラレルであるかと関わる問題です。それに、large spineとsmall spineとはスイッチのon-off的な二つのdistincなpopulationではなさそう(suppl.Fig.2)なので、もっと確率的なものと捉えた方がよさそうで、著者のvolume=0.1micrometer^3をcriteriaとするのはかなり便宜的なもののようです。
ま、そのうち直接話を聞く機会もあるでしょうからこんど聞いてみようっと(建物が2キロくらい離れているので普段まったく会わない)。
追記:と書いたら河西研で平瀬さん@理研のセミナーがあったので聞きに行ってきました。それについては明日レポートします。Natureに関してはまたの機会ということで。
- / ツイートする
- / 投稿日: 2004年08月03日
- / カテゴリー: [Papers_unclassified]
- / Edit(管理者用)
2004年07月31日
■ Science AOP
"Distinct Ensemble Codes in Hippocampal Areas CA3 and CA1." Edvard I. Moser。
先週のNature("Comparison of population coherence of place cells in hippocampal subfields CA1 and CA3.")へのコメンタリかと思ったら違うようです。"We examined the function of this structural differentiation ..."なんて書いてある。CA3-CA1のfunctionのdissociationを出すのに競走をしていたようですな。しかも微妙にずれた出版日。Natureの方はreceive 1/8、accept 6/10で online publicationが 6/30、Scienceの方はsubmit 5/13、accept 7/8でonline publicationが 7/22、というわけでNatureの方が早かったけど、(その情報を察知してか、)Scienceのほうが追い上げたということらしい。Journal clubで採り上げる人はこの二つをセットにしてプレゼンしないと、あっちの方はどうなのと問い詰められること必至ですな、これは。
内容については紙で出版されてからにします。ていうかうちではAOPのpdf取れないんですよね。
Edvard I. Moserといえば私にとってはScience '98 "Impaired Spatial Learning after Saturation of Long-Term Potentiation."です。Barnes and McNaughtonの"Recovery of spatial learning deficits after decay of electrically induced synaptic enhancement in the hippocampus."みたいな、LTPを海馬全体に惹き起こしてやると空間学習がimpairされるというやつでした。なんかすげ―感心した憶えがあるのだけれど、どの点だったか思い出せない。SFNでこの内容を見たときは夫婦並んでポスターで、そろって美男美女だった印象がある、どうでもいいことだが。
Science '02 "Place Cells and Place Recognition Maintained by Direct Entorhinal-Hippocampal Circuitry."のほうはCA3 -> CA1の入力を取り除いてentorhinal cortex -> CA1の入力だけで何ができるか、という議論だから、今回のCA1-CA3のfunctional dissociationへ向かうのは当然の流れだったと言えましょう。
- / ツイートする
- / 投稿日: 2004年07月31日
- / カテゴリー: [Papers_unclassified]
- / Edit(管理者用)
2004年07月12日
■ Trends in Cognitive Sciences 6月号
"Neuronal representations of cognitive state: reward or attention?" John H. R. Maunsell。世で研究されているattentionやreward expectancyのneural correlateはしばしば分離できていないことを指摘している。その理由としてabstではreward contingencyのパラメータのみがそれらの研究では振られていることを挙げています。このへんの分野の概念の複雑化と曖昧さとを考えるととても重要です。将来この辺の概念はもっと整理され、それぞれの概念間の関係がより確立していくことでまだしばらく先はやることはあるでしょうが*1、次は何か、それが問題なのです。
*1:実際には今まで40年の歴史を見て考えればしばしば複雑化の道を進むだけだったりするのだけれど。
- / ツイートする
- / 投稿日: 2004年07月12日
- / カテゴリー: [Papers_unclassified]
- / Edit(管理者用)
# mmmm
全般的にMaunsellの述べていることはもっともですが、reward or attention?という問いに意味があるのかどうかは疑問です。概念としてはattentionの方がreward expectationよりも上位だと思います。例えばMounsellはtask-difficultyとreward expectationとは逆相関するからそれらを分離することを提案してますが(それ自体は面白い実験であるし、Shidara & Richmond 2002やWalton et al. 2002はそれに関連したデータを示していると思いますが)、両者ともある種のattentionと見るのが妥当という気がします。passive attentionはともかく、active attentionの一般性を脳がどのように実装しているかというのは難しい問題で、reward expectationを含め、多くの詳細な研究の先にある問題だと思います。もちろんreward expectation意外のattentionという言い方はありで、そのうちのひとつがtask-difficultyとかeffortであったりするのではないでしょうか。
