[カテゴリー別保管庫] 細胞外電位
2022年03月27日
■ 「細胞外電極はなにを見ているか」をちょっとアップデート
昔作った「細胞外電極はなにを見ているか」をぜんぶ繋げ直してアップデートしておきたい。
ラボセミナー用にちょっとアップデートしたので、そのときのスライドを貼っておく。まだ直すべきところはたくさんある。先は遠い。
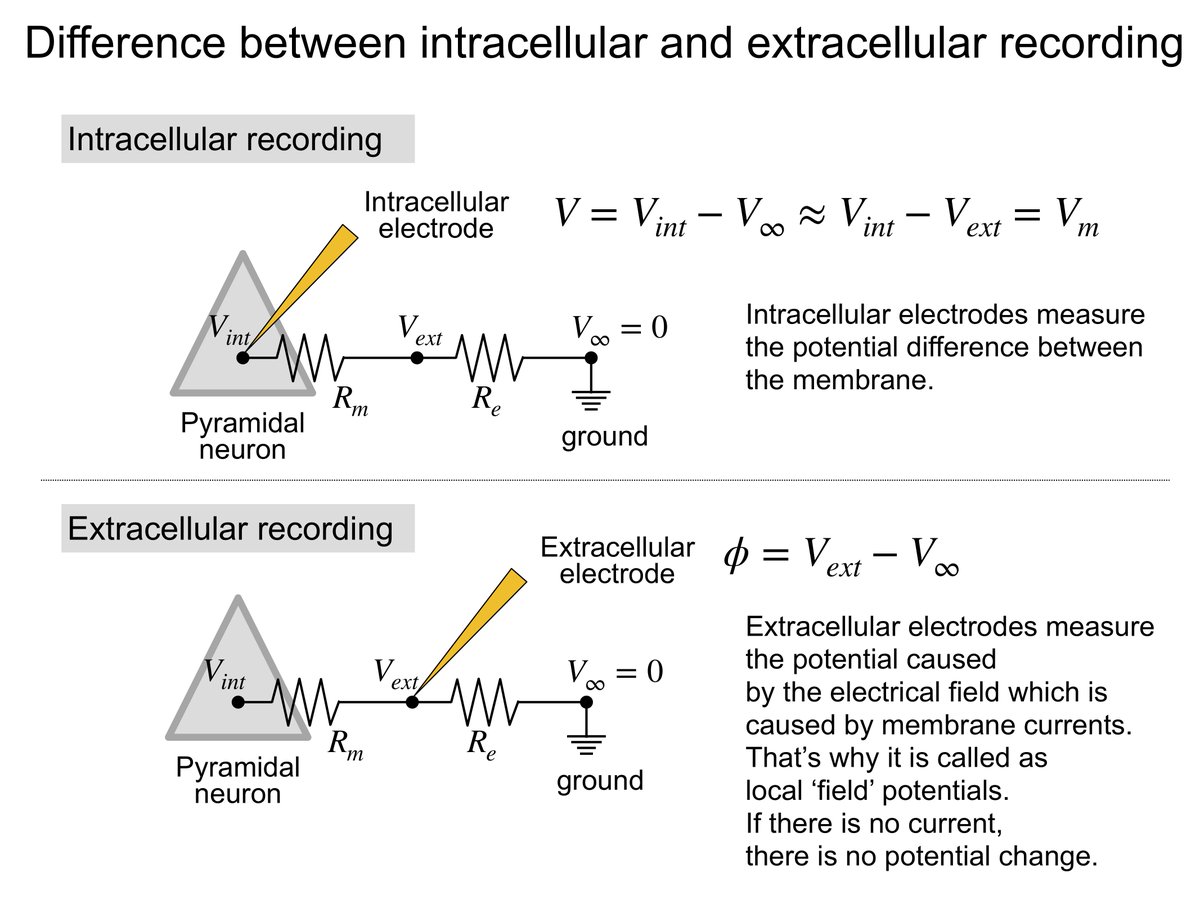
図1
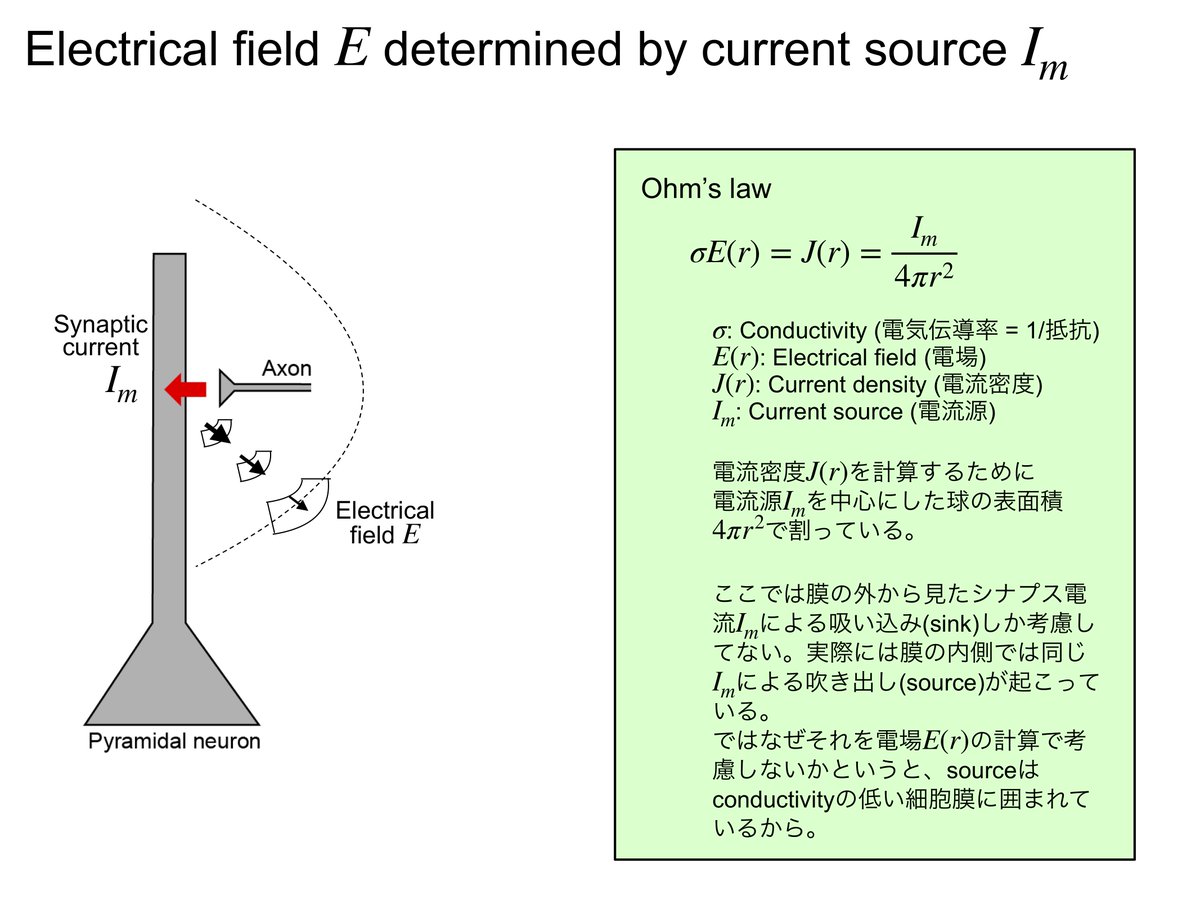
図2
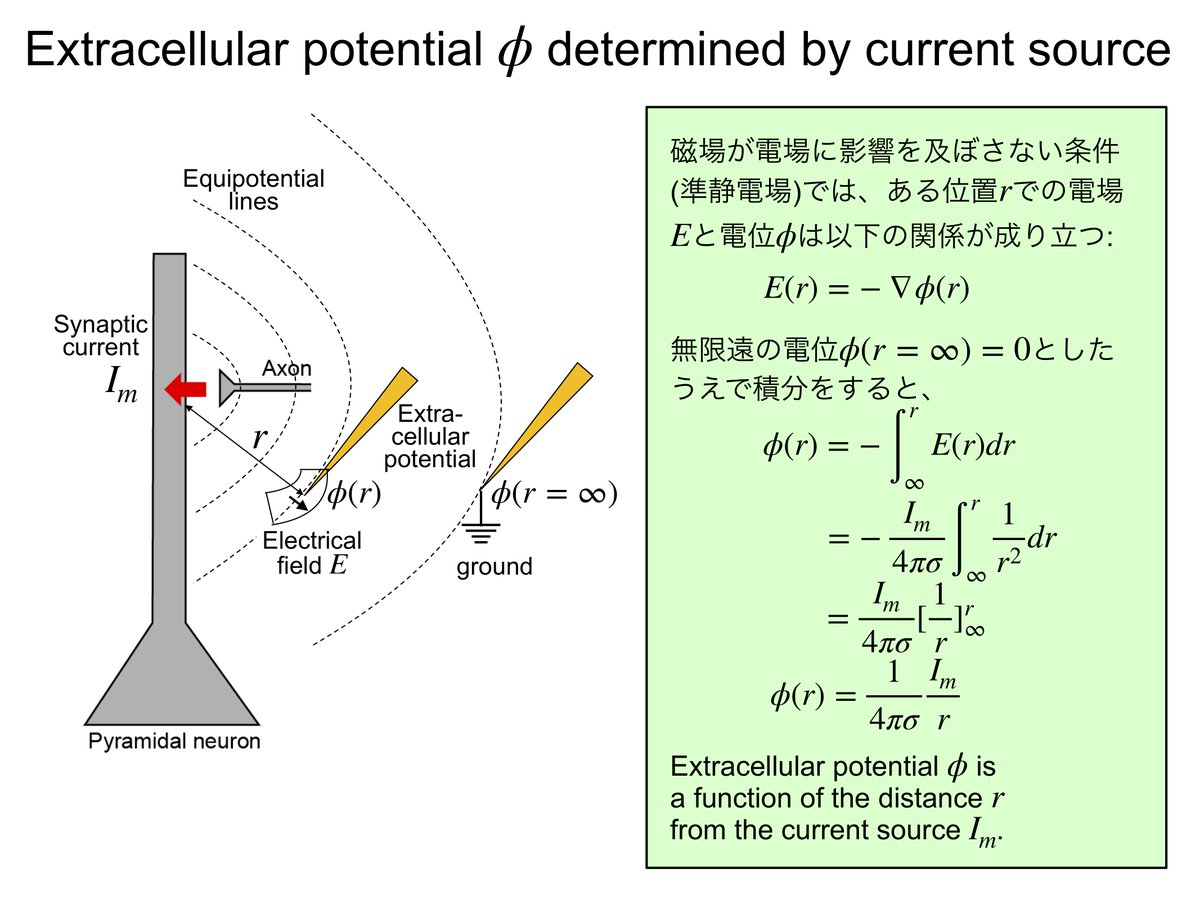
図3
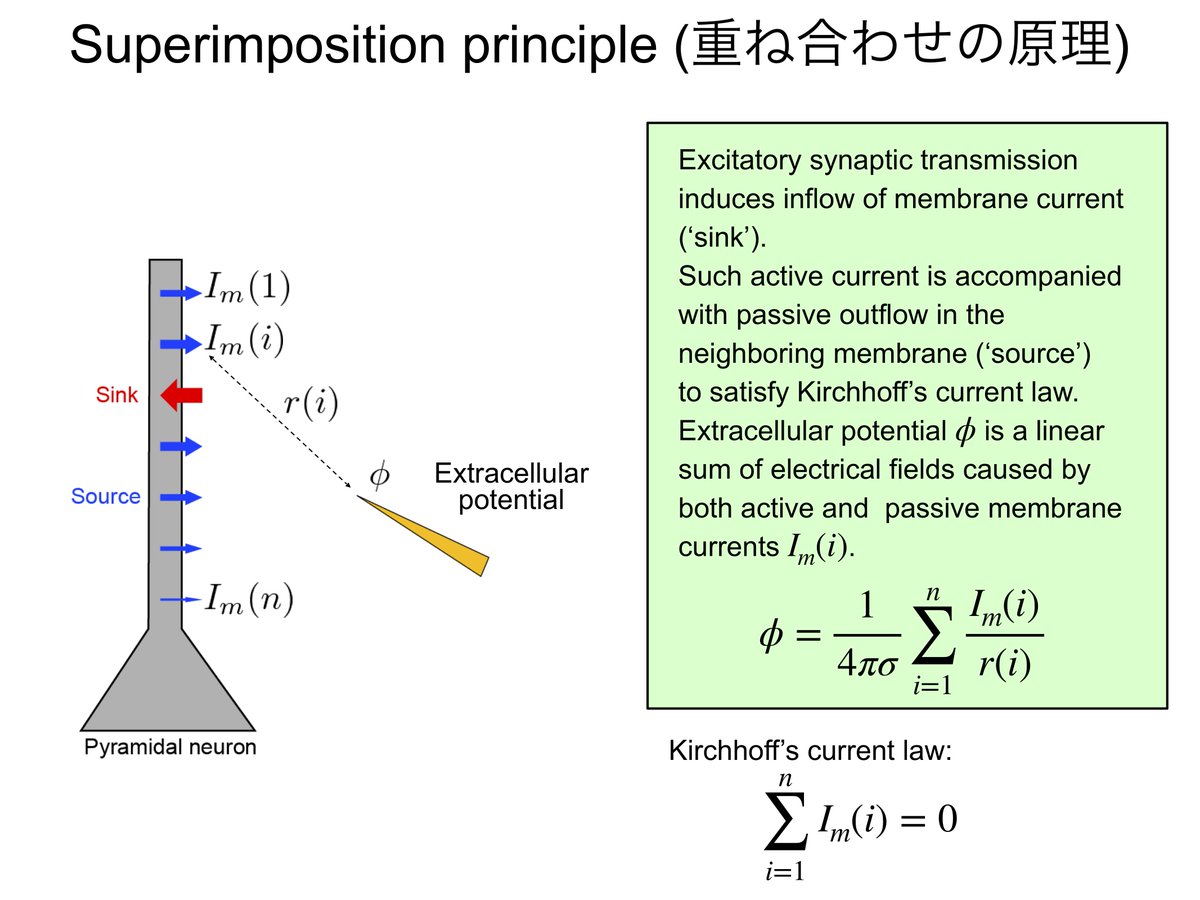
図4
2008年12月31日
■ 応用問題:海馬sliceのfield EPSP
海馬スライスを使ったLTPの実験では、CA3からCA1への入力繊維であるSchaffer collateralを電気刺激して、CA1でのシナプス応答を記録するという実験が盛んに行われた。Whole-cell recordingだと1時間以上の記録は難しいから、おそらく現在でも長時間の記録をしたいときとかには使われるんではないかと思う。このときなにを記録しているかを考えてみよう。
CA1のstratum radiatum(apical dendriteのあるlayer)に細胞外電極を置いてSchaffer collateralを電気刺激すると、下向きのfield potentialが記録できる。これはSchaffer collateralからCA1 pyramidal neuronのapical dendriteに入る興奮性シナプス電流を反映している(field EPSPと呼ばれる)。一方で、CA1のpyramidal layerに細胞外電極を置いてSchaffer collateralを電気刺激すると、上向きのfield potentialに混じって鋭いnegativeな波形が見られる。前者はdendriteへ入力する興奮性シナプス電流が離れた場所でsourceとして見えているということ。鋭いnegativeな波形は電極直下のpyramidal neuronのaction potentialがsourceとして記録されていることを意味している(population spikeと呼ばれる)。Preのfiberを電気刺激しているため、多くのpyramidal neuronが同期して発火するので、細胞外電位として検出できるわけだ。
以前書いたことのある疑問(20050818)だけど、ではなぜstratum radiatumから記録しているときにはpopulation spikeの成分がsourceとして見えてこないのだろうか。これに対する答えは今回のエントリの範囲は超えるのだけれど、細胞外電位の広がりには周波数特性があることが知られている。つまり、ローパスになっている。だから、離れたところで起こったaction potentialはdendritic layerでは記録できないから、というのがとりあえずの答えになる。じっさい、single-unitの記録の経験からもローパス特性についてはよくわかる。電極がニューロンから離れているときはポジの成分ははっきりとしているけれども、ネガの成分が充分シャープでないため、単一のニューロンとして記録する(「isolateする」と言う)ことができない。電極を進めていってニューロンの近傍まで近づくと、ネガの成分が鋭く大きくなってきて、isolateできるようになる。ポジもネガも同じニューロンのaction potentialが元なので、このような違いはaction potential(による細胞膜を流れる電流)による電位変化がvolume conductorを伝播しているところで、高周波成分が落ちたということを示している。
