[カテゴリー別保管庫] 内側側頭葉と記憶システム
2013年03月10日
■ perirhinalでの刺激-刺激連合と刺激-報酬連合
2012/10/30
Science 2012 "Preference by Association"に関する話だけど、nhpではperirhinalには刺激-刺激連合記憶と刺激-報酬連合の両方がある。この両者をどう繋ぐかって問題になる。海馬関係はヒト、nhp、rodentsでいつも話が食い違う。
.@kosukesa なるほど「種による知識構造が違うのかも」これはあり得ますね。Empiricalな問題としても面白いところだとおもいます。
ただ、海馬周りに関しては連合学習にspecificな話でもないので、端的にrodentでは海馬に(電極で)アクセスしやすくて、monkeyではアクセスしにくい。ヒトではperirhinalは小さくて活動が見えにくい。このへんのサンプリングバイアスが話に混ざってる。
そういう意味では、最近のNature論文で出てきた、nhpでもentorhinal cortexにgrid cell見つかったってのは面白いと思う。
.@xmatumo 連合を形成している場所と、それをstoreしている場所が違うってこと? それはありうるとは思います。
.@kosukesa @xmatumo nhpでのlesion実験で、perirhinal cortexが刺激-刺激の連合記憶ニューロンの形成(連合記憶自体ではない)に重要であるという仕事があります。 PNAS1996
刺激セットAを学習した後で、lesionしてから刺激セットBを学習する。両方の連合記憶ニューロンが無くなる。視覚応答ニューロン自体は消えない。つまり、perirhinal cortexは形成にも保持にも関わる。
Perirhinalを両側性にlesionすれば、学習は出来なくなる。Gaffanの論文のどれか。もう10年近くこの分野から離れているので忘れてしまった。
.@kosukesa ここで見ているのはニューロンの活動だけなので、両側性lesionの結果を探してるんだけどすぐには見つからないです。
ラットではaquisitionだけに関わるって言ってる: Learn Mem. 2010
だんだん思い出してきた。まとめると、1) もともとnhpでは海馬+扁桃体lesionによって視覚連合記憶の形成が阻害されるという話があったのだけれども、それは海馬+扁桃体にアクセスするためにperirhinal cortexを損傷したせいだということが分かった。
2) Perirhinal cortexが記憶で、inferotemporal cortexが視覚で、二重乖離するという損傷実験 Buffalo et.al. 1999 (Squire陣営) Buckley et.al. 1997 (Gaffan陣営)で意見の統一したようだったが
3) Perirhinal cortexは視覚にも関わってるんだって証拠がいろいろ出てきた Annual Review of Neuroscience 2007 このあたりからフォローしてない。
Perirhinal cortexが刺激-報酬連合に関わるって話は Liu Z, Murray EA, Richmond BJ (2000) Nat Neurosci 3:1307–1315. この中で、線条体->Perirhinal cortexのドパミンが寄与してるってのを読んだときは正直信じなかった。(解剖学的に結合が弱いから)
@xmatumo さんが言うとおり、orbitofrontal cortexからの入力で考える方が尤もらしいと思う。鮫島さんの仕事にも関係するかもしれない。
- / ツイートする
- / 投稿日: 2013年03月10日
- / カテゴリー: [内側側頭葉と記憶システム]
- / Edit(管理者用)
2010年09月09日
■ 総説 長期記憶の脳内メカニズム
ずいぶん前の話になるのだけれど、大学院講義で「長期記憶」について担当したことがあって、そのときにneurologyについてかなり調べ物をしました。(「大学院講義「記憶の脳内機構」」のスレッド参照)