# pooneilありがとうございます。「Active attentionの一般性」、「Shidara & Richmond 2002やWalton et al. 2002」、このへんキーワードですね。もう少し読んどきます。それで、うーむ、Maunsellも挙げているように、Platt and Glimcherのreward probabilityにselectiveなLIPニューロンがじつはattentionによるものではないか、という問題はじっさい、このあいだのNewsome論文でも当てはまる話でして、ここはよくよく考えるべき問題ではないかと考えます。じつはこの間のjournal clubでもそういう指摘が出たのですが、Platt and Glimcherでとりあえず片はついていると思う、と言ってお茶を濁してしまいました。ふつうreward or attention?といった問題はより簡単な説明で済むほうで説明されるべきなのですが、attentionとrewardではどちらがより簡単な概念かとは言えないわけで、そこが難しい点なのだと思います。それが私が書いた、それらの概念が整理されていくことが必要である、と書いたことの動機でもあります。あと、mmmmさんが書いたことを展開すると、attentionというのは必ず何かほかのものに付随していて、reward expectationやtask difficultyやeffortというものがあるときにそれらに付随してくる、というふうに捉えられるかもしれません。この辺でattentionの心理学的研究についてもう少し知っておきたいところです。つまり、attentionがそれ以外のcognitiveなfactorとどのように関連付けられているか、といった点について。もしくはWilliam Jamesでも読んだ方がよいのかも。
2004年07月07日
■ Nature Neuroscience 7月号
- "Recalibration of audiovisual simultaneity." NTTの西田先生とCaltechの下條先生の名前が入っている。 私たちが目の前でボールがバウンドしているのを見るとき、バウンドする動きと地面に叩き付けられる音とが同じ物体(ボール)から来ていることを間違えることなく結び付けます。しかし、これはじつは不思議な話で、視覚情報が処理される脳の領域と聴覚情報が処理される脳の領域が違うだけではなくて、それらの処理されるのにかかる時間も違います。つまりそのような光と音との同時性(simultaneity)がどのようにして確立するか、という問題群があります。これはもちろん光と音だけの話ではなくて、視覚情報内でも動きと色との同時性がどう確立するか、という問題があり、そのような同時性が崩れるようなillusionがいろいろ考え出されています。Flash-lag illusionに関しても動きと光の点滅とのあいだでの同時性の問題であるといえます。ひとことで言ってしまえば、binding problem関連ですな。 そしてこの論文では、そのような視覚と聴覚との同時性の確立がその別々の時間に処理されたものを同時である、というふうに再構成している、という視点に立ちます。つまり、視覚の処理が行われた時間t1と聴覚の処理が行われた時間時間t2が経験上同時であったとき、この処理のずれt1-t2がゼロであるようにrecalibrationされている、という仮説を立てます。重要な点はこのrecalibrationがそれまでの経験によってcalibrateされているということです。よってわざとこの同時性をずらしたような刺激を使ってやってそれにadaptationさせるとこのrecalibrationが起こって同時性の基準がずれる、というのがこの論文でわかったことです。うーむ、面白い。そんなに重要なことが日常の経験から決められているとは。 この話は物体が近くにあるときの話なのだけど、もちろん光速と音速の違いによるcalibraionの問題というのもあることでしょう。
- "Automatic avoidance of obstacles is a dorsal stream function: evidence from optic ataxia." "The visual brain in action"をGoodaleと書いたDavid Milnerです。Parietal cortexにlesionのある患者さん二人へのテストの結果です。これについてはまた。
- "Deficits in saccade target selection after inactivation of superior colliculus." Edward L Keller @ The Smith-Kettlewell Eye Research Institute。これについてはまた。
- / ツイートする
- / 投稿日: 2004年07月07日
- / カテゴリー: [Papers_unclassified]
- / Edit(管理者用)
2004年05月26日
■ Tirin Moore
ヤバイ。超重要。
玉川のシンポジウムに来るってんで、Tirin Moore @ Princeton の論文をいくつか読んでいった。
Mooreのいちばん有名な論文は
Nature '03 "Selective gating of visual signals by microstimulation of frontal cortex."
であろう。FEFを微小電気刺激することで、刺激部位とretinotopicallyに対応したV4のニューロンの視覚応答が増大する。つまり、FEFでのsaccadeの指令シグナルが遠心性コピーとして視覚野に戻ってきて視覚処理をコントロールしていると考えられるのだ。また、これと関連するものとして、
PNAS '01 "Control of eye movements and spatial attention."およびそのfull paper versionの
JNP '04 "Microstimulation of the Frontal Eye Field and Its Effects on Covert Spatial Attention."