Computationalにこの問題を扱っている人がいて、前述のBedard C et.al (2004)ではこれを電流発生部分から電極までの広がり(volume conductor)でのconductivity(=抵抗の逆数)が場所によって不均一だから、としている。つまり、simulationをしてやって、conductivityが均一なモデルだとそのようなローパス特性が起こらないが、不均一だと起こるということを示している。
ただ、よくわかんないのは、Buszakiの本を読んでいると、 (書きかけだけどアップ。)
2008年11月07日
■ 細胞外電極はなにを見ているか リニューアル版
京都で開催された脳プロ分科会の「皮質脳波」のセクションで「細胞外電極はなにを見ているか」ということで10分トークをしてきました。
以前のブログのエントリ(「細胞外電極はなにを見ているか」)を元ネタにして作成していたのですが、あれは電気生理はじめての方も含めたトレーニングコースでのチュートリアルでしたが、今回は神経生理、工学系、臨床系の先生方の前ですので、かなり変更を加えることになって、けっきょくのところほとんど新しいものを作ることになりました。
そうしたら準備した内容が10分ではとてもしゃべりきれないものになりましたので、当日はいちばん関連する部分だけしゃべることにして、増補したものをハンドアウトで配布しました。さらに増補したものをLaTeXを使ってPDFファイルとして文書を作成しましたので公開します。ダウンロードはこちらから:「細胞外電極はなにを見ているか」(pdf)
最重要ポイントは、ニューロンの活動から細胞外電位が発生する過程の「順問題」と細胞外電位からニューロンの活動を推測する「逆問題」とに分けて考えることができるということです。図にするとこんなかんじです:
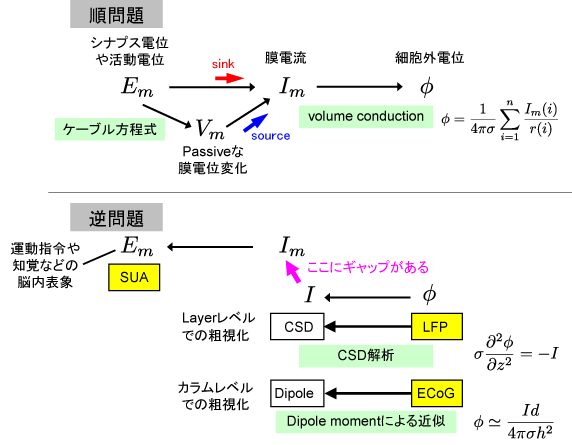
くわしいことはPDFの文書をご覧下さい。
今回のプレゼンとこの文書にはいくつか狙いがあって、1) BMIという文脈で期待されていることにきっちり答える。2) Decodingとデータベース作成に関する自分の立ち位置を明確にする。3) トレーニングコースとブログで再利用。4) チュートリアル記事の執筆依頼お待ちしております。というかんじだったのですが、当日はECoG電極のspacingの議論に質問が集中しました。わたしはECoG使ったことないんですけど、専門家面して意見言ってよかったんだろうか。
ともあれプレゼンの方はけっこうおもしろがっていただけたのではないでしょうか。あとで何人かの方に声をかけていただきましたし。私も第一旭と新福菜館と駅ビルの「すみれ」でラーメン三食食って満足。(<-食い過ぎ)
LaTeXはインストールが面倒なのでなるたけmimeTexとかでなんとかしてきましたが、今回はさすがにそういうわけにもいかないなと思ってひさびさに調べてみたら、TeXインストーラ3というのでW32TeXを含むすべてが一括インストールできるようになっていて感激しました。ほんとはアンインストールも一発でさせてほしいけど。
今回powerpointに数式を貼りつけるときにTeXclipにお世話になりましたが、これは素晴らしいですね。(いま気づいたけど作者の方はMouseoverDictionaryの作者でもあるんですね。こちらもお世話になってます。)
こんなかんじでlatexの原稿ファイルをアップロードするとPDFが作成できる、みたいなwebサービスが出てこないですかね。本一冊書くとかでなければ、そういうので全然じゅうぶんだと思うんですけど。精神としてはhtmlみたいに、コンテンツ作る人はコンテンツ作りに集中すべきで、タグ打ちとかもなるたけ省力化する方に行くのがあるべき方向だと思うんですけど。数式書くときしか必要ないからなあ。論文書くのだってバージョン管理システムを使うよりはワードの履歴機能使う方が圧倒的によいですからね。
って話がずれてきたので、終了。