そのときはこれを再構成してweb講義でも作ろうとか言ってたんですけど、けっきょく放置してしまいました。今回別件で長期記憶の総説を書くことになったので、そのときのネタを元にまとめを作ることにしました。編集部の人に許可を得たので原稿を公開しときます。ここから:
はじめに
この項では長期記憶のうち、宣言的記憶の脳内メカニズムに関する知見をまとめる。宣言的記憶は大脳皮質の内側側頭葉記憶システムによって処理されていると考えられている。このシステムは海馬も含むので、海馬が関わっている事項についても言及する。
長期記憶の神経システム
1. 長期記憶の分類
長期記憶は宣言的記憶および手続き的記憶の二つに分類することができる(図1A)1)。
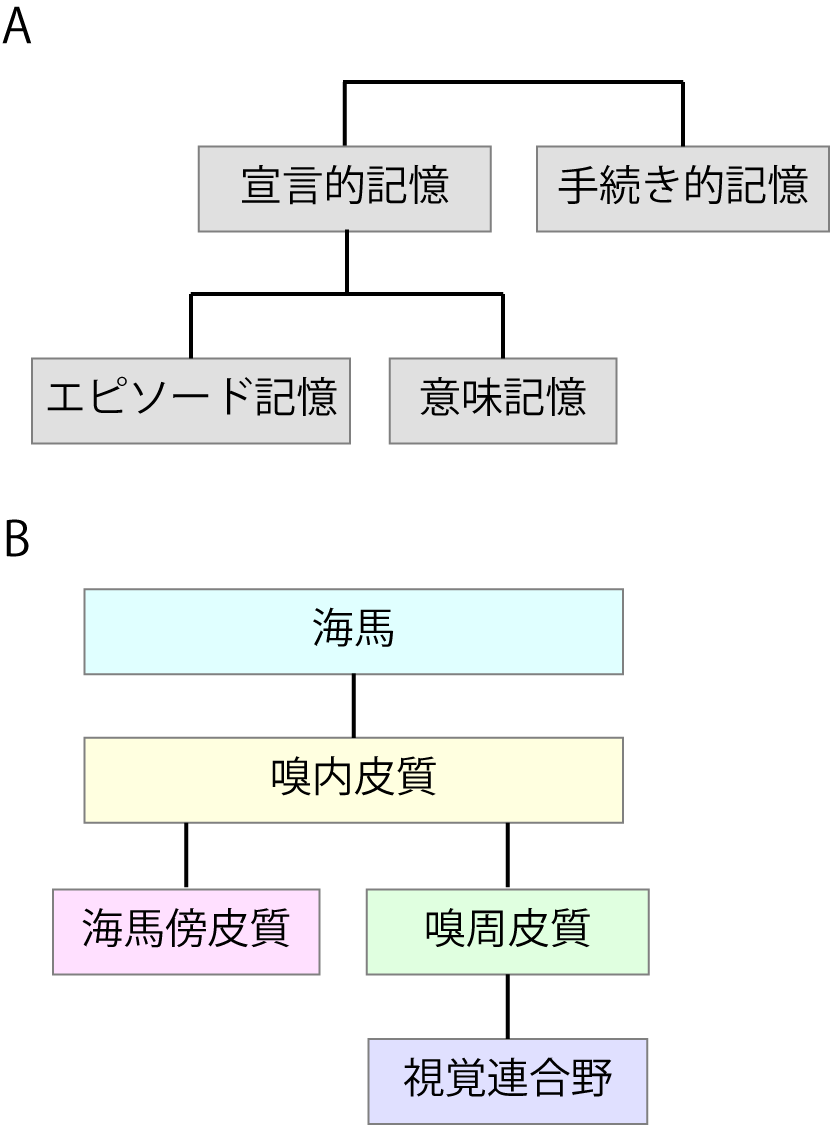
図1長期記憶のシステム。
A) 長期記憶の分類。
B) 長期記憶に関わる脳部位。
宣言的記憶は学習によって獲得された事実やデータに関する記憶のことを指す。「宣言的」というのはつまり「内容について言語で述べることができる」ことがその特徴であるということだ。一方で、手続き的記憶は学習された技能や認知的操作の変容にあたる記憶を指す。本項目では、大脳皮質で主に行われていると考えられている宣言的記憶について話を絞ることにする。
宣言的記憶はさらにエピソード記憶および意味記憶に分類することができる。Tulvingの定義2)では、エピソード記憶とは、出来事の時間的情報とそれらの出来事の時間的空間的関係を獲得して保持することである。一方で、意味記憶とは、ヒトが言葉や他の言語的シンボルの意味や指示対象に関して保持している組織だった知識のことである。
ざっくりとこの二つの概念を説明するなら、「911の事件のとき、あなたはどこで何をしていましたか?」がエピソード記憶のテストであり、「911の事件のときの日本の首相はだれですか?」が意味記憶のテストとなる。
2. 長期記憶に関わる脳部位
これまでの記憶障害の患者や動物使った脳損傷の実験などから、宣言的記憶は内側側頭葉の記憶システムによって支えられているということが明らかになっている。
図1Bは内側側頭葉の各部位とその結合を示している。このシステムの中心となるのは海馬だが、海馬への入力として、嗅内皮質があり、さらに嗅周皮質および海馬傍皮質がある。これらの領域はさまざまな感覚属性からの入力を受けているが、視覚情報に関しては、隣接する下部側頭葉から神経投射がある。
両側の海馬および隣接する皮質の摘出手術を受けた患者H.M.では重篤な宣言的記憶の障害が起こった3)。またより広範な両側性の損傷をもつE.P. (海馬、嗅内皮質、嗅周皮質を含む)の研究からもH.M.と同様な記憶障害を示すことが明らかになった4)。つまり、逆行性健忘によって、損傷前数年のエピソードおよび事実に関する記憶が失われていた。エピソード記憶、意味記憶の両方が影響を受けることから、内側側頭葉が宣言的記憶に関わることが明らかになった。
それでは、エピソード記憶と意味記憶とは関連する脳部位が違うだろうか。両側性の海馬選択的な損傷によってエピソード記憶の障害はあったが意味記憶は保持されていた症例がある5)。この症例では、たとえば、さっき聞いた話を憶えていることができない。しかし、言葉を憶えてしゃべることはできるし、学校に通って授業を受けていた。
逆に、意味記憶の選択的障害としては意味性痴呆がある。この症例では言葉や物の意味がわからなくなる。物体の名前が言えなくなる。つまり意味的記憶の障害がある。いっぽうで、エピソード記憶は比較的保持される。この障害は嗅内皮質や嗅周皮質を含む側頭葉前方部の萎縮によって引き起こされる。海馬の損傷は比較的少ない6)。このような知見を元にすると、海馬がエピソード記憶、嗅内皮質や嗅周皮質が意味記憶という機能局在があると考えることができるが、まだ論争は決着していない。
長期記憶の脳内メカニズム
3. 知識に関する記憶の情報表現
