« 神経科学大会終了 | 最新のページに戻る | V1から上丘へ行く経路はなにをしているか 補足 »
■ V1から上丘へ行く経路はなにをしているか
今回もサーベイとJCを兼ねて。上丘への視覚入力にはretinaから直接来るretinotectal (retinocollicular)な経路と大脳視覚皮質を介してくる経路があります。V1からも来るし、V2からも来る。LGNからは来ないみたいだけど。Schillerの論文でJ Neurophysiol. 1974 Jan;37(1):181-94. "Response characteristics of single cells in the monkey superior colliculus following ablation or cooling of visual cortex."というのがあります。nhpのV1をcoolingもしくはlesionして急性で上丘の視覚応答を記録すると、SGSの応答はcoolingまたはlesionによって影響を受けないけれども、SGIの視覚応答は全くなくなる、というものです。つまり、SGSはretinaからの直接入力によってdriveされるけれども、SGIはV1経由の信号がないとvisual responseを引き起こすことができないのです。解剖学的にはV1のlayer5,6からSCへの投射があることが知られていますし、LIPやFEFを回ってくればいくらでも上丘へ行く経路はあります。ただ、麻酔下なんで、あんまり高次なところは回ってないんではないかと思うんですが。
ひきつづき彼らはJ Neurophysiol. 1976 Nov;39(6):1352-61. "Quantitative studies of single-cell properties in monkey striate cortex. IV. Corticotectal cells."にて、上丘(表面から0.5mmあたりと書いてあるので基本的にSGSからSoあたり)を刺激しながら、V1からunit記録をして、上丘の刺激によってantidromicに刺激されるunitを見つけます。こういうニューロンはV1のlayer 5,6にあって、antidromicの応答潜時は4.6ms。でもって、そういうニューロンの性質を調べると、complex cellで、orientation tuningはbroadで、direction selectivityがあって、spontaneous activityが高い。そういう意味ではSGSのニューロンの性質には似ていません。そこで彼らのdiscussionとしては、V1から上丘へ来る入力は「SGSの応答特性を作っている」というよりは、「SGS->SGIの情報の伝達をゲートしている」のではないか、と言っています。これは重要かつ面白い指摘です。
さて、このあとで彼らはJ Neurophysiol. 1979 Jul;42(4):1124-33. "Composition of geniculostriate input ot superior colliculus of the rhesus monkey."を発表します。これは上丘SGIからunit記録して、LGNのmagnolayerをlidocaineでblockすると応答が消えるけど、parvo layerをblockしても応答が消えない。どっちをblockしてもSGSの応答は消えない、という論文です。彦坂先生の仕事以降だとmuscimolになるのですが、この時代は局麻でblockしているのでpassing fiberもblockしてしまうという問題があります。ただ、SGSの応答は消えない、というのがcontrolで効いているので、だいたい妥当なのではないかと思うのですが。というわけで上丘はV1から受けている入力はmagno系だけだ、というわけです。色の情報とかはもらってない。
さて、この論文の意義を説明するためには視覚情報がRGC->LGN->V1->V2と進む情報伝達にはM,P,K経路の三つのチャネルがあるということの説明が必要です。図を作っておきました。
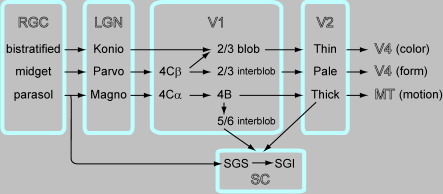
Schillerのだとcatの論文を引いて、X-type, Y-type, W-typeって話になるんでそこは翻訳しないといけないんですが、最新のレビューをもとにしてこれまでの知見を再構成してみました。図の元ネタはAnnu Rev Neurosci. 2005;28:303-26. "The circuitry of V1 and V2: integration of color, form, and motion." Sincich LC, Horton JC.および "The M, P, and K Pathways of the Primate Visual System" Ehud Kaplan in The Visual Neurosciences Ch.30です。図の表現上、一番上がkonio、真ん中がparvo、一番下をmagnoとしてあります。それぞれがcolor, form, motionのべつべつのチャネルとなってventral pathway, dorsal pathwayに行っていることはよく知られたことです。
それにさらに上丘への投射を組み込んでみました。LGNのparvocellular layerに終止しているRGCのX-type cell (midget cell)は上丘へは投射していません。LGNのmagnocellular layerに終止しているRGCのY-type cell (parasol cell)は上丘へ投射しています。あと、catではRGCにW-cellというのがあって、上丘に選択的に投射しているという話があります。(Klaus-Peter HoffmannによるBrain Res. 1972 Aug 25;43(2):610-6. "Very slow-conducting ganglion cells in the cat's retina: a major, new functional type?"およびJ Neurophysiol. 1973 May;36(3):409-24. "Conduction velocity in pathways from retina to superior colliculus in the cat: a correlation with receptive-field properties.") あと、conduction velocityが非常に遅い8m/secのはこのW-cellです。