# mmrl
ども、おひさしぶり、mmrlです。
現在新しい多点電極で深いところの細胞外記録チャレンジ中なのですが、スパイク波形が従来のやり方と異なってとれてきていて、スパイクソーティングに悩んでいました。
むか〜し勉強した電磁気学の教科書と、Kochのを引っ張りだしてきてうんうんうなっていたのですが、大変参考になる資料を作っていただきまして非常に勉強になります。
ご無沙汰しております。
>>現在新しい多点電極で深いところの細胞外記録チャレンジ中
おー、たぶん深いへんだとニューロンのdendriteの形状が違うからpyramidal neuron用のアルゴリズムだと最適ではないのかもしれませんね。
"Biophysics of Computation"とか"Methods in Neuronal Modeling: From Ions to Networks"とかも読んでおかないといけないのですが、けっきょくHolt-Koch論文とかですませました。
すみません。気になったので投稿します。
以前のエントリですが、
Vext = R/(4*pi)*Σ(Ij / rj) ---(3)
ただし、Vextは細胞外電極で測定された電位、Rは細胞間隙の抵抗でだいたい200-300ohm/cm。Ijはある場所jで流れた電流の大きさ、rjは場所jから細胞外電極までの距離。Σはすべての電流源jでの総和。(Nunez and Srinivasan(2006)より)
なんとなく意味は通じるのですが、空間上の一点を間接的に表現している r と一点では表現し得ない I が混在している時点で(3)の表式に違和感あるんですが。
生物系では慣例的にこういう表式するんですか?
psycho-resiさん、こんにちは。はてなダイアリー拝見しました。
物理出身で精神科医をされていて、ECTの効果を脳内でのマクスウェル方程式を解いてシミュレーションしてるということは、私よかよっぽどこのへんについてくわしいと思うんでこっちの方が教えていただきたいです。
ご質問いただいた件ですが、何段階かポイントがありまして、わたしもしぼりきれないのでそれぞれ書いてみます。
1) まず、ご質問への直の答えですが、上記の式自体は言及している教科書Nunez and Srinivasan(2006)の中でじっさいに使われている式ですし(p.168)、他の論文などでもニューロンの活動から周りの電位をシミュレーションするときによく使われる式です。
2) 新しい記事の方もそうですが、わたしの言葉遣いとして、currentとcurrent sourceや、currentとcurrent densityを厳密に使い分けていないところがあります。たとえば上記のIjはcurrent sourceと言うべきでした。このへんを正しくしたら問題は解決するでしょうか?
3) もし上記の式が∑ではなくて積分形で、微小領域dSで流出入するcurrent source I(x,y,z)の足し合わせ(∫I/R dS)だったら違和感は解消するでしょうか?
4) 電磁気学の教科書によく出てくるのは上記のcurrent sourceがpotentialを作るものではなくて、chargeがpotentialを作るものですが、こちらの場合も上記の教科書では φ = 1/(4*pi*ε)∑(qj/Rj) という表記がされています(φは電位、qjは位置jでのcharge sourceの量で、εは誘電率)。これには違和感はありますか? 積分形なら問題ないですか?
5) 細胞外電位で見ている現象は、細胞膜が静止膜電位で分極しているchargeによって影響を受けるようなミクロな系ではないので、4)のchargeの式ではなくて上記のcurrentの式を使うこと、上記のcurrentの式はあくまで細胞外電位で見ているようなmacroscopicな物理現象でのみ妥当であること、が上記の教科書には書かれています。このあたりが論点でしょうか?