内側側頭葉において宣言的記憶がどのように実現しているかを明らかにするためには、ニューロンのレベルでの情報表現を調べる必要がある。このために有効なのは微小電極によって実験動物の神経細胞の活動電位を捉える方法だ。ここではサル内側側頭葉から単一ニューロン活動記録を行うことで得られた知見を紹介する。
動物を被験者として用いる研究では、脳内に電極を刺入することができるというメリットと引き替えに、ヒトでの記憶テストのように言語を用いることができない。このため、動物に合わせて記憶テストを改変する必要がある。サルは視覚情報処理の能力がよく発達しているので、記憶課題としては図形を用いた対連合記憶課題を採用した。この課題は、2枚1組の図形対の一方を提示し、もう一方の図形がなんであったかを答えさせる課題である(図2A)。百人一首をやるときに上の句と下の句の組み合わせを憶えるのと基本的に同じ頭の使い方をすることになる。つまり図形の対の関係という知識を長期記憶として獲得することが必要になる。よってこのテストはエピソード記憶ではなくて意味記憶のテストと考えることができる 。
この課題を行っている最中のサルの嗅周皮質から単一ニューロン活動を記録する。すると、12対で合計24枚の図形のうち対になっている図形3および図形3’それぞれを見たときだけ活動して、ほかの図形に対しては活動しないニューロンが見つかった(図2B)7)。つまりこのニューロンは図形3と図形3’とが対になっているという情報を表現している。つまり、個々のニューロンのレベルで長期記憶の情報表現が行われているということが明らかになったのだ。
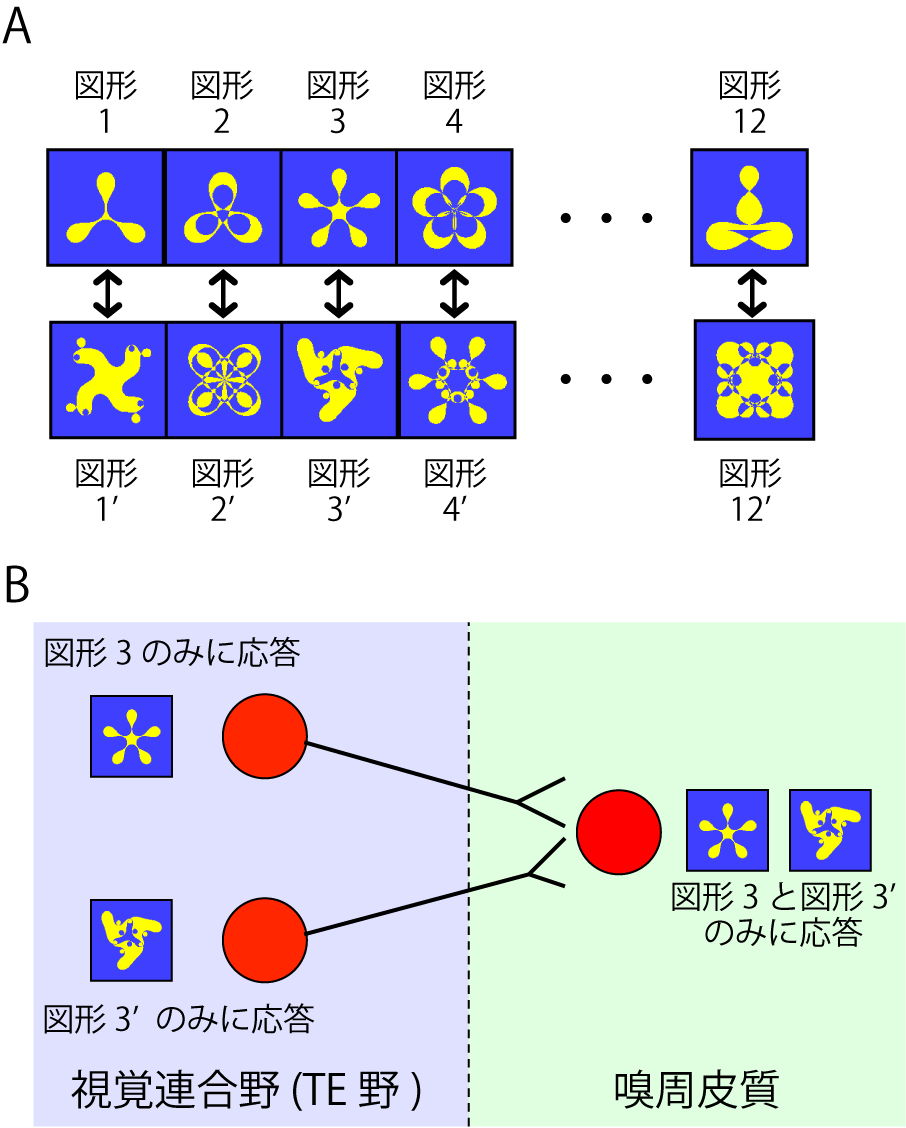
図2 対連合記憶課題
A) 使用した図形対。
B) 長期記憶の情報表現のメカニズム。
ではこのような情報表現はどのようにして形成されるか。嗅周皮質は隣接する視覚連合野(TE野)から視覚情報を受けている。このTE野から同様にニューロン活動を記録してみると、こちらはより視覚情報処理に関わっていることが明らかになった8)。つまり、図形3だけに応答するニューロン、図形3’だけに応答するニューロンが見つかってきた(図2B)。そしてこれらのニューロン群のあいだに線維結合があることも確認された9)。また、類似した研究では、このような長期記憶を獲得中の嗅周皮質を記録して、徐々に長期記憶の情報表現が形成されることが明らかになった10)。
このような状況証拠からすると、嗅周皮質の長期記憶ニューロンはTE野の視覚ニューロンの情報を連合することによって形成されるのだろう(図2B)。神経回路モデルではヘブ則(他の項参照)によってこのような情報表現を作ることが可能だ。
この一連の研究ではさらに、保持していた記憶を思い出す過程では、嗅周皮質からTE野に向かって情報が伝播していること11)や前頭葉からのシグナルによって内側側頭葉記憶システムが活動すること12) 13)などがあきらかになっている。
4. エピソード記憶の脳内メカニズム
それではエピソード記憶に関してはどれだけ明らかになっているかというとじつはまだ充分解明が進んでいるとは言えない。前述の「知識に関する記憶」と比べるとまだ個々のニューロンの活動のレベルでは解明されていない。ここからは未解決問題を含んだ研究の現状についてまとめる。
4-1. エピソード「様」記憶
Tulvingによればエピソード記憶とは「出来事の時間的情報とそれらの出来事の時間的空間的関係を獲得して保持すること」だった。Nicola Claytonはこの定義に基づいて、エピソード記憶を動物実験によって解明するために、行動を用いた基準を作った14)。つまり、もしある動物が「なにがどこでいつ起こったか」(what-where-when)を保持することができたとしたらそれは人間のエピソード記憶に近いものがあると言えるのではないか。Claytonはこの定義を人間のエピソード記憶と分けて扱うためにエピソード「様」記憶 (episodic-like memory)と呼んだ。それでは人間以外の動物にはエピソード様記憶はあるのだろうか?