において、FEFを微小電気刺激することでサッケードに影響を及ぼすだけでなく、covert shift of attentionにも影響を及ぼすことを示した。これはRizzolattiらのいわゆるpremotor hypothesisというやつのsupportとしての役割を果たしている。つまり、サッケード(眼球を動かすことによる注意の移動:overt attention)と目を動かさずに注意だけを移動するcovert attentionとが共通のニューロンメカニズムを使っているというもので、これは進化上でサッケードがどのようにしてOKRなどから独立したか、さらに眼球の動きとは独立に注意を向けられるようになったか、ということを考察するに当たって非常に面白い。
話をMooreの方に戻すと、つまりFERFの段階でサッケードの指令を出していると思われる領域がcovert shift of attentionの制御にも関わっていることを示して、両者が共通のニューロン群を使って行われていることを示唆している。このへんのreviewはこちら:
Neuron '03 review "Visuomotor Origins of Covert Spatial Attention."
さらにその前にMooreは
Science '99 "Shape Representations and Visual Guidance of Saccadic Eye Movements."
においてV4でpresaccadic activityがあることを示している。つまり、このScienceと上記のFEF微小電気刺激とを組み合わせてNature '03を出したということを考えるといかに理路整然と、狙ったとおりにパーツがはまっていくかのように仕事が進められていることに感動を覚える。
で、このへんを読んだ上で話を聞きに行ったし、この辺についての話を聞いてきたのだが、この人にはまだまだ重要な仕事があった。つづく(あと二日分の原稿完成)。
- / ツイートする
- / 投稿日: 2004年05月26日
- / カテゴリー: [Papers_unclassified]
- / Edit(管理者用)
# antares
それはもしかして、この間も話題にのぼっていた、Neuron 2002のMicrostimulationのことでしょうか?それとももっとNewなもの?Mooreのプレゼンでどのような質疑応答があったか、ぜひ知りたいです。楽しみにしてます。
# pooneilそう、それです。でも、M1の話はシンポジウムではしゃべりませんでした。シンポジウムでは微小電気刺激のパラダイムに付いての質問があったと思いますが、聞き逃しました。シンポジウムの内容はいままでのまとめという感じで上記のNeuronのレビューの通りにFEF microstimulation関連の話をしていたので、新しい結果はなかったように思います。
んで、明日はM1についての話を書きます。StrickがNature NeuroscienceでMooreのM1論文を批判したことはキャッチしております。このバトルはStrickの勝ちではないかというのが私の印象ですが、どうでしょう?
私も同感です。的を得た指摘だなと思います。いずれにせよ、やはりずいぶん注目を集めた論文で、関係者の関心は高かったと思われます。
2004年05月21日
■ Nature
"Steady-state misbinding of colour and motion." 下條先生 @ Caltech。
シンポジウムに来てたが、話したネタはこれとは別もんだった。
視覚野は視覚の特徴、たとえば色、形、動き(たとえば赤、花、揺れる動き)などをそれぞれ別の領野で処理していると考えられている。では、そのようにしてばらばらに抽出された特徴をどうやって一つの視覚像に統合するか(たとえば、赤い花が揺れている)、これがbinding problemというものだ。今回の論文はそのようなbindingが崩れる例を提示している。
下へ動く赤の点と上へ動く緑の点がランダムに混ざった状態で動くランダムドットを見ていると、ある条件で、下へ動いているのは緑の点で上へ動いているのは赤の点であるというように逆転してしまう。つまり色と動きとの結びつき(binding)が崩れているのだ。
このようなbindingの崩れという例は今までにも見出されてきたが、今回の例はそのようなbindingの崩れが一時的に起こるようなもの(つまりそれゆえに注意がちゃんと向いていないことで説明される可能性がある)ではなく、安定して起こるという点が重要であって、たとえば今後、この実験パラダイムを使った神経メカニズムの解明などに活用できる可能性がある。
- / ツイートする
- / 投稿日: 2004年05月21日
- / カテゴリー: [Papers_unclassified]
- / Edit(管理者用)
2004年05月14日
■ Consolidation and retrieval
5/9の「モスバーガー」さんの質問からつづき。質問ありがとうございます。
質問1:「retrievalなしのconsolidationを調べるテストってあるんですか?」
答え1:私だってべつに専門家じゃありません(<-学問の分業化の弊害)が、どんなtaskでも単一のcognitiveな成分を見ているということはありえないわけで、(1) あるタスクの中でretrievalとconsolidationとそれぞれの成分を独立に操作できるようになんか条件を振ってperformanceを見る、(2)薬理、lesion study、neurology、geneticな方法を(1)に組み合わせる、(3) 電気生理やimagingなら時間の軸を使って分けられる、あたりの方策が一般的には取られていると思います。(1)のほうは心理学研究のほとんどで行われていることだと思うので、Experimental psychologyあたりの論文を探してみるとよいのではないでしょうか。もし分離できないようなときはそれらをまとめた対応物を操作的に定義してやるということになるでしょう。そのへんの操作的概念の扱い方が、episodic memoryやrecognition memoryや、familiarityとrecollection(RK judgement)などの概念を生んできたものであると考えております。(3)についてはもちろんご存知でしょう。