以上です。ほかにもぜひこれらのエントリや文書で直すべき点、改良すべき点などありましたらご指摘いただけるとありがたいです。
いえ、あんまりわかってないです。
ECTの科学的な基礎ができてないと感じたので着手したんですが、できたのは静解析のみです。マクロの大雑把な電位勾配や電流密度はそれらしく計算できたんですが、ミクロのつまりニューロンとそれらがどう相互作用するのかは課題として残ってます。
質問に関してですが、私、その教科書持ってないので印象でしかいえないんですが、膜上の局在した領域でのみ電荷の移動がおこるという前提があるようですね。ならばそのn番目の領域dSでの電流密度ベクトルjnを定義して
In=∫jndS
細胞外電極で測定された電位Vextはこれらの足し合わせだから
Vext = R/(4*pi)*Σ(In / rn)
とすると私の場合は激しく納得します(少なくとも表記の上では。式自体はどうやって誘導されたかわかりません)。
前提の理解が違ってたらたぶん見当違いのこといってます。
(4)は違和感なし。
(5)は難しいんでまた考えさせてください。
たぶん、問題の式がわかりにくい理由の一つは、Imがcurrentやcurrent densityではなく、current sourceをあらわしていることによると思われます。current sourceは-div(Jsource)(Jsourceは膜を通過するcurrent densityのうち、passiveな抵抗性電流で説明されないもの)と定義される量で、マクロ系の電磁気学でみかけるものです。
currentやcurrent densityを考えると、電荷の偏りがないかぎり、電流は閉じているはずなので(d(rho)/dt=0 -> divJ=0)、電流がある点にしかないというのはありえないことです。しかしながら、-div(Jsource)は、点状に分布し得ます。
ここのところは、マクロ系の電磁気学の議論に類似しています。すなわち、current density(J)をオームの法則の法則に従うpassiveな抵抗性電流(sigma*E)と、それ以外(=容量性電流 + active conductanceなど)に分解します。この後者をJsourceと呼ぶことにします。J = sigma*E + Jsource。Jについては、電流は閉じているはずなので、divJ=0。
active conductanceや容量性電流などがなければ、(sigma=const.として)常に、div(E)=0で、磁場でもあたえない限り、rot(E)も0で電場もなにも発生しないのですが、active conductanceが働くと、div(E)は0とは限らず、電場および電位が発生することになります。
ここでdiv(J)=0、およびJ = sigma*E + Jsourceより、
div(E)= - div(Jsource) / sigma = (current source) / sigma。
したがって電場(phi = - grad(E))は、
phi = -Laplacian (current source) / sigma
となって、rot(E)=0より、r->infinity でphi=0の境界条件のもとで
(3)式のように解くことができます。
psycho-resiさん、どうもありがとうございます。基本的にはわたしのコメントの2)での「たとえば上記のIjはcurrent sourceと言うべきでした。」というのが論点だったようですね。
あと、教科書(Nunez and Srinivasan(2006))での記載では、たしかにI_mは点ではなくて、ある一定の大きさを持った小領域になっています。つまり、球をどんどん小さくしていって、その外側ではdiv(J)=0が成り立ち、その球面より内側ではdiv(J)=s(source current)になっていて、この球の中心からr離れた場所での電位φを計算する式となっています。この関係を使って、膜に広がって分布しているcurrent source/sinkを小領域に分割して足し合わせる、というのがここでやっていることであると理解しました。
式(3)の表現の仕方としては、ケーブル方程式で求まるtransmembrane current I_mという細胞の中から見た値が、細胞外からはsource current s (≡-div(J_source) )として捉えられていて、中からと外からの表現が混ざっているというあたりの問題なのかなとも思いました。
OK さん、どうもありがとうございます。ご指摘の「細胞膜ではdiv(J)=-I_m、それ以外ではdiv(J)=0」については、今回のPDFファイルの注5で多少言及しておりますが、以前のエントリのときはまだこのへんがわかっていませんでした。
J = σ*E + Jsourceの式に関してはOKさんの解説で非常にすっきりわかりました。どうもありがとうございます。(じつはこの式はNunez and Srinivasan(2006)のp.166で扱われていまして、該当部分を読んではいたんですが、十分理解できていませんでした。) Active conductanceとcapacitative currentだけがdiv(J) ≠ 0の点を作る、というのはとてもクリアーで、なんで膜にだけcurrent source/sinkがあるのかが明確になりました。(なんとなく、電極からは膜の向こうは抵抗が大きすぎて「見えない」から、その境界から出入りする電流はみな無から有が生まれたように見えるので、current source/sinkになる、というような理解の仕方をしていました。)
そうするとひとつ新たな疑問が生まれてきたのですが、わたしの図式では、ケーブル方程式で transmembrane potential V_mを計算した後に、transmembrane current I_mを抵抗性成分g_m*V_mと容量性成分cm*dV_m/dtの和として計算していました。その上でI_m/rを足し合わせる(式3)ということをしていました。しかし、電位に影響を及ぼすのが容量性成分だけであるなら、source currentの値としては、抵抗性成分g_m*V_mを差っ引くべきではないのでしょうか? つまり、ある位置での膜のsource current s = I_m - g_m*V_m であって、&phi = ∑ (I_m - g_m*V_m)/r とすべきではないのでしょうか? ただ、今回参照したPettersen KH, Einevoll GT. Biophys J. 2008 Feb 1;94(3):784-802では、式(3)のI_mがlinear leak currentとcapacitative currentの両方を含んでいると書いていますので、たぶんわたしがいま言ったことは間違っているんだろうと思いますが、このへんがまだよく理解できていません。
"Jsourceは膜を通過するcurrent densityのうち、passiveな抵抗性電流で説明されないもの"と書いたのは間違いでした。膜を通過する電流は抵抗性成分、容量性成分どちらも含める式で正しいと思います。
div(E)= - div(Jsource) / sigma とするときに、sigmaは一定でなければいけないので、細胞外空間だけを考えて、細胞膜のところは境界条件として扱うので、膜を通過する電流の抵抗性成分はsigma*Eには含まれず、Imのほうに入ってくるのだと思います。
OKさん、長々とおつきあいくださいましてどうもありがとうございます。これは細胞外の(σ1)と細胞膜の(σ2, ε)というふたつの領域があって、元々の話は細胞外の(σ1)の環境でのポアソン方程式を解くのに細胞膜の表面を境界条件として使うという話であって、私がケーブル方程式云々で言っていたのは細胞膜の(σ2, ε)の環境でのポアソン方程式を解くときの話であって、両者をごっちゃにしてしまったということのようです。またもう少し時間を取って勉強してみたいと思います。とりいそぎ御礼まで。
# 元物理系学生このスレ長いですね。
この話題に微妙に関係するご相談なのですが、先日投稿した論文にレビュアーからコメントが返ってきました。
かなり見当違いなコメントで正直相手にしたくないのですが、なかでも凄いのが
between the field strength in the brain tissue with ECT and possible neurophysiological responses
の関係を明らかにせよというもの。
それがなかなかできないからシミュレーションしているんでしょうに…。
それでなんですが、仮に実験系を組むとしたらどんな感じがいいかご相談よろしいですか。
考えようによっては、この分野、基礎から臨床までシームレスですね。
>>このスレ長いですね。
以前はコメントが15個並んだこともあったのですが
(two-photon in_vivo imagingのカテゴリ)、これだけ長いのも久しぶりです。
>>between the field strength in the brain tissue with ECT and possible neurophysiological responsesの関係を明らかにせよというもの。
ちょっとこれだけだとよくわかりませんが、rodentとかで電気刺激によるseizureの誘導と記録の実験とかそういった過去の論文を引いて議論すればよいのではないですか?