Claytonが実験に使ったのはアメリカカケスという鳥だが、この鳥は食べ物を砂の中に隠す習性を持っている。実験でアメリカカケスはピーナツとイモムシを与えられると、それを砂の中に隠す。時間をおいてまたその砂を掘り返すチャンスが与えられる。ピーナツは日持ちするけど、イモムシは5日後には腐っている。もしアメリカカケスがイモムシをいつ、どこに埋めたかを憶えているならば、5日前にイモムシを埋めた場所は掘り返さないけど、4時間前にイモムシを埋めた場所は掘り返すはずだ。そして実際その通りになった。つまりアメリカカケスはwhat (イモムシ/ピーナッツ) -where (砂のどこか)-when (5日前/4時間前)の情報を保持することができた。つまりエピソード様記憶があるのだ14)。同様な実験はラットでも行われ、ラットもwhat-where-whenの情報を保持できることが報告されている15)。
しかしこの実験課題で本当にwhenの情報を保持していると言えるだろうか? 5日前かどうかを憶えていることと、その日がX月X日であることを憶えていることとは別だ。このような視点から批判論文も出ている16)。
ところでこの課題を行っているときにニューロンはどのように活動しているのだろうか? そのような研究はまだない。What-where-whenがニューロンのレベルでどのような情報表現をしているのかを明らかにするためには単一ニューロン記録の実験が必要だ。
4.2. エピソード記憶の定義再訪
上記のエピソード記憶の定義はTulvingが1972年に提出したものだが、それ以降もTulvingはエピソード記憶の定義についてなんどか更新を加えてきた。新しい定義ではエピソード記憶とは「私的に経験された出来事に関する記憶」であり、“mental time travel”、つまり過去に遡って自分のかつての経験を追体験することである17)。なるほど、たしかにこの表現は我々がふだん「記憶」という言葉でイメージするものをうまく言い当てているように思う。
しかしこのエピソード記憶の定義は意識的経験の存在を前提としている。動物に意識があるかどうかどうやって行動で確認できるだろうか? つまりClaytonが行ったような方法で、エピソード記憶を行動のみを指標とした操作的定義によって捉えることはできない。また、エピソード記憶とはある単一の出来事に関する記憶であって、実験室の条件で繰り返しテストを行うのにはそぐわない。じっさい、ある出来事にだけ活動するニューロンがあったとして、それを確かめるのはどうすればよいのだろう? ある出来事が繰り返されるならばそれはもう出来事ではなくて、実験条件の一つでしかない。このへんにエピソード記憶の脳内メカニズムを明らかにする際の本質的な難しさがある。
4-3. 「想起」と「親近性」の区別
すこし違った角度からの研究が進んでいる。宣言的記憶のサブカテゴリーとして再認記憶というものがある。再認記憶課題では、文字カードなどを提示して、一定時間経ったあとでまたカードを提示してそれが以前提示されたものかどうかを答える。この課題を解く際に被験者の内観としては「そのカードを憶えたときの記憶がよみがえる」という「想起」をしている場合と、「想起の感覚はないけれど、そのカードに見覚えがある」という「親近性」の感覚がある場合とがある。この二つのうち、「想起」の要素がエピソード記憶に対応していると考えることができる。
ヒト患者を被験者とした再認記憶課題で、想起と親近性について報告してもらった報告がある。海馬のみを損傷した患者では想起の要素のみ成績が低下したが、海馬と嗅周皮質などの周辺皮質を損傷した患者では想起だけでなく親近性の要素でも成績が低下した18)。この結果からYonelinasは海馬がエピソード記憶的な想起に、嗅周皮質が親近性に関わっていると結論づけた。
この研究は大きな反響を巻き起こし、反論を含む多くの報告が続いた19)。その結果現在では、想起と親近性に関わる脳部位は当初主張されていたほどに厳密に分かれているわけではないと考えられている20)。
この方法をラットの再認課題で応用した例もある。とはいえ動物に想起と親近性の内観報告をさせることはできないので、行動データに対してモデルを当てはめて間接的に予測することになるのだが、海馬を除去したラットでは、再認課題のデータから、想起に関連するスコアだけが低下することを示している21)。
むすび
長期記憶の脳内メカニズムは現在も新しい発見が続いているホットな領域である。経験の継続としての長期記憶は、私たちがひとつの自己を形成する際に重要な意味を持っている。長期記憶のメカニズムの解明は私たちの心の解明だ。
文献
- 1) Larry R. Squire著、河内十郎訳「記憶と脳」医学書院、1989
- 2) Tulving, E. (1972). Episodic and semantic memory. In E. Tulving & W. Donaldson (Eds.), Organization of memory, (pp. 381–403). New York: Academic Press.
- 3) Corkin S. What's new with the amnesic patient H.M.? Nat Rev Neurosci. 2002 ; 3 : 153-60.
- 4) Stefanacci L, Buffalo EA, Schmolck H, Squire LR. Profound amnesia after damage to the medial temporal lobe: A neuroanatomical and neuropsychological profile of patient E. P. J Neurosci. 2000 ; 20 : 7024-36.
- 5) Vargha-Khadem F, Gadian DG, Watkins KE, Connelly A, Van Paesschen W, Mishkin M. Differential effects of early hippocampal pathology on episodic and semantic memory. Science. 1997 ; 277 : 376-80
- 6) Hodges JR, Graham KS. Episodic memory: insights from semantic dementia. Philos Trans R Soc Lond B Biol Sci. 2001 ; 356 : 1423-34.
- 7) Sakai K, Miyashita Y. Neural organization for the long-term memory of paired associates. Nature. 1991 ; 354 : 152-5.
- 8) Naya Y, Yoshida M, Miyashita Y. Forward processing of long-term associative memory in monkey inferotemporal cortex. J Neurosci. 2003 ; 23 : 2861-71.
- 9) Yoshida M, Naya Y, Miyashita Y. Anatomical organization of forward fiber projections from area TE to perirhinal neurons representing visual long-term memory in monkeys. Proc Natl Acad Sci U S A. 2003 ; 100 : 4257-62.
- 10) Messinger A, Squire LR, Zola SM, Albright TD. Neuronal representations of stimulus associations develop in the temporal lobe during learning. Proc Natl Acad Sci U S A. 2001 ; 98 : 12239-44.
- 11) Naya Y, Yoshida M, Miyashita Y. Backward spreading of memory-retrieval signal in the primate temporal cortex. Science. 2001 ; 291 : 661-4.
- 12) Hasegawa I, Fukushima T, Ihara T, Miyashita Y. Callosal window between prefrontal cortices: cognitive interaction to retrieve long-term memory. Science. 1998 ; 281 : 814-8.