Encoding, consolidation, retrievalといったステージについてはCurrent Opinion in Neurobiology '01 "Molecular mechanisms of memory acquisition, consolidation and retrieval."あたりが取っ掛かりになるのではないでしょうか。ほかに今見つけたのは、Behavioural Brain Research "Intracranial self-stimulation facilitates memory consolidation, but not retrieval: its effects are more effective than increased training."がありました。この辺でなにやってるかわかったら是非知りたいのでまた投稿してください(強制しないっす)。
質問2:「学生の時に,研究者として将来自立するためにあったらよかった機関とか機会とかってありましたか」
答え2:べつにまだ自立していないですよ。自立というのは、自分のアイデアを、自分でお金を集めてやることが出来るところまで行かないといけないわけで。
で、あったらよかったものですが、やっぱり横のつながりによる情報というものが前から、今でもあった方がよかったと思います。移動の時にはそれなりに情報を集めてから新しい環境へ入っていったものですが、それでもやっぱり足りなかったし、もっといろいろわかっておくとよかったと今でも思います。私は研究者の流動性は高くあるべきであると考えて、恐れず新しい環境へ行こうと心がけておりますが、そのためにはもっといろんなことがわかってないとハマる、とけっこう切実に思います。
- / ツイートする
- / 投稿日: 2004年05月14日
- / カテゴリー: [Papers_unclassified]
- / Edit(管理者用)
# モスバーガー
お返事ありがとうございます。質問が悪くてすみません。Current Opinion in Neurobiology ’01を知らなくて質問したわけではなかったんですが,JNS 5/19のSilva&Kidoで疑問がちょびっと解けそうな気がするので読んでみます。日本語ってむづかしい。
# pooneil”Memory Reconsolidation and Extinction Have Distinct Temporal and Biochemical Signatures” http://www.jneurosci.org/cgi/content/full/24/20/4787 これですね。なんかわかったらぜひ書き込んでください。私が見たところ、reconsolidationやextinctionというような概念は主にratやmouseの研究で使われるようですので、この概念がhumanなどではどのように使われているのか、というあたりが取っ掛かりになる気がします。
2004年05月12日
■ Trends in Cognitive Sciences 五月号
"The brain circuitry of attention."
Stewart ShippはZekiとずっとやってきた視覚野の研究者。いちばん有名なのはV2からV4へ行く経路(ventral pathway)とV2からMTへ行く経路(dorsal pathway)とがV2の中ですでにsegregateしていることを示したNature '85 "Segregation of pathways leading from area V2 to areas V4 and V5 of macaque monkey visual cortex."であろう。
で、ShippはZekiから離れたあとにV2,V3,MTなどのextrastriate cortexとpulvinarとの間の投射の解剖学的論文を出した。(The Journal of Comparative Neurology '01 "Corticopulvinar connections of areas V5, V4, and V3 in the macaque monkey: A dual model of retinal and cortical topographies.")
今回のレビューはこのデータをもとにして、pulvinarを中心としたattentionシステムのモデルを作っている。かつてCrickが視床(とくにreticular nucleus)が大脳皮質の各領野と相互に結合を持ち、それらの情報を監視、統合できる可能性に注目して、視床がattentionのスポットライト的な役目を果たしているとする仮説をPNAS '84に出したことがあるのだが、今回のレビューはCrickのやつのアップデートバージョンであると言える。
で、いろいろあるのだが、けっきょくventral pulvinarがV1/V2/V4/TEO/TEといったventral pathwayへsaliency mapの情報を供給している。また、dorsal pulvinarやand mediodorsal nucleusからLIPやFEFへは情報が行っている。というわけで基本的にはextrageniculateのpathwayだけで話がつくようになっている。
大脳皮質の研究者は私を含めてどうしても大脳皮質だけで情報処理を考えすぎなのだが、response latencyなどのことを考えると、そんなに単純にfeedforwardでserialに情報が伝達されていると考えるだけでは、間違った方向へ進んでしまっているのかもしれないのだ。
- / ツイートする
- / 投稿日: 2004年05月12日
- / カテゴリー: [Papers_unclassified]
- / Edit(管理者用)
2004年05月09日
■ Science 5/7
"Independent Cellular Processes for Hippocampal Memory Consolidation and Reconsolidation."