実験系、というとちょっとわからないですね。
今回作ったPDF文書の中ではLogothetisのNeuron 2007論文というのを言及していて、それとかは微小電気刺激をしたときの応答からconductanceを計算してたりします。ただ、たぶんECTの場合はseizureまで起こすのが必須なんだと思うんで、ちょっと違いそうではありますが。
ではまた。
どもです。現在LFPとスパイクの関係を調べてる者で、共同研究者が吉田さんのレジメを発見して来てくれたのは良かったのですが、僕も式(1)のphi = (1/4pi sigma) I_m/rで混乱してしまい、このスレで納得しました。感謝感謝。それで細かい事で申し訳ないですが、OKさんの
>>したがって電場(phi = - grad(E))は、
>>phi = -Laplacian (current source) / sigma
>>となって、rot(E)=0より、r->infinity でphi=0の境界条件のもとで
>>(3)式のように解くことができます。
はE = -grad phiと
current source = -(1/sigma) Laplacian phi
の間違いですよね。
Kensukeさん、どうもありがとうございます。ちょっともう内容を忘れかけてますが、「E = -grad phi」の式はたしかにひっくり返ってましたね。
PDFのファイルのほうも、ここで皆様が書いてくださったことを元にしてアップデートしておきたいのですが、頭がそっちに戻らないので止まってます。
Kensukeさん、物理屋さんですよね。もしよければ、PDFの式(1)のあたりの表現をどう変えるといいかsuggestしていただければ、acknowledgement付けてPDFをアップデートしたいと思いますので、ぜひご考慮いただければ。
もしくはappendixに新たに項目を設けるとか。pLatexで書いてくだされば対応できますのでぜひ。
2008年08月05日
■ トレーニングコース無事終了
トレーニングコースのほうは無事終了しました。それでレクチャーノートを作る際にいくつかアップデートしたのでこのあいだのエントリ「細胞外電極はなにを見ているか」をアップデートさせておきました。
元のはけっこう間違いがあったのだけれども、ずいぶんマシになりました。なによりも、わたしCSD完全に勘違いしてましたよ。それがそのまま通ってるっていう恐ろしさ。
いろいろ付け加えた分、簡潔さは失われたかもしれなせん。このへんのさじ加減がなかなか難しい。
この一時期にだーっと考えてまた来年のトレーニングコース前になるとまたあわてて資料をひっくり返すとかそういうかんじになると思われ。(<-さいきん「よくわかる現代魔法」読んだもんで。)
さて、こっちはもはや忘却の彼方へ押しやって、これからは生理研研究会の予習シリーズへと行くことにしましょう。
2008年07月27日
■ 細胞外電極はなにを見ているか
夏が来れば思い出す・・・のは生理研トレーニングコースの準備です。ここ数年は当研究室では、「麻酔下動物での急性電気生理実験」というタイトルで、細胞外電極を使って麻酔下ratからLFPを記録して、視覚刺激に対する上丘の応答をみたり、ひげ刺激に対するbarrel cortexの応答を見るということをやっております。でもって、LFPで記録しているものがなにか、ということを毎年わたしがちょこっと講義しています。
問題なのは、適切な教科書がないということです。この問題については以前にも書きました(20050817および20050818)。細胞内電極やパッチクランプ用電極を使った細胞内電位の記録に関しては、ホジキン・ハックスレーあたりについて書いている生理学の教科書を読むことから始めて、"the axon guide"あたりまで読んでおけばだいたい入門できるかと思うのですが、細胞外電極を使った細胞外電位の記録に関しては良いものがありません。わたしが日々行っている、行動中のnhpのsingle-unit recordingがまさに、タングステン製の細胞外電極を使って、単一細胞のaction potentialを細胞外電位から記録しているものなのですから、よろしくない事態です。また、multiunit recordingやLFPなどが今後BMIなどの応用の面からさらに重要になってくるであろうことを考えると、このへんの基礎をきっちり押さえておくことは大切なことだと思います。
それで、今年もトレーニングコースの時期がやってきましたので、いそいそと準備しているのですが、しょうがないので自分でいろいろ調べて作ってみました。まだ自分で納得いけてないところがあるのでここで皆様の意見を聞きたい、というわけです。物理をわかっている方ぜひご協力お願いします。それではここから:
追記081108:リニューアル版のエントリを作りました。そちらもご覧ください。
(印刷用にPDFファイルを作りました。ファイルをダウンロード。数式のフォントがきれいでないのでhtmlを印刷した方がよいかも。)
(追記:いろいろ書き直してます。オリジナルのエントリはpdfファイルとして保管しておきます。)
[まとめ]
- 細胞外電極はなにを計測している? - 細胞膜を流れる電流。
- 細胞膜の間を流れる電流の正体は? - シナプス電流とaction potential。
- 低周波成分はLFPと呼ばれ、シナプス電流を反映している。
- 高周波成分はMUAと呼ばれ、action potentialを反映している。
- LFPは必ずしも電極の近傍で起こっている電流を反映しているとは限らない。離れたところで起こっている電流を反映している可能性がある。
- CSD解析では、さまざまな深さから記録した細胞外電位から、細胞膜の間を流れる電流の発生位置を推定する。
- 細胞外電極が神経細胞の非常に近傍にあるとき(<50µm)には、単一の神経細胞からの活動電位を反映したsingle-unitを記録することができる。
[細胞外電極はなにを計測している?]
ひとことで言えば、電極近傍の細胞膜を流れる電流を計測している。細胞膜電位ではない点に注意。
もっと正確に言うために、細胞内電極での記録と比較してみよう。図1(1)にあるように、細胞内電極は細胞内の電位Vintと無限遠に置いた不関電極での電位V∞との差を計測している。これはけっきょくのところ、細胞膜間の電位差(細胞膜電位)を計測していることになる。
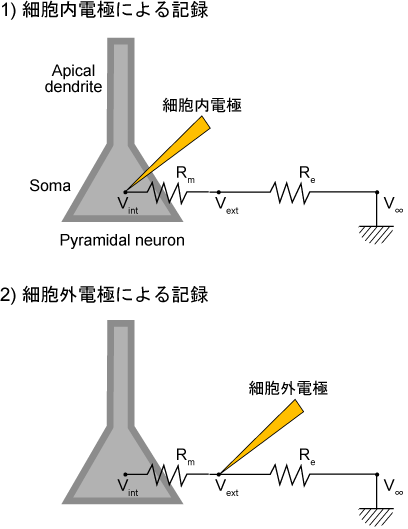
図1 細胞内電極(1)と細胞外電極(2)の記録の違い
Vint - V∞ ~= Vint - Vext (Rm >> Rextより、) = Vm ---(1)
細胞膜の両端にかかっている電位を計測しているという意味で、これはテスターである部分の電位差を測るのとまったく同じである。
一方で、細胞外電極(図1(2))は細胞間隙に置いた細胞外電極での電位Vextと無限遠に置いた不関電極V∞とのあいだの電位差(細胞外電位)を測定している。
Vext - V∞ ---(2)
しかし、ここで測定されたReの両端の電位差そのものには生理学的な意味がない。つまり、テスターでの計測をイメージするとおかしなことになる。
じつは、細胞外電極で記録される電位差は、細胞間隙に置いた細胞外電極の周りで起こった電流によって出来た電場を反映しているのだ。
そして、重ね合わせの原理から、細胞外電位は近傍で細胞膜の間を流れる電流量による電位変化の足し算である。電流の発生源から電極までの距離に反比例して弱くなる。数式で書くと、
Vext = R/(4*pi)*Σ(Ij / rj) ---(3)
ただし、Vextは細胞外電極で測定された電位、Rは細胞間隙の抵抗でだいたい200-300ohm/cm。Ijはある場所jで流れた電流の大きさ、rjは場所jから細胞外電極までの距離。Σはすべての電流源jでの総和。(Nunez and Srinivasan(2006)より)
だから、無限遠に置いた不関電極では、r=∞でVext = V∞ = 0となり、電流の影響が全くないところでの電位を計測していることになる。不関電極との差分を計測することで細胞外電極は電流の影響を計測していると言える。
上記の(3)式についてもうすこしくわしく書くと、クーロンの定理から
E(r) = R/(4*pi)*I/r2 ---(4)電流Iがあると、r離れた部分に電場E(r)ができる。後述の通り、電場E(r)はベクトル場。
電場ができるとそこには電位が生まれる。つまり、
E(x,y,z) = -grad(V(x,y,z)) ---(5)ある地点(x,y,z)の細胞外電位V(スカラー場)の勾配gradientを取るとその地点の電場E(ベクトル場)になる、というふうに電位が定義される。
以上の(4)(5)からEとqを消去して、sourceごとに足し併せると(3)式になる。
ここでつかうべき(4)の式はcharge sourceの式ではなくてcurrent sourceの式であるべきだからこれで良いと思うのだけれど、Nunez and Srinivasan(2006)には記載がない...
[細胞膜の間を流れる電流の正体は?]