- 13) Tomita H, Ohbayashi M, Nakahara K, Hasegawa I, Miyashita Y. Top-down signal from prefrontal cortex in executive control of memory retrieval. Nature. 1999 ; 401 : 699-703.
- 14) Clayton NS, Dickinson A. Episodic-like memory during cache recovery by scrub jays. Nature. 1998 ; 395 : 272-4.
- 15) Babb SJ, Crystal JD. Episodic-like memory in the rat. Curr Biol. 2006 ; 16 : 1317-21.
- 16) Roberts WA, Feeney MC, Macpherson K, Petter M, McMillan N, Musolino E. Episodic-like memory in rats: is it based on when or how long ago? Science. 2008 ; 320 : 113-5.
- 17) Tulving E. Episodic memory: from mind to brain. Annu Rev Psychol. 2002 ; 53 : 1-25.
- 18) Yonelinas AP, Kroll NE, Quamme JR, Lazzara MM, Sauvé MJ, Widaman KF, Knight RT. Effects of extensive temporal lobe damage or mild hypoxia on recollection and familiarity. Nat Neurosci. 2002 ; 5 : 1236-41.
- 19) Squire LR, Wixted JT, Clark RE. Recognition memory and the medial temporal lobe: a new perspective. Nat Rev Neurosci. 2007 ; 8 : 872-83.
- 20) Diana RA, Yonelinas AP, Ranganath C. Imaging recollection and familiarity in the medial temporal lobe: a three-component model. Trends Cogn Sci. 2007 ; 11 : 379-86.
- 21) Fortin NJ, Wright SP, Eichenbaum H. Recollection-like memory retrieval in rats is dependent on the hippocampus. Nature. 2004 ; 431 : 188-91.
われながら、「長期記憶のメカニズムの解明は私たちの心の解明だ。」とかちょっと飛ばしすぎな気はする。だがそれがよい。
追記20110202 依頼元で原稿が出版されました。「月刊 臨床神経科学 Clinical Neuroscience」中外医学社 2011年2月号 特集「記憶のメカニズムとその障害」のなかの「大脳皮質 - 長期記憶」です。
- / ツイートする
- / 投稿日: 2010年09月09日
- / カテゴリー: [内側側頭葉と記憶システム]
- / Edit(管理者用)
2008年12月22日
2007年04月23日
■ Perirhinal cortex(嗅周皮質)の構造と機能つづき
さてさて、つづきで機能についてですが、non-human primatesとrodentsとでのperirhinal cortexのfunctionを比較して議論するレビューというのはなかったような。だいたい、rodentsでは海馬至上主義が蔓延していて、海馬内でDG-CA3-CA1で分担してencodingからrecallまでやってしまうようなストーリーになってたりするのであれでいいのだろうか、と思うことがあるのですが。ま、このへんは種差の問題もあって簡単ではない。Non-human primatesでは伝統的に視覚経路についての研究が進んでいるので、視覚入力がどのように処理されて、内側側頭葉記憶システムに入力してくるか、というような視点を持っているのが強みです。Rodentsだったらたぶん、視覚入力よりは違うものを使った方がよいのかもしれない。最近のRGM Morrisの話とか、Eichenbaumの話とかはだいたいolfactory系の刺激で砂を掘らせるとかそういう方向に行ってる。あのラインで感覚処理系と記憶系とを繋げてゆくというのがいちばん将来性があるのではないか、というのが私の持論です。うお、脱線。でも脱線が面白いもので。(このフレーズ使うのは何回目だろう。)
なんてエラソーなこと書いてて久しぶりにPubmed検索してみたら、MurrayのAnnual Reviewというのが出てますね。これが「non-human primatesとrodentsとでのperirhinal cortexのfunctionを比較して議論するレビュー」というのではいちばん良さそうです。
- Annual Review of Neuroscience Vol. 30 (Volume publication date July 2007) "Visual Perception and Memory: A New View of Medial Temporal Lobe Function in Primates and Rodents" Elisabeth A. Murray, Timothy J. Bussey and Lisa M. Saksida
ただ、このレビューはMurrayが近年主張している、perirhinal cortexはmemoryだけでなく、percetual aspectにも関わっているのだという話に偏っている傾向があります。そういう意味ではnon-human primatesだけになりますが、
- Trends in Cognitive Sciences Volume 3, Issue 4 , 1 April 1999, Pages 142-151 "Perceptual-mnemonic functions of the perirhinal cortex" Elisabeth A. Murray and Timothy J. Bussey
こちらの論文のほうがperirhinal cortexのmemoryに関する機能についてレビューするという機能をよく果たしていると思います。(上記の論文を間違えていたので訂正しました。ご指摘ありがとうございます。)
なお、Elisabeth A. MurrayはMishkinの弟子です。その昔、海馬のlesionよりも扁桃体のlesionのほうがrecognition memory testの成績が落ちるなんて話があったのですが、けっきょくそれは扁桃体のlesionでは扁桃体にアプローチするときにperirhinal cortexを損傷させていたからだなんてオチがあったわけです。そのへんの仕事をしてたのがMurray EA and Mishkin Mでした。
このへんで力尽きました。
- / ツイートする
- / 投稿日: 2007年04月23日
- / カテゴリー: [内側側頭葉と記憶システム]
- / Edit(管理者用)
# NORI
pooneilさん、こんにちは。私もperirhinal cortexに興味を持っているので、とても楽しく読ませてもらっています。ところで、ここで紹介しているMurrayたちの論文は同じ物なのですが、それでいいのでしょうか?