Fear conditioningのタスクを使ってラットの海馬へのアンチセンスの効果を見た。Consolidation (短期記憶から長期記憶へ変換する過程)にはBDNFが関わっているし、そのような長期記憶が呼び出されることによってさらに強く固定されてゆくreconsolidationの過程にはZif268が関わっている。しかもこれはdouble dissociationであると。BDNFが宣言的記憶の長期記憶の形成に関わっていることはすでにわかっていたわけで(Nature Neuroscience '00 "BDNF upregulation during declarative memory formation in monkey inferior temporal cortex.")、重要なのはたぶん"reconsolidation"の方だろう。これってどんなもんなんだろう。たんなる時間差の問題である可能性を消しているかどうか、読んだ人教えてください。
- / ツイートする
- / 投稿日: 2004年05月09日
- / カテゴリー: [Papers_unclassified]
- / Edit(管理者用)
# モスバーガー
初心者の質問1.consolidationでもreconsolidationでもいいんですけど,保持していることを確認するテストってretrievalが常に伴っちゃうんじゃないかって思うんですけど(conditioningでいうextinction phase),retrievalなしのconsolidationを調べるテストってあるんですか? 質問2. 学生の時に,研究者として将来自立するためにあったらよかった機関とか機会とかってありましたか(あるいは今でもある)?
# pooneilどうもありがとうございます。長くなったので返事は5/14に書きました。D’Espositoはぜんぜん読んでないです。
2004年05月07日
■ BBS online
いつだったか忘れたけどMLにBehavioral and Brain Sciencesのtarget articleがアナウンスされていた。Arbibの"From Monkey-like Action Recognition to Human Language: An Evolutionary Framework for Neurolinguistics."(PDFファイル)だ。
というわけでまたもやmirror neuronと言語との関係についてなのだが、さて、どんなものだろうか。なんにしろ、p.45のfigure.6など見ていると楽しくなってくる。TptとPFextendedがWernicke野だそうな。DLPFとSTSaから視覚由来の情報をPFextendedがもらい、Broca野へ入力することで[sign languageの基礎]となり、聴覚情報はTptからBroca野へ入力することで[speechの基礎]となる。視覚情報はSTSa->PF->F5mirrorへ行って[行動の認知]に関わり、また、視覚情報はAIP->F5cannonicalへ行って[行動の選択]に関わる、本当かどうかは別としても、こういうのが回路っぽく書かれること自体が驚きであり、面白いところである。
こういうアプローチについて酒井さんは「言語の脳科学」で批判的に書いているわけだが、ま、この辺についてはもう少し勉強してからコメントする必要がある。
- / ツイートする
- / 投稿日: 2004年05月07日
- / カテゴリー: [Papers_unclassified]
- / Edit(管理者用)
2004年03月28日
■ Neuron 三月号
"Effects of Cholinergic Enhancement on Visual Stimulation, Spatial Attention, and Spatial Working Memory." Dolan @ Wellcome Department。
Physostigmineを投与して視覚応答や注意への影響をfMRIでみる。薬理のパラダイムをfMRIでやるのは難しい。コントロールと薬物応答とのあいだに時間の開きがあるため(after controlまで入れれば最低30分はかかるであろう)、BOLDシグナルのゆっくりしたドリフトと応答とのinteractionの可能性(もしシグナルが頭打ちになる可能性があるのならば、baselineが上がればevokedの大きさが小さくなるのはあたりまえだが、そのことからevokedをinhibitしているとは言えない)を考えなければならないからだ。だから、たんにドリフト成分を除去したあとでevent-relatedをやるだけでは私は不充分であると考える。また、このようなドリフトとbase lineに効いてくる薬理作用の成分とを分離するのも難しいと思う。で、読んでみるとplacebo/physostigmine と working memory task / control taskでのinteractionを見ているだけらしい。興味失った。
- / ツイートする
- / 投稿日: 2004年03月28日
- / カテゴリー: [Papers_unclassified]
- / Edit(管理者用)
2004年03月26日
■ Nature
"General conditions for predictivity in learning theory."
TOMASO POGGIO @ MIT。David Marrと一緒に仕事をした超有名人。3D objectのrecognitionのモデルとかがいちばん有名なのではなかろうか。News and viewsはこちら。
"Learning theory: Past performance and future results."