では、「細胞膜の間を流れる電流」とはいったいなにか。主に二つのものが考えられる。
- シナプス電流(EPSCとIPSC)。
- Action potential(に伴ったNa,Ca流入、K流出)。
これらはべつべつの周波数帯域と空間解像度を持っている。よって同じ電極からフィルタを使って両方の活動を分離することができる。
Local field potential (LFP):
- 周波数成分 1Hz-300Hzあたり
- 主に電極近傍のsynaptic current (EPSC, IPSC)による。
- synaptic currentはamplitudeは小さいが、time courseが長いので、近くの入力のspatial summationが起こる。
- 電極先端から0.5-3mm程度の範囲のシナプス電流が寄与する。(Logothetis (2008)より孫引き。) 電極の抵抗に依存する。
- あくまでシナプスの位置が重要であり、電極先端に細胞体があることを保証しない。このへんについては「Volume conduction、sinkとsource」にて後述。
Multi unit activity (MUA):
- 周波数成分 300Hz-5KHzあたり
- 電極近傍のaction potential による。
- action potentialはamplitudeは大きいが、time courseが短いので、近くの入力と<1msオーダーで同期しない限り、キャンセルアウトする。
- 電極先端から50-350µm程度の範囲のニューロンの活動電位が寄与する。(Logothetis (2008)より孫引き。) 電極の抵抗に依存する。
つまり、ざっくり言えば、一つの細胞外電極から記録される信号を周波数で分けることによって、LFPは入力(=シナプス電流)、MUAは出力(=action potential)、と違ったものを分けて見ることができる。
(なお、さいきんでは、action potentialのあとで見られるafterhyperpolarizationなどの遅いnon-synapticな成分がLFPに効いていることがわかってきている。けれども、それを入門のときに言うと混乱するので言わない。)
[Volume conduction、sinkとsource]
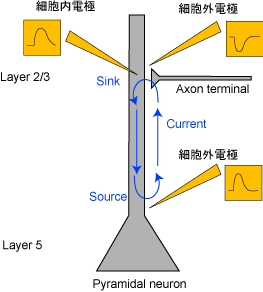
図2 興奮性シナプス電流
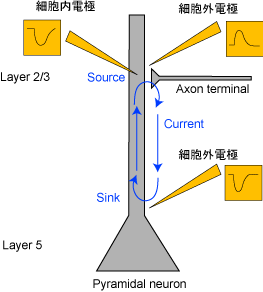
図3 抑制性シナプス電流
上の段で「細胞膜の間を流れる電流」には二つある、と書いたがそれは正確でない。上記のようなシナプス電流やaction potentialなどでactiveな電流が出来たときには、閉回路を作るためにpassiveな電流も流れる。よって、電極から記録されている現象が電極の近傍で起こっているactiveな電流によるものなのか、それとも離れたところで起こっている電流から作られたpassiveな電流によるものなのかは自明ではない。
たとえば、図2のようにシナプスで興奮性シナプス電流(EPSC)が起こっているとき、dendriteおよびsomaで細胞内電位の上昇が見られるが、細胞外電位はdendriteでは近傍の電流流入を反映した下向きのpotential(sink)が記録されるが、somaではpassiveな電流流出を反映した上向きのpotential(source)が記録される。よって、電極からsourceが記録されたとしても、かならずしも近傍の抑制性シナプス電流(IPSC)を反映しているわけではない。これは図3のように抑制性シナプス電流(IPSC)が起こっているときの図と併せて考えてみればわかる。つまり、下向きの細胞外電位が記録できたとしても、それは近傍での興奮性シナプス電流(EPSC)によるものかもしれないし、離れたところでの抑制性シナプス電流(IPSC)によるものかもしれない。
ともあれ、action potentialやシナプス電流によるイオンの流入流出よるactiveな電流にはpassiveな電流が伴う。これによってsinkとsourceのペアが生まれる。
このsink(吸い込み)とsource(湧き出し)という概念は、電磁気学の教科書で大昔たぶん習った、ラプラスの方程式、ポアソンの方程式で言ってることを数式を使わずに表現したもの。数式を使うと、電極の先端では電荷がないので、
div(grad(Vext)) = 0つまりラプラスの方程式となるが、この方程式を二つの境界条件、つまり、シナプスなどの電流源では
div(grad(Vext)) = -ρ/εポアソンの方程式が成り立つし、無限遠では
Vext = 0が成り立つ、というのを使って解くことで、Vextを計算することができる。
[Current source density (CSD)解析 その1]
上記の(3)の式にあったように、細胞外電極から記録される電位は電極近傍の電流の重み付き加算をしている。よって、一ヶ所の細胞外電位の記録だけではどこで電流流入が起こっているかを正確に推定することはできない。そこでさまざまな深さから細胞外電位を記録したデータをもとにして、電流の発生位置、つまりsinkとsourceの分布を推定する。これがcurrent source density (CSD) analysisだ。
結論だけ書くと、大脳皮質のように層構造を持っている脳部位で、深さ方向z(i) (i=1...n)で記録した細胞外電位をVext(z(i))とすると、それぞれの深さでの電流Im(z(i))は、
Im(z(i)) = -(1/R) * d2Vext(z(i))/dz2
となる(Mitzdorf 1985より)。つまり、深さz方向の軸で2階微分を計算すればよい。ただし、Rは細胞間隙の抵抗で、だいたい200-300ohm/cm。
深さ方向で等間隔に記録をしておけば、2階微分はある深さz(i)での細胞外電位Vext(z(i))とその隣り合った深さの細胞外電位Vext(z(i-1))およびVext(z(i+1))とから計算できる。
Im(z(i)) = -(1/R) * ( Vext(z(i+1)) + Vext(z(i-1)) - 2 * Vext(z(i)) )
よって、等間隔ということが非常にcriticalなので、一本の電極を進めてゆくよりかは等間隔で記録部分が並んでいるミシガン・プローブのタイプの電極を使って記録する方がよい。
くわしく導出仮定を書くと、
連続方程式より、 div(J) = Im ---(1)オームの法則より、
J = (1/R) * E ---(2)(ここでは1/R = σは均一の定数とする。)
前述の電場と電位の関係より、
E = -grad(Vext) ---(3)(2)と(3)よりEを消去すると、
J = -(1/R) * grad(Vext) ---(4)(1)と(4)からJを消去するとポアソンの方程式ができる。
(1/R) * div(grad(Vext)) = -Im ---(5)これをx,y,z成分にばらすと、
(1/R) * (∂2Vext/∂x2 + ∂2Vext/∂y2 + ∂2Vext/∂z2) = -Im ---(6)大脳皮質のように、深さz方向に延びたニューロンがx,y方向で同じように並んだ構造をしているときにはx方向、y方向の2階微分は0になるから、z成分だけが残る。
(1/R) * ∂2Vext/∂z2 = -Im ---(7)(Mitzdorf (1985)より。)
[Current source density (CSD)解析 その2]
CSD解析で得られたImから、ある時間tでの深さ方向のsink-sourceの分布を推定することができる。しかし、上記の通り、このようなsink-sourceのペアは[activeな電流(channel開閉によるイオンの移動)によるもの]と[閉回路を作成するためのpassiveな電流]のどちらに対応するかを一意に決めることはできない。そこで、解剖学から得られた細胞構築のパターンなどを援用することによって、activeな電流の位置を推定する必要がある。
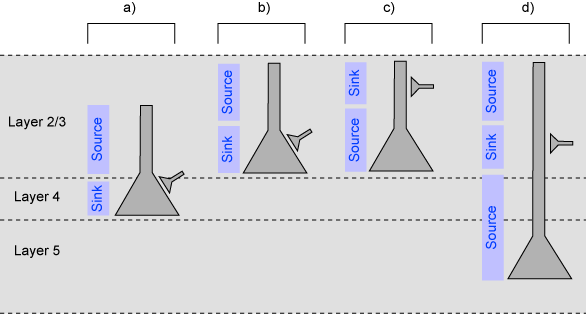
図4 CSD解析
かなり単純化した話だけれども、大脳皮質のおもなpyramidal neuronはlayer 3 (図4b-c)かlayer 5(図4d)にあって、apical dendriteを上に延ばしている。大脳皮質への入力はlayer 4 (図4a)にsomaのあるgranule cellに入力する。また、大脳皮質ではIPSCはあまりfield potentialに寄与していないことがわかっているので、EPSCだけに限る。(海馬とかだとこうはいかない。) また、sink-sourceの分布は細胞体を超えて広がることはない。
これらのことから、a)-d)のそれぞれ左にあるsink-sourceの分布パターンが見られたときには、a)-d)のそれぞれ右にあるようなニューロンへの興奮性シナプス入力が寄与していると推定することができる。
おわかりの通り、かなりざっくりとした推定なので、他の情報、たとえばsliceでの細胞内記録とそのあとのmorphologyの再構成とかと組み合わせていかないといろんなところに落とし穴がある。
このようなテクニックは1970年代後半から実用化した、けっこう古いテクニックではあるけれど、今後のLFPやMUAを使ったBMI研究においてinverse problemを解く際に、生理学的知見を用いたforward modelを作成するにあたって再び重要なテクニックとなるんではないだろうか、というのが私見。
[Single-unitの記録]
もし細胞外電極が神経細胞の非常に近傍にあるとき(<50µm)には、単一のニューロンからの活動電位を反映したsingle-unitを記録することができる。上記のMUAの特殊な例と捉えることができる。私自身はこっちが本業だけど。
教科書とかで見る細胞内電極で記録した細胞膜電位Vmはposiの山が一つで、ゆっくり落ちてくる。いっぽうで、single unitの波形Vextはそれより速くて、だいたいtriphasic。posi-nega-posi。このへんにはHenze et.al.(2000)とかの図を入れておきたい。
Single unitの波形Vextがなんであんな形をしているか。細胞内電極で記録した細胞膜電位Vmと関係づけたい。Henze et.al.(2000)では、
Vext = dVm/dt
時間ドメインでの1階微分としている。ただし、波形の後ろの方はafterhyperpolarizationとかそのへんの影響で、計算からかなりずれる。
[細胞膜電位と細胞外電位との関係の導出] (未完結)
これを導出するためには以下の手順が必要となる。(Gold et.al. (2006)より。) 複雑なのでだいたいNEURON simulatorとかが使われる。なんかもっと簡単な説明があると良いのだけれど。
- まず、Na channel、K channelの開閉によるチャネルコンダクタンスgNa, gKのダイナミクスとそれによる電流INa, IKを計算する。
- この電流によって起こる細胞膜電位Vmの変化をRallのcable方程式から計算する。
- この電流によって起こる細胞外電位Vextの変化をHolt-KochのLSA(line source approximation)によって近似的に計算する。
- 以上を(近似でいいから)解析的に解けばたぶん1階微分であることが示せるはず。
ただ、よくわからないのは、昔の本(Geddes(1972)とか)を読んでると、Lorente de No(1947)とかTasaki (1959)を引いて、2階微分だって書いてあるのがある。Axonでの記録だけど。(正確にはImとVmの関係についてだけど。) Llinas R, Nicholson C (1974)とかだと、Rallのcore conductor modelで近似できるような条件では、
the field potentials in the vicinity of extracellular sources or sinks are proportional to the first derivative of transmembrane voltage with respect to time.