# pooneilご指摘どうもありがとうございます。間違えてたので直しました。
ところで、ratとmonkeyとでのperirhinalの相同性ってどのくらい明らかになっているのでしょうか。言及したBurwell 1995とかだとニッスル、Timm's stain、AChEを並べて議論しているようですが、いまだったらSMI-32とかもっといろんなマーカーでのimmunohistochemistryを頼りに出来そうです。Richmondの話であった、dopamine系の投射も重要ですし、いろんなものの発現プロファイルがこの問題に大きく貢献できるのではないかと思ってます。
また、私はどうしてもventral visual pathwayからの入力に注目しがちですが、rodentの海馬がspatial mappingに大きく関わっているという点で、dorsal pathwayからの入力がどういう経路を介しているかにも興味があります。Non-human primatesだったら、parietal-retrosplenial-parahippocampal-hippocampusという流れが確立しているように思えますが、rodentsだったらより入力が強烈なのではないか、とか考えたり。
それではまた。
2007年04月21日
■ Perirhinal cortex(嗅周皮質)の構造と機能
とおおげさなタイトルで始めてしまいましたが、non-human primatesとrodentsとでのperirhinal cortexのanatomyとfunctionについてということで関連するレビューをまとめてみました。しばらくこの分野はご無沙汰なので情報が古いかも知れません。お気づきの方はご指摘を。
まずは解剖学ということで。Perirhinal cortexは側頭皮質と隣接したarea 36と、より内側でentorhinal cortexと隣接したarea 35とからなっています。Area 36はlayer 4があるけど、area 35はlayer 4がないなどいくつかの点で構造が異なっています。しかし、どちらも側頭皮質からの視覚入力やその他のmodalityの感覚入力を受け、entorhinal cortexへ投射しているという点では共通しています。
Cytoarchitectureの違いに基づいた領野の区分についてのレビュー。
- Hippocampus (1995) Volume 5, Issue 5 , Pages 390 - 408 "Perirhinal and postrhinal cortices of the rat: A review of the neuroanatomical literature and comparison with findings from the monkey brain" Rebecca D. Burwell, Menno P. Witter, David G. Amaral
1995年とやや古くなっていますが、領野の区分および他領野との結合に関してratを中心にして、macaque monkeyと比較してあります。なお、perirhinal cortexのanatomyに関してはmonkeyもratも、UC DavisのDavid G. Amaral(Squire系列)のところで出しているものが現在のスタンダードになっていると言えます。
Monkeyに関してはさらに細かいsubdivisionが提唱されているのですが、
- The Journal of Comparative Neurology (2003) Volume 463, Issue 1, Pages 67-91 "Perirhinal and parahippocampal cortices of the macaque monkey: Cytoarchitectonic and chemoarchitectonic organization" Wendy A. Suzuki, David G. Amaral
ちょっとこれは話が細かすぎるかもしれないので、
- Neuroscience Volume 120, Issue 4 , 15 September 2003, Pages 893-906 "Where are the perirhinal and parahippocampal cortices? a historical overview of the nomenclature and boundaries applied to the primate medial temporal lobe" W. A. Suzuki and D. G. Amaral
こちらの方が領野の区分については歴史的経緯を追っていてわかりやすいかもしれません。時代ごとにみんな違った名称を使っていて、古い論文を読むとTE1/2/3(Pandyaによる分類)とか出てきて面食らうのですが、そういうときに便利。Humanとmonkeyの比較の資料としても役に立ちます。そっちはそっちで重要なポイントですが、以前の大学院講義スレで多少触れてます。(Humanでのsemantic dementiaが起こるところがperirhinal cortexに相当するのでは、とかそういう話。)
解剖学のつづきで他の領野とのconnectivityについて。大脳皮質連合野との相互結合、海馬やentorhinal cortexなどの内側側頭葉記憶システムとの相互結合それぞれを押さえる必要があります。だいたい双方向性なのですが、細かいことを言うといろいろある。んでもって、解剖学者は話が細かい。わたしを含めてのことだけど。Rodentsでのconnectivityに関しては、
- The Journal of Comparative Neurology (1998) Volume 398, Issue 2 , Pages 179 - 205 "Cortical afferents of the perirhinal, postrhinal, and entorhinal cortices of the rat" Rebecca D. Burwell, David G. Amaral
この論文のfig.14がいちばんよいまとめではないでしょうか。これをみると、area 36とarea 35とでやや違いがあるような書き方をしてます。Area 36のほうが側頭皮質との結合が強くて、area 35のほうがentorhinal cortexとの結合が強いとのこと。
Monkeyでのconnectivityに関しては、
- J Comp Neurol. 1994 Dec 22;350(4):497-533. "Perirhinal and parahippocampal cortices of the macaque monkey: cortical afferents." Suzuki WA, Amaral DG.
これの最後の図がいちばん良いまとめだと思います。かつてわたしはこの図を机の前に貼ってました。(と言っても私の机は荷物置き場になっていて機能してなかったのだけれど。)
うお、構造の方だけでいっぱいいっぱいなので、機能については次回。
- / ツイートする
- / 投稿日: 2007年04月21日
- / カテゴリー: [内側側頭葉と記憶システム]
- / Edit(管理者用)
2005年08月02日
■ Nature 7/28
"Robust habit learning in the absence of awareness and independent of the medial temporal lobe" Larry R. Squire
Medial temporal lobeを両側で損傷している患者E.P.さんと患者G.P.さんはdeclarative memoryを保持することが出来ないにもかかわらず、concurrent discrimination taskを繰り返すことで成績がよくなってゆく。
患者E.P.さんに関してはこれまでもSquireグループが研究してきました。以前の「記憶の脳内機構」スレッドにも関連記事があります。
また、しげさんのところの20050728のエントリに詳しい記載がありますので、そちらへどうぞ。
- / ツイートする
- / 投稿日: 2005年08月02日
- / カテゴリー: [内側側頭葉と記憶システム]
- / Edit(管理者用)
2004年07月02日
■ JNS 6/30
"Double Dissociation between the Effects of Peri-Postrhinal Cortex and Hippocampal Lesions on Tests of Object Recognition and Spatial Memory: Heterogeneity of Function within the Temporal Lobe." Timothy J. Bussey以前からrecognition memoryをラットで研究することあたりの話をしましたが、これも関係しそうです。Perirhinalのlesionとhippocampusのlesionとで機能のdouble dissociationがあることについてはMishkinの時代からAmaralやらGaffanやMurrayなどがさかんにやってきているわけですが、それをratの系に持ってきた、ということのようです。これはもちろんラットでやることのメリットがいろいろあるわけでして(ほんとうはマウスの方がもっとよいでしょうけど)、可能であるならより単純な系でanalogousなものを見れるようにしたい、という意味で評価できると思います。内容は読んでないけど、perirhinalがobject recognition、hippocampusがspatial memory、というのはあたりまえのようだが重要です。こんどはBuckley,Gaffan and MurrayのJNP '97 "Functional double dissociation between two inferior temporal cortical areas: perirhinal cortex versus middle temporal gyrus."を再現するようなITとperirhinalの比較に向かったら面白いじゃないでしょうか。Perirhinalはただのobject recognitionじゃないだろうし。
- / ツイートする
- / 投稿日: 2004年07月02日
- / カテゴリー: [内側側頭葉と記憶システム]
- / Edit(管理者用)
2004年04月09日
■ JNS 2/25
"Impaired Recency Judgments and Intact Novelty Judgments after Fornix Transection in Monkeys." David Gaffan @ Oxford。
3/4に貼っといただけのやつだが、これは重要だった。Gaffanはずっと海馬周りのlesion studyをやってきた。再認記憶にはいろんなコンポーネントがあるわけだが、この論文で彼らはRecency(最近見たことがあるか前に見たことがあるか)とNovelty(見たことがあるかないか)とが機能的に分離できることを示している。Fornix lesionではnoveltyだけ落ちてrecencyは落ちないそうだ。
- / ツイートする
- / 投稿日: 2004年04月09日
- / カテゴリー: [内側側頭葉と記憶システム]
- / Edit(管理者用)
2004年04月07日
■ JNS
"Climbing Neuronal Activity as an Event-Based Cortical Representation of Time."