N&Vだけしか読んでないが、こういうことらしい。
Learningによってどう新しい状況に対して行動できるか、についてlearning theoryはrisk minimization algorithmを使っていると考えた。つまり、今までのすべての経験を使って、新しい状況において起こりうるリスクが最小になるように最適化されている、というものだ。しかし著者らはここで"stability"という概念を持ち出す。今までの経験のうちの一回の経験を除いても学習による予測が大きくは変わらないように最適化されている、というものだ。NVではケプラーの理論がそれまでの観察のうちのどれかが欠けたとしてもvalidであろうことを例にあげている。
このstabilityという概念は非常に尤もなように思われる。ベイズ統計を思い起こさせるところもあると思うが、私がここで思い浮かべたのは、統計でのcross validation法についてだ。いま私はkernel density estimationやgeneralized additive modelなどについていろいろ調べているのだが、カーブフィッティングをするときにkernelの幅やsmoothing splineの性質などを決める必要がある。このため、fittingの誤差のmasure(RMSとか)があるデータ点を除いたときにも変わらないという拘束条件のもとに、全データをそれぞれ一つずつ除きながらパラメータを最適化する。これが最適なものであるのかどうかは私は知らないが、empiricalにはよく使われている方法であるようだ。
というわけで、どっかで関係してこないかなと思って斜め読み。
- / ツイートする
- / 投稿日: 2004年03月26日
- / カテゴリー: [Papers_unclassified]
- / Edit(管理者用)
# pooneil
ガヤ、日記での指摘ありがとう。直しました。
で、コメントありがとう>estimationとprediction。調べてみた。予測のためのモデルをestimateする、そのモデルを使ってある変数をpredictする、ということのようだけど、うーん、正直よくわからなかったです。どっち(cross-validationも今回のstabilityのルールも)もpredictionしてると思ったから。Nature論文もcross-validationは道具として使っているようなので、問題は道具とアルゴリズムとの関係になるのかもしれない。とはいえ、論文を読んでみることが先決でしょうな。ということでツッコミまたよろしく。
私も今勉強中。最適化とか最尤度といったキーワードからなんとなく共通項を探そうとしてしまいがちですが、estimationとpredictionではやはり本質的に概念が違うかと。まずは私もそのNature論文読んでみます。
# ガヤすまん、あまりよく理解しないまま書き込んでしまいました。ラボ発表で『ガヤのやっていることは神経活動の「予測(prediction)」ではなくて単なる「推定(estimation)」ではないか』と指摘されまして今頭が混乱しています。そして”Applied Optimal Estimation (A. Gelb, MIT)”という読むように薦められたのですが知ってます、この本? 調べてみたら結構昔に出版された本のようでまだ購入に踏みきれません。有名な本なんでしょうか。
「(cross-validationも今回のstabilityのルールも)もpredictionしてる」→これはまさにその通りですよね、たぶん。
なるほど、そういう経緯ですか。うーむ、optimal ”estimation” だしなあ。上で私が言ったことも間違っているのかも。ここはプロに現れてほしいところですな。私自身は非線形最小二乗法を使おうとしてmatlabのoptimization toolboxを使ってみたぐらいでして、たぶん研究所の図書館の本でなんか調べたと思いますけど、お勧めできる本はありません。”Applied optimal estimation”は1974年らしいけど、よさそうですね。いまアマゾンで検索かけたら一番上に出てきました。
ネットでカルマンフィルタについての記述を発見。「なお、未来の状態ベクトルを推定することを予測、現在の状態ベクトルを推定することをフィルター、過去の状態ベクトルを推定することを平滑化と呼びます。」http://www.ichigobbs.net/cgi/readres.cgi?bo=economy&vi=0753&rm=20の423
おお、そしたら推定はすべてのケースを包括したタームということですね。情報どうもです。
# pooneilいやいや、ほんとのところはどうだかよくわからないし、上記の意味ではガヤの1sec先までを推定できるのはpredictionと言えるはずで、まだその質問の論点を見切れてはいなさそう。
2004年03月11日
■ JNS 4月号
- "Neural Correlates of Beauty" Zekiなんだけど、ガヤが言うように「脳は美をいかに感じるか」とあわせて読むモノなのだろうなあ、といちおうreferenceを確認してみたら、自著は入ってなかった。いいんだか、悪いんだか。内容は、絵画を見せてそれを美しいか醜いか分類させて、それらの絵を見たときの反応の差分を取る、というもの。いいのか? いろんな要素、たとえばemotionalな要素とかいろいろな可能性があると思うのだけれど。押さえとしては絵画を別の基準で分類したときにはどういう差分が出るかをやっていて、風景かそうでないか、とかをやってる。結果としては、美しいか醜いかの差ではorbitofrontalに活動の差が出るそうな。美しいと満たされて報われる、といったところだろうか?
- / ツイートする
- / 投稿日: 2004年03月11日
- / カテゴリー: [Papers_unclassified]
- / Edit(管理者用)
2004年03月03日
■ JNP
"Synaptic Noise Improves Detection of Subthreshold Signals in Hippocampal CA1 Neurons."がGouldさんのところで言及されている。Stochastic resonanceは前から興味があって、Fersterの論文を読んだときにちょっと調べたのだけれど、いまだにどういうことなのかわからない。たんにthresholdがあるかないかの問題か、というとなんかそういうものでもないらしいし。"Stochastic resonance without tuning."だっただろうか? なんにしろ、そのころ考えたんだけど、心理学的にもnear-thresholdの現象には効いてきそうな話だ。たとえば、psychophysicsの実験なんかでthresholdを決定するときとかにはニューロンレベルでもかなりノイジーな条件(SDTのふたつのgaussianが重なっているあたりをイメージしながら)なわけで、それによってdetectionがenhanceされるようなことでもあれば、かなり普遍的な現象のはず。そういうのは見たことがないのだけれど、もしそうだとしたらdetectabilityをモデル化するときにはどこかに入ってこないといけないはず。
- / ツイートする
- / 投稿日: 2004年03月03日
- / カテゴリー: [Papers_unclassified]
- / Edit(管理者用)
# ガヤ
Noise-induced orderはかつてテーマに関係して調べたことがあります(結局まだ役立っていません)。んで思うのですが、確率共振現象はもっと高次な非線形ダイミクスですから、thresholdingよりも、たとえばBoltzmann machineにおけるnoisyな自由度でシグナルを効率よく検出していく操作に近いイメージが私にはあります。もちろん無根拠です(汗)。これに比べるとFersterはずっと単純な考え方なので「確率共振」とは呼べないのではと思っています。
# GouldJNPの論文、言及有り難うございます。僕には正直ちんぷんかんぷんですが、確率共鳴という現象自体は以前にちょっとだけ知識を仕入れたことがありました。
# pooneil3/6のところに書きました。
2004年02月25日
■ Science
"Attention to Intention."