とか書いてある。いっぽうで、excitable membraneでは、Imにはchannelによるcapacitanceの成分が入ってくるので、
the extracellular potential will be proportional to the second derivative of potential with respect to time
とか書いてある。たぶんこのへんを読めばわかるのかな。Lorente de Noまではさかのぼらずに済ませたいのだけれど。
そっか、基本的にはImとVmの関係さえわかればいいのだから、これは細胞外電極に限った話ではないな。Rallのcable方程式で、興奮性シナプスのある場合のバージョンを用いて、ImとVmとの関係の式にしてやればよいのだな。このへんを進めてみることにしよう。
[参考文献]
- Bedard C et.al., (2004) "Modeling extracellular field potentials and the frequency-filtering properties of extracellular space."(pdf) Biophys J. 2004 Mar;86(3):1829-42.
- G. Buzsaki (2006) Rhythms of the Brain: Oxford University Press (のうち、Cycle 4 "Windows on the brain")
- Geddes L.A. (1972) Electrodes and Measurement of Bioelectric Events (Wiley-Interscience, New York).
- Gold C et.al., (2006) "On the origin of the extracellular action potential waveform: A modeling study." J Neurophysiol. 95(5):3113-28.
- Henze, D. et.al., (2000) "Intracellular features predicted by extracellular recordings in the hippocampus in vivo." J Neurophysiol. 84, 390-400.
- Holt, GR (1998) "A Critical Reexamination of Some Assumptions and Implications of Cable Theory in Neurobiology."(pdf) Ph. D. Thesis, California Institute of Technology, Computation and Neural Systems Program
- Holt GR and Koch C. (1999) "Electrical Interactions via the Extracellular Potential Near Cell Bodies". Journal of Computational Neuroscience 6:169-184
- Kandel ER, Schwartz JH, Jessel TM, eds. (2000) "Principles of Neural Science", 4th ed. McGraw-Hill:New York, NY.
- Llinas R, Nicholson C (1974) "Analysis of field potentials in the central nervous system." In: Handbook of EEG and clinical neurophysiology (Stevens CF, ed.), pp. 61-85. Amsterdam: Elsevier.
- Logothetis NK. (2008) "What we can do and what we cannot do with fMRI." Nature 12;453(7197):869-78. (とくにsupplementary informationの"Neural Signals"のセクション)
- Mitzdorf U. (1985) "Current source-density method and application in cat cerebral cortex: investigation of evoked potentials and EEG phenomena."(pdf) Physiol Rev. 65(1):37-100.
- Nunez, P. L., & Srinivasan, R. (2006). Electric fields of the brain : The neurophysics of EEG (2nd. ed.). New York : Oxford University Press.
以上です。それで、いろいろまだ不備はあるのですが、なにがいちばん納得いってないかというと、volume conductionでのpassiveなcurrentという概念とdipoleの概念と電場の形成の概念との関係がまだ明確でないと思うのです。Dipoleによる電場の形成はdipoleの周り全体に起きることですが、passiveなcurrentの話は電荷保存の法則に従うもので、細胞膜のかたちによって影響を受け、ニューロンのサイズを超えて分布することはありません。よって二つは別ものなのだけれど、このへんをきっちりと書いている資料というものがあまりなくてわかりにくい。
それから、現在の説明(図2、図3)でもそうなのだけれど、field potentialそのものを使ってsinkとかsourceとか議論するのは正しくないわけです。しかし、われわれが慣れているのはそういう説明で、このへんが混乱の元になっているのではないかと思います。(正しい説明はたぶん、Vext = f(Im)という関係を示したあとすぐに、CSD解析でsource-sinkの推定をしたあとで、それが[activeなイオンの移動による電流]と[passiveな閉回路を作るための電流]のどちらなのかを解剖学的見地などから推測する、というものだと思われ。それはそれでわかりにくいけど。)
また、そうやって考えると、海馬で記録されるfield EPSPというのが、sinkそのものではないものを使って興奮性シナプス伝達を見るという簡便法である、ということも言えるのではないかと思います。
だいたい今年はこんなもんです。つづきはおそらくまた来年の今頃となることでしょう。ではまた。
# Ryohei
簡潔にまとまっていて、とてもよいですねー。PDFにして、講義ノートも公開してください!そういえば、もうすぐ岡崎でセミナーする予定です。お会いしたいですねー。
# pooneilどうもです。Ryoheiさん、Science出版おめでとうございます。
講義ノートの公開、いいですね。とりあえず現在のエントリのPDFファイルを作ってリンクしておきます。
内容ですけど、間違いはなかったですか? Vmの1階微分のあたりとか、それからdipoleによる電場の形成とleak currentの関係とか、直せるところがあったらぜひコメントしてください。
岡崎のセミナーは生理研ですか? 分子研ですか? ぜひお会いしましょう!
こんにちは。文献までコンパクトにまとめられていて、とても参考になります。
このエントリについて同僚と議論しているときに話題になったのですが、Single-unitのところではVextが時間微分になっているのに、細胞膜電位と細胞外電位との関係の導出のところでは空間微分になっているのはなぜなのでしょうか。
また一回微分、二階微分、2回微分という書き方が混在しているけど、どれが正しいのかというのも議論になりました。
僕は学生の頃一階微分、二階微分という風に教わりました。
なんだか指摘ばかりで申し訳ありませんが、
E(x,y,z) = -grad(V(x,y,x))
は
E(x,y,z) = -grad(V(x,y,z))
のタイポでしょうか。
# pooneilどうもです。Randy Pausch亡くなってしまいましたね。
>細胞膜電位と細胞外電位との関係の導出のところでは空間微分
しまった、ここでこの式を持ってくるのは間違いでした。とりあえず消しときます。Cable theoryあたりをもうすこし加えるy予定です。
>一階微分、二階微分
「2階微分」で統一しておきました。
>V(x,y,x)
直しておきました。
いちおうHodgkin-HuxleyのPropagationモデルでは∂2Vm/∂z2 ~∂Vm/∂tだから、その式でもよいのかも。というか、H-Hモデルなら、Vext ∝ Σi = -Cm dVm/dtでよい気がします。
# pooneilどうもありがとうございます。Im-Vmの関係だけ考えればよいというのに気がつきました。もうすこし考えてアップデートしてみます。
# Koidarman良い具合にまとまってて読んでて楽しかったです。もとは物理出身なのに、この手の数式をいじって細胞外記録データを記述するということをやったことがありませんでした。さすがです。
その昔、細胞外記録は電磁波を読み取っているんだ、という話を聞いたときに、ためしに電線にパルス電流を流して、それを近傍においた電極で読み取る計測をしてみたことがあります。
パルス波形をいろいろ変更しても、読み取った波形は結局アンプのフィルターのせいでか二相性や三相性の波形となることがわかって、パルス波形を適当にEPSPみたいな形にすると、いつも見ているような神経活動の波形が現れたので、それで納得していました。
# pooneil>ためしに電線にパルス電流を流して、それを近傍においた電極で読み取る計測をしてみた
どうもありがとうございます。さすが、これは物理やってる人の行動ですね。生理学実習でaxonを刺激して、みたいなのをやってるところだとこのへんまで勉強してるんでしょうね。
2005年08月18日
■ extracellularで記録されるsingle-unitの波形つづき
昨日のつづきです。生理学の教科書ではホジキン-ハックスレーの仕事で細胞内記録と膜チャネルの動態の話はだいたい出てくるのですが、私がこのサイトで挙げている論文で使われるような、細胞外電極によるsingle-unitの記録の原理というものはちょうどいい教科書がない、という話でした。
詳しいことは置いといて、おおまかな骨組みだけまとめておきましょう。三段階の説明のレベルがあって、それらを関連づけられるようにすればよいのかな、と考えました。第一段階は膜電位のdipoleが作る電界によるsinkとsourceを計測する、という説明。第二段階はaxon方向の一次元での等価回路を使った近似。第三段階は電流密度の連続の方程式を使った三次元での取り扱い。