Yakovlevらのイスラエルグループ。代表作はNature Neuroscienceの電気生理、Natureの行動実験だろう。
ITニューロンのdelayが上がってくるのがdelayの時間を変えるとslopeを変えてadjustするというもの。で、最も関連があると思われるわれわれの2001年の論文をreferしてない。われわれの論文ではclimbing activityは2secのうちにピークに達する。これが彼らのslopeの概念とのconsistencyが取れなかったが故だろうが、ああ、(省略)。
- / ツイートする
- / 投稿日: 2004年04月07日
- / カテゴリー: [内側側頭葉と記憶システム]
- / Edit(管理者用)
2003年08月04日
■ PNAS論文「霊長類側頭葉TE野から傍嗅皮質の視覚長期記憶を表象する神経細胞への前向性投射の解剖学的構成」
Anatomistかつphysiologistとしての仕事がPNASに載る。精細はPubMedまたはPNASへ。
A number of studies have shown that the perirhinal (PRh) cortex, which is part of the medial temporal lobe memory system, plays an important role in declarative long-term memory. The PRh cortex contains neurons that represent visual long-term memory. The aim of the present study is to characterize the anatomical organization of forward projections that mediate information flow from visual area TE to memory neurons in the PRh cortex. In monkeys performing a visual pair-association memory task, we conducted an extensive mapping of neuronal responses in the anteroventral part of area TE (TEav) and area 36 (A36) of the PRh cortex. Then, three retrograde tracers were separately injected into A36 and the distribution of retrograde labels in TEav was analyzed. We focused on the degree of divergent projections from TEav to memory neurons in A36, because the highly divergent nature of these forward fiber projections has been implicated in memory function. We found that the degree of divergent projection to memory neurons in A36 was smaller from the TEav neurons selective to learned pictures than from the nonselective TEav neurons. This result demonstrates that the anatomical difference (the divergence) correlates with the physiological difference (selectivity of TEav neurons to the learned pictures). Because the physiological difference is attributed to whether the projections are involved in information transmission required for memory neurons in A36, it can be speculated that the reduced divergent projection resulted from acquisition of visual long-term memory, possibly through retraction of the projecting axon collaterals.
吉田による訳:傍嗅皮質(内側側頭葉記憶システムの一部に当たる)が宣言的な長期記憶に関わっていることは多くの研究が示している。サルでの実験から、傍嗅皮質に視覚長期記憶を表象するニューロン(記憶ニューロン)があることがわかっている。本研究の目的は視覚連合皮質であるTE野から傍嗅皮質の記憶ニューロンへの情報伝達を担う前向性投射の解剖学的構成の特徴を見出すことにある。このため、視覚対連合記憶課題を遂行するサルのTEav野(TE野の前内側部)および36野(傍嗅皮質の外側部)の神経活動の精細なマップを作成した。つづいて、三種類の逆行性標識色素を36野に注入して、逆行性に標識された神経細胞の分布を解析した。本研究では、TEav野から36野の記憶ニューロンへの投射の神経側枝の広がりの具合(以降放散度と呼ぶ)について注目した。以前からTEav野から36野への投射の放散度の高さが記憶の機能と関連があると考えられていたからである。36野の記憶ニューロンへの投射の放散度は、TEav野のニューロンのうち学習した図形に選択的応答をするニューロンから投射するものでは、学習した図形に選択的応答をするニューロンから投射するものと比べて低かった。このことは解剖学的な違い(放散度)と生理学的な違い(TEav野のニューロンの学習した図形に対する応答)とが相関していることを表している。この生理学的違いとは、あるTEavニューロンが36野の記憶ニューロンの応答に関する情報処理に関わっているか、そうでないかの違いであると言える。よって、本研究で明らかになった放散度の減少は長期記憶の獲得から、おそらくは神経側枝の退縮によって引き起こされたものと推測する。
これが前のラボのメインプロジェクトでした。7年かかりました。
- / ツイートする
- / 投稿日: 2003年08月04日
- / カテゴリー: [内側側頭葉と記憶システム]
- / Edit(管理者用)
2001年04月08日
■ Science論文「霊長類側頭葉における記憶想起信号の逆行性伝播」
Scienceに論文が載る。Bossがcorresponding author、はじめの二人がequal contributionのfirst authorという形。精細はPubMedまたはScience magazineへ。
Science 2001 Jan 26;291(5504):661-664私が別のところで書いた考察:
Backward Spreading of Memory-Retrieval Signal in the Primate Temporal Cortex.