Richard E. Passingham @ University of Oxford と Patrick Haggard @ University College London。
またもHaggardが出てきて、回る時計の針を見て時間を報告させるタスクをやっている。Motor intentionの時間(ボタンを押そうとした時間)と実際のmotor actionの時間(実際にボタンを押し出した時間)とをそれぞれ報告したときとを比べている。12/31に採り上げたSiriguの論文と同じパラダイム。ただ、今回はmotor intentionにattentionがかけられているときと実際の運動の実行へattentionがかけられるときとを比べることで(実際にしている運動などの条件はそろっている)attentionがintentionに向けられているときにpreSMAがより活動することを見出している。せっかくこういうパラダイムにするならmotor actionへattentionを向けているときはどこがより活動するのか見つけて、二つの条件でのdouble dissociationを出すのが筋なのではないだろうか。たぶんそういうところはないんだろうが。
- / ツイートする
- / 投稿日: 2004年02月25日
- / カテゴリー: [Papers_unclassified]
- / Edit(管理者用)
# Correggio
こんにちは.ご無沙汰しています.今度,岡崎でお会いできるのが楽しみです.やっぱり,Siriguの論文は,intentionへのattentionがかからないって話しなんですね.内部モデルと結びつけるのは,どうなんでしょうかねえ.彼女の考え方はとっても面白いんですけど.ところで,Correggioの絵は大好きです.ParmaのDuomoのだまし絵みたいな3Dの天井画がとってもすばらしい.では
# pooneilご無沙汰しております。メールの返事しないままでしたがお会いできるのを楽しみにしております。アブスト読みました。ポスターは三日間貼りっぱなしなので、充分お話できる機会が見つけられるかと思います。問題は私がどのくらい勉強できるかでして。
で、なるほど、Siriguの方をそういうふうに読み直すことができるわけですね。Time perceptionの話はやはりかなりややこしいなあと思います。そうやってみると、ShadlenのNeuron ’03なんかもattentionとdicisionで読めないところがどのくらいあるか精読する必要があるのかもしれません。(まだ読んでませんが。)
これですね。3Dの天井画。http://www2t.biglobe.ne.jp/~provence/italia/visit/visit01/parma.html これを見たときAIPはどう活動するんでしょう。
写真ではわかりづらいですが,本物はもっとすごい。4隅に柱の飾りなど本物かと思っていたら,てっぺんは絵でした。AIPはともかく,CIPは元気でしょう。両眼視差でない立体の手がかりにもよく反応しますから。
# pooneilうわっ、さっそく付け焼刃なのがばれてしまいました(AIPとCIP)。
2004年02月04日
■ Cerebral Cortex
"Neural Responses during Interception of Real and Apparent Circularly Moving Stimuli in Motor Cortex and Area 7a."