ちょっと話が細かくなるのでここで改パラグラフ。CatのW cellがnhpになにに対応するかですが、koniocellularと考えた方がよいようです。Shapleyの80年代のTINSとかではそんな書き方をしています。そうすると、hnpではW-cellに対応するものは上丘へは投射していないと考えて方が良さそうです。ただ、Visual neurosciencesにもありましたが、W-cell、konioてのはどちらかというとmagno系、parvo系以外を全部入れたヘテロな集団と考えた方がよいので、これからの研究の進展によって話が変わってくる可能性はあります。なお、Schiller 1979では、W-like cellという言い方はintroでしているけど、それ以降ではまったく言及していません。また、それ以降の論文をしらべてみるとnhpでもW-like cellと表現しているのがあって、PNAS 2007 "Geniculocortical relay of blue-off signals in the primate visual system"とかではW-like = blue-off = koniocellularと書いてます。上丘にS coneからの入力が行っていないことはS cone isolating stimulusによる刺激による注意の効果の論文(Current biology)などから示唆されていますが、実験的には証明されていないようです。SGSからvisual response記録して、L-, M-, S-cone isolating stimulusで刺激するという実験をしたらいいと思います。きれいにS coneだけinsensitiveであることが示せたらJNSあたりいけるんではないでしょうか。
話を戻します。視覚皮質から上丘への入力はいくつかありますが、細かいことがわかっているのはV1,V2からのものです。
上丘にretrograde tracerを入れてV1で染まったニューロンの位置と、V1をCO染色してCOバンドとの位置とを比較したのがVis Neurosci. 1996 "The distribution of corticotectal projection neurons correlates with the interblob compartment in macaque striate cortex." Lia B, Olavarria JF.です。Layer 5/6のinterblobのニューロンが上丘へ投射しています。残念ながらinjectionは限局してないので、terminalはSGSなのかSGIなのかは不明。それ以前にもW. FriesがEBR 1985でlayer 5/6のMynert cellで上丘とMTの両方に投射しているものがあることを示しています(retrograde tracerによるdouble labellingを使用)。
同じグループが同様にしてV2で染まったニューロンの位置と、V2をCO染色してCOバンドとの位置とを比較したのがJ Comp Neurol. 1997 "Distribution of neurons projecting to the superior colliculus correlates with thick cytochrome oxidase stripes in macaque visual area V2." Abel PL, O'Brien BJ, Lia B, Olavarria JF.です。V2のthick band、つまりmagno系からおもに入力が上丘に来ています。
以上をまとめると図のようになるのでは、というわけです。
補足。ちなみに近年はganglion cellのレベルではなくて、coneのレベルでL+M-Sとかいったやり方で機能を描写するので昔の書き方を翻訳しないと使えません。このへんに関する最新の成果がEdward M. CallawayのNature 2003 "Parallel colour-opponent pathways to primary visual cortex"です。これを踏まえてconeからRGCまでの変換を書くと、parasol cellはcone全部の信号を足していて、L+M+Sで色に関わらない明るさをコードしていて、midget cellではL-Mでred/greenのシグナルを作っていて、bistratified cellではS-(L+M)でblue-onの受容野を持っています。そこでblue-offのニューロンを見つけてそれがV1のlayer 4Aに入力しているというのを見つけたのがNature 2003の仕事でした。これは図に組み込んでないけど、bistratifiedがblue-on cellでlayer 3Bあたりにメインで行ってて、それとはべつにRGCにblue-off cellがあって、これが4Aに終止していて、この二つを合わせてblue-on,offに関わるkoniocellular系を作っている、というのが最新バージョンではないでしょうか。
意外に充実してきたので今度のセミナーはこれにするかも。ちと専門的すぎるけど。
- / ツイートする
- / 投稿日: 2007年09月13日
- / カテゴリー: [上丘、FEFと眼球運動]
- / Edit(管理者用)