Naya Y,1* Yoshida M,2* Miyashita Y1,2,3†
1Laboratory of Cognitive Neuroscience, National Institute for Physiological Sciences, Okazaki, Aichi 444-8585, Japan., 2Mind Articulation Project, International Cooperative Research Project (ICORP), Japan Science and Technology Corporation, Yushima, Tokyo 113-0034, Japan., 3University of Tokyo School of Medicine, Hongo, Tokyo 113-0033, Japan.
* These authors contributed equally to this report.
† To whom correspondence should be addressed.
Bidirectional signaling between neocortex and limbic cortex has been hypothesized to contribute to the retrieval of long-term memory. We tested this hypothesis by comparing the time courses of perceptual and memory-retrieval signals in two neighboring areas in temporal cortex, area TE (TE) and perirhinal cortex (PRh), while monkeys were performing a visual pair-association task. Perceptual signal reached TE before PRh, confirming its forward propagation. In contrast, memory-retrieval signal appeared earlier in PRh, and TE neurons were then gradually recruited to represent the sought target. A reasonable interpretation of this finding is that the rich backward fiber projections from PRh to TE may underlie the activation of TE neurons that represent a visual object retrieved from long-term memory. 私による少し砕いた訳:長期記憶を想起するときには、大脳新皮質と大脳辺縁系との間での双方向の信号伝達が寄与している、と考えられてきた。我々はこの仮説を検証するために、視覚性対連合課題を解くサルの神経活動を記録して、側頭葉の隣接する脳部位であるTE野(新皮質)と傍嗅皮質(辺縁系)とでの知覚信号及び記憶想起信号の時間経過を比べた。図形を見たときに伝わる知覚信号は、まずTE野、それから傍嗅皮質へと順方向性に伝わっていた。一方、図形を思い出すときに伝わる記憶想起信号は、まず傍嗅皮質に先に現れ、それからTE野にだんだんと現れていった。以上の発見は、TE野のニューロンが長期記憶から想起した物体を表象するように活性化する際に、傍嗅皮質からTE野へと逆行性に投射する神経線維が関わっている、と解釈することができる。
対想起指数(pair-recall index)は傍嗅皮質の方がTE野と比べてより早く対連合図形がコードされ始めていた。このことは、視覚情報の流れとは逆に、長期記憶から取り出された想起された図形の情報は傍嗅皮質からTE野へと伝播していることを示唆している。また、順方向の視覚情報の伝達にかかる時間が10ms程度であるのに対して、逆方向の記憶情報の伝播には350msの時間がかかっていることは注目に値する。順方向の視覚情報の伝達はフィードフォワード的な伝達の連鎖と考えられるのに対して、逆方向の記憶情報の伝播はその様式が別のものであると考えられる。おそらくここで起こっていることは、単なるフィードフォワード的な伝達の連鎖ではなくて、傍嗅皮質とTE野を含むニューラルネットワークの中で決定付けられる状態遷移のようなものであるのであろう。このような考え方は、ニューラルネットワークのモデルの研究でも、非線形的なシステムに見られるアトラクター的ダイナミクスとして扱われている。 ところで、このような逆方向性の情報の伝達の生理的意義とは何であるだろうか。Logothetisら(Sheinberg and Logothetis 1997)はマカクサルに両眼視野闘争の課題を行わせた実験によって、TE野の活動が[視覚入力そのもの]ではなく、[何が見えたか(visual awareness)]に関わっているものであることを示唆している。もし我々の実験で見られたTE野の想起過程の活動も同様にして、[これから現れる図形を思い描くこと(visual imagery)]に対応しているのであるとするならば、一方で傍嗅皮質の活動は無意識の/自動的な連想過程を表現している、というようにTE野とは役割が分かれているのかもしれない。これは記憶、表象が明示的/暗示的(意識に上るか否か)であるとはどういうことかを明らかにするのに重要な鍵となるかもしれない。 考察すべきもう一点は、今回の発見でわかったことと前頭葉の活動との関連である。記憶/想起のシステムを考えると、さらに第3のプレーヤーとして前頭葉を置いて考えることが有意義だ。以前我々グループは前頭葉から下部側頭葉へとvoluntary recallに関わるトップダウンシグナルが流れていることを報告している(Hasegawa et.al.1998及びTomita et.al. 1999)。すると、傍嗅皮質からTE野への想起信号の自動的な伝播過程は前頭葉からの自発的な想起のシグナルによってトリガーされている、という図式が考えられる。新聞記者を前に記者会見をするという経験を生まれてはじめてした(教授の横に座っていただけだけど)。5年かかってやっとプロジェクトの成果が一つ出たといったところ。この論文の内容についてもっとわかりやすく説明した文章を書こうと思うのだが、いつのことになるやら。なお、生理研のホームページにこの論文のまとめ(日本語)があり。
- / ツイートする
- / 投稿日: 2001年04月08日
- / カテゴリー: [内側側頭葉と記憶システム]
- / Edit(管理者用)
お勧めエントリ
- 細胞外電極はなにを見ているか(1) 20080727 (2) リニューアル版 20081107
- 総説 長期記憶の脳内メカニズム 20100909
- 駒場講義2013 「意識の科学的研究 - 盲視を起点に」20130626
- 駒場講義2012レジメ 意識と注意の脳内メカニズム(1) 注意 20121010 (2) 意識 20121011
- 視覚、注意、言語で3*2の背側、腹側経路説 20140119
- 脳科学辞典の項目書いた 「盲視」 20130407
- 脳科学辞典の項目書いた 「気づき」 20130228
- 脳科学辞典の項目書いた 「サリエンシー」 20121224
- 脳科学辞典の項目書いた 「マイクロサッケード」 20121227
- 盲視でおこる「なにかあるかんじ」 20110126
- DKL色空間についてまとめ 20090113
- 科学基礎論学会 秋の研究例会 ワークショップ「意識の神経科学と神経現象学」レジメ 20131102
- ギャラガー&ザハヴィ『現象学的な心』合評会レジメ 20130628
- Marrのrepresentationとprocessをベイトソン流に解釈する (1) 20100317 (2) 20100317
- 半側空間無視と同名半盲とは区別できるか?(1) 20080220 (2) 半側空間無視の原因部位は? 20080221
- MarrのVisionの最初と最後だけを読む 20071213