Georgopoulos @ University of Minnesota。
Georgopoulosはかつて運動ベクトルの方向をM1ニューロンがベクトルの和としてpopulation codingしているのを提唱して、これが一時は支配的な説となっていた("Neuronal population coding of movement direction." Science '86)。現在はこれにはたしかいろいろ異論が出ている。たとえば12/16に言及したStephen ScottはNature '01 "Dissociation between hand motion and population vectors from neural activity in motor cortex"で反証を出している。
で、この論文はこの話とは直では多分まったく関係ないはずなのだが、特記事項としては円を描いて動く軌道についての反応をM1と7aから記録していることだ。読んでないんで目的はわからないけれど、この軌道とそれに対するreaction timeを計測するのはtime perceptionについて扱おうとしているのではないだろうか。前にも言及したSiriguの論文とかでparietal系の障害によって時計の針の動きを報告する課題に障害が出る。これとの関係があるのではないだろうか、とreferenceを見ると関係なさそう……めげずに……time perception解明への応用が可能になるのではないでしょうか。
- / ツイートする
- / 投稿日: 2004年02月04日
- / カテゴリー: [Papers_unclassified]
- / Edit(管理者用)
2004年01月13日
■ Neuron
Neuronal Correlates of a Perceptual Decision in Ventral Premotor Cortex
Romo @ Universidad Nacional Autónoma de México。
触覚で振動の周波数を弁別するタスクをずっとやっていて
Natureを二つ出している。
Somatosensory discrimination based on cortical microstimulation
Neuronal correlates of parametric working memory in the prefrontal cortex
データがあまりにきれいで、しかも振動数を発火頻度がlinearにコードしているという結果で、ちょっと信じがたい。
今回はこのタスクでPMvから記録したらしい。
- / ツイートする
- / 投稿日: 2004年01月13日
- / カテゴリー: [Papers_unclassified]
- / Edit(管理者用)
# Pooh
Romoの言うとおりならprefrontalもPMvもすべて触覚への道なんだけれどとりあえずsomatosensoryなのに本当にF5?F4じゃなくて?というあたりからツッコミ。周波数の差と主張している発火は、サルがどれだけ確信を持ってVisualなMotor planを持っているかというだけで説明できるし、たぶん計算部位はPMvじゃない。でf1刺激中はたぶん振動周波数に応じて筋緊張が変動するので意味のあるデータはf1-f2間でその後の運動に依存しない緑線の活動それは434の関連細胞活動、59のdelay活動のうち8つだけ。斜め読みでもイタいよ。Natureも少しは真面目にツッコめばいいのに。と書いてNeuronだと気がついた。Neuronのawake monkeyの審査もScienceと似たり寄ったりだし。何となく納得。
# pooneil(注記:movable typeへ移動する際にコメントの改行を編集しました。)
2003年12月28日
■ Neuron
Goal-Related Activity in V4 during Free Viewing Visual Search: Evidence for a Ventral Stream Visual Salience Map
Gallant @ UC Berkeley。
V4ニューロンが目を動かす直前からすでに目を動かしたら受容野に入るであろう刺激をコードしている、というもの。LIPにも同様なものがあって、ME Goldbergはsaliency mapをコードしている。これとの関係が興味ある。なお、V4はFEFと強い結合があり、FEFからV4へのtop-down controlというやつが効いている、というストーリーは1月のNature (selective gatingってやつ)とも共通している。
http://dx.doi.org/10.1038/nature01341
- / ツイートする
- / 投稿日: 2003年12月28日
- / カテゴリー: [Papers_unclassified]
- / Edit(管理者用)
2003年12月16日
■ The Journal of Comparative Neurology
Cytoarchitecture and cortical connections of the posterior cingulate and adjacent somatosensory fields in the rhesus monkey
Pandya @ Boston University School of Medicine。
posterior cingulate cortexのcytoarchitectureとconnectivity。
Pandyaは'70年代からcortexのcytoarchitectureとconnectivityの研究をやってきた。
まえはSTSとかparietal cortexとかdorsolateral frontal cortexとかを扱っていたけど、だんだん大脳の領野を探求していくうちについに一番手付かずだったcingulateまでやってきた、というかんじ。
その意味では99年にEJNに出たretrosplenial area 30のanatomyも同系統。
Architecture and connections of retrosplenial area 30 in the rhesus monkey (macaca mulatta)
- / ツイートする
- / 投稿日: 2003年12月16日
- / カテゴリー: [Papers_unclassified]
- / Edit(管理者用)
お勧めエントリ
- 細胞外電極はなにを見ているか(1) 20080727 (2) リニューアル版 20081107
- 総説 長期記憶の脳内メカニズム 20100909
- 駒場講義2013 「意識の科学的研究 - 盲視を起点に」20130626
- 駒場講義2012レジメ 意識と注意の脳内メカニズム(1) 注意 20121010 (2) 意識 20121011
- 視覚、注意、言語で3*2の背側、腹側経路説 20140119
- 脳科学辞典の項目書いた 「盲視」 20130407
- 脳科学辞典の項目書いた 「気づき」 20130228
- 脳科学辞典の項目書いた 「サリエンシー」 20121224
- 脳科学辞典の項目書いた 「マイクロサッケード」 20121227
- 盲視でおこる「なにかあるかんじ」 20110126
- DKL色空間についてまとめ 20090113
- 科学基礎論学会 秋の研究例会 ワークショップ「意識の神経科学と神経現象学」レジメ 20131102
- ギャラガー&ザハヴィ『現象学的な心』合評会レジメ 20130628
- Marrのrepresentationとprocessをベイトソン流に解釈する (1) 20100317 (2) 20100317
- 半側空間無視と同名半盲とは区別できるか?(1) 20080220 (2) 半側空間無視の原因部位は? 20080221
- MarrのVisionの最初と最後だけを読む 20071213