まずは一番プリミティブかつ直感的な説明は、Lorente de Nó 1947などを元にした、dipoleがaxonを伝播してゆくモデル。Action potentialが起こると、一つの細胞のaxonの中で膜電位の変化が起こり、細胞膜を通した電流の流入とその周りでの流出とでループが出来る。これは細胞膜にdipoleができてそこを中心とした等電位曲線が描ける、ということ。細胞外電極はこの電位を測定している。等電位曲線に垂直な電気力線が細胞内へ流入してるところが吸い込み:sink(オシロでは下向き)で、流出がわき出し:source(上向き)。脱分極と過分極の二つのdipoleを考えると細胞外記録でのポジ-ネガ-ポジの三層性の電位変化が再現できる。
第二段階のaxon方向の一次元での等価回路では、細胞外電極を置いたところでsinkが起こったのをキルヒホッフの電流保存の式を使って定式化。細胞外電位をaxon方向の一次元で二回微分するとその場所での細胞内への電流の流入、流出が算出できる。これがcurrent source density analysisの原理。膜電位変化を時間で二回微分すると膜電流変化になる(Tasaki I)、というのもここからの帰結。Rall Wのcable theoryもこのレベルでのこと。私自身もむかしじぶんで勉強したときはこのレベルまでだったと思う。細胞内膜電位を時間で一回微分すると細胞外電位になる、というのもこのレベルの話のはずなんだけど、元ネタ見つからず。
第三段階の電位ポテンシャルに関するラプラス-ポワソン方程式を使った三次元での取り扱い。電荷がない点では電流のsinkもsourceもないので電界の発散はない(divE=0)。膜電流のあるところでの電荷密度の時間変化は電界の発散に比例する(連続の方程式)。このレベルで、さまざまな仮定が第一段階や第二段階で置かれていることを明示的に扱えるようになる。現在のモデル屋さんはこのレベルを扱っている。ベクトル解析で電磁気学と流体力学の対応とか読んでるとおもしろいけど、生物系の教養課程ではそのへんまでたどり着いていなかったような。
んでもって、これらのレベル間での対応を付ける。各レベルでのsinkとsourceの扱いとか、第二段階でのキルヒホッフの電流保存の式と第三段階での連続の式との対応付けとか。 第三段階の式からcurrent source densityの式を導出とか。
とだいたいこんな感じになるのではないかと。前回書いたエントリでは、この第二段階が抜けていたからギャップを感じていたのですな。むかし勉強したことをすこし思い出してきました。
これを肉づけして、かんたんに説明できる資料が作れたらいいかな。ここで書いていることの完成版を作るのに私より向いている人はたくさんいるはずですし、どなたかぜひ。
Evoked field potentialとかLFPのoscillationとかを議論するにはまだ遠そうです。むかしラット海馬のslice cultureでevoked field potentialのLTPをやってたんですけど、そのときにはSchaffer collateralの入力層で記録すればfield EPSP(ネガ一層性)、pyramidal cell layerで記録すればpopulation spike(ポジ-ネガ-ポジの三層性)、と納得いってた気がするけど、somaでのcurrentのvolume conductionとか、dendriteへのaction potentialのpropagationとか考えるとじつはかなり深い。当時は疑問に思ってなかったけど、なんでfield EPSPにはsomaのspikeのvolume conductionがかぶってこないんだろう。Population spikeのポジにはdendriteでのEPSPのvolume conductionがかぶっているなんて理解してたけど、それでは対称性がない。それとも充分離れていれば、それこそextracellular mediumがinhomogeneousだからspikeのような速い周波数成分はさっさと減衰してしまうということか。海馬のような解剖学的に均一な構造ですらこんな調子なのだから、ましてやcortexとかは私にとっては複雑すぎ。
ところで私自身はこのへんのバックグラウンドが全然ないままにこれを書いているので、かなり恥をさらしています。つっこみ、訂正歓迎します。たとえば、ガウスの法則の微分形式と、ラプラス-ポワソン方程式と、連続の方程式の関係とか……本質的に同じなんだろうと思うのですが、系統的にそういうことを勉強した経験がありません。第二段階でのキルヒホッフの電流保存の式と第三段階での連続の方程式との関係とかも。物理のバックグラウンドを持っている方からすれば、教養課程レベルの知識もないことがモロばれでしょう。アンプを自作していた時代の生理学者から見れば嘆かわしいことに違いありません。まあ、なにより自分の勉強のため、ということで明日のために。
つづきがあります:20080727
2005年08月17日
■ extracellularで記録されるsingle-unitの波形
LFP関連ですが、研究所のトレーニングコースで、LFPのできるメカニズムを簡単にレクチャーできるようにと思っていろいろ資料を集めてたのですが、これが意外に難しい。そもそも教科書的にはホジキン-ハックスレーでイカのaxonからintraとかそういう話になってしまっていて、extracellularで記録されるsingle-unitの波形がどうできるかを説明している教科書というものはわたしが知る限りではよいものがありません。かたやsourceとsink、といった端的な説明の仕方(*1)になってしまうし、かたやラプラス・ポアソン方程式になってしまう(*2)。なんかその中間をいくようなうまい説明の仕方はないか思案中です。というわけで、いまさら電磁気学の勉強です。おすすめの資料がありましたら推薦歓迎します。
*1: たとえば、Roger Lemonの"Methods for neuronal recording in conscious animals"にあるように。
*2: Mitzdorfのcurrent source density analysisの論文 physiological review '85 "Current source-density method and application in cat cerebral cortex: investigation of evoked potentials and EEG phenomena"(pdf)とか、Alain DestexheのBiophysical Journal '04 "Modeling Extracellular Field Potentials and the Frequency-Filtering Properties of Extracellular Space"(著者サイトのpdfファイル)とか。
追記:STさんありがとうございます。推薦論文をリンクしておきます:
"Intracellular features predicted by extracellular recordings in the hippocampus in vivo." Henze, D., Borhegyi, Z., Csicsvari, J., Mamiya, A., Harris, K. and Buzsáki, G., 2000. J Neurophysiol. 84, 390-400.
"Electrical Interactions via the Extracellular Potential Near Cell Bodies". G. R. Holt and C. Koch. Journal of Computational Neuroscience 6:169-184, 1999
A Critical Reexamination of Some Assumptions and Implications of Cable Theory in Neurobiology.(pdf file) Holt, GR Ph. D. Thesis, California Institute of Technology, Computation and Neural Systems Program (1998)
上記のDestexhe論文ではKoch and Segevの"Methods in Neuronal Modeling" 1998がLFPの簡単なモデル(extracellular mediumがhomogeneousであることを仮定)の例としてreferされてます。
次回へ続きます。
# ST
最近の資料として以下のようなものはいかがでしょうか。
Henze, D., Borhegyi, Z., Csicsvari, J., Mamiya, A., Harris, K. and Buzsaki, G., 2000. Intracellular features predicted by extracellular recordings in the hippocampus in vivo. J Neurophysiol. 84, 390-400.
Electrical Interactions via the Extracellular Potential Near Cell Bodies. G. R. Holt and C. Koch. Journal of Computational Neuroscience 6:169-184, 1999.
確かHoltのdissertaionもWeb上にあったような。
STさん、ありがとうございます。さっそくリストに追加しておきました。とくにBuzsákiのほうは、in vivoの皮質でのintraとextraを比較して提示するのによい資料になりそうです。
お勧めエントリ
- 細胞外電極はなにを見ているか(1) 20080727 (2) リニューアル版 20081107
- 総説 長期記憶の脳内メカニズム 20100909
- 駒場講義2013 「意識の科学的研究 - 盲視を起点に」20130626
- 駒場講義2012レジメ 意識と注意の脳内メカニズム(1) 注意 20121010 (2) 意識 20121011
- 視覚、注意、言語で3*2の背側、腹側経路説 20140119
- 脳科学辞典の項目書いた 「盲視」 20130407
- 脳科学辞典の項目書いた 「気づき」 20130228
- 脳科学辞典の項目書いた 「サリエンシー」 20121224
- 脳科学辞典の項目書いた 「マイクロサッケード」 20121227
- 盲視でおこる「なにかあるかんじ」 20110126
- DKL色空間についてまとめ 20090113
- 科学基礎論学会 秋の研究例会 ワークショップ「意識の神経科学と神経現象学」レジメ 20131102
- ギャラガー&ザハヴィ『現象学的な心』合評会レジメ 20130628
- Marrのrepresentationとprocessをベイトソン流に解釈する (1) 20100317 (2) 20100317
- 半側空間無視と同名半盲とは区別できるか?(1) 20080220 (2) 半側空間無視の原因部位は? 20080221
- MarrのVisionの最初と最後だけを読む 